Островной гигантизм - Island gigantism

Островной гигантизм, или же островной гигантизм, представляет собой биологическое явление, при котором размер вида животных, изолированных на острове, резко увеличивается по сравнению с его родственниками на материке. Островной гигантизм - один из аспектов более общего "эффект острова" или "правило Фостера", который утверждает, что когда материковые животные колонизируют острова, мелкие виды имеют тенденцию эволюционировать более крупные тела, а крупные виды имеют тенденцию эволюционировать более мелкие тела (островная карликовость ). После прибытия людей и связанных с ними интродуцированных хищников (собак, кошек, крыс, свиней) многие гигантские, а также другие островные эндемики стали вымерший. Подобное увеличение размеров, а также усиление древесности наблюдается у некоторых островных растений.
Возможные причины

Крупные плотоядные млекопитающие часто отсутствуют на островах из-за недостаточного ареала или трудностей с их обитанием. рассеяние над водой. В их отсутствие экологические ниши для крупных хищников они могут быть заняты птицами, рептилиями или более мелкими хищниками, которые затем могут вырасти до больших, чем обычно, размеров. Например, на доисторических Остров Гаргано в Миоцен -Плиоцен Средиземноморье, на островах в Карибский бассейн подобно Куба, и дальше Мадагаскар и Новая Зеландия, некоторые или все высшие хищники были птицы как орлы, соколы и совы, включая некоторые из самых крупных известных примеров этих групп. Однако птицы и рептилии, как правило, становятся менее эффективными крупными хищниками, чем развитые. плотоядные.
Поскольку небольшой размер обычно облегчает травоядные животные Чтобы убежать или спрятаться от хищников, снижение давления хищников на острова может позволить им вырасти.[1][а] Мелкие травоядные также могут извлечь выгоду из отсутствия конкуренции со стороны отсутствующих типов крупных травоядных.
Преимущества большого размера, которые были предложены для островных черепах, включают снижение уязвимости к нехватке пищи и / или воды, способность выжить в течение более длительных интервалов без них или способность преодолевать большие расстояния, чтобы получить их. Периоды такой нехватки могут быть большей угрозой для океанических островов, чем на материке.[4]
Таким образом, островной гигантизм обычно представляет собой эволюционную тенденцию, возникающую в результате снятия ограничений на размер мелких животных, связанных с хищничеством и / или конкуренцией.[5] Однако такие ограничения могут действовать по-разному в зависимости от размера животного; например, в то время как мелкие травоядные животные могут избежать хищников, скрывшись, крупные травоядные могут отпугивать хищников запугиванием. В результате дополнительный феномен островная карликовость может также быть результатом снятия ограничений, связанных с хищничеством и / или конкуренцией в отношении размера крупных травоядных.[6] Напротив, островная карликовость среди хищников чаще возникает в результате введения ограничений, связанных с ограниченными ресурсами добычи, доступными на островах.[6] В отличие от островной карликовости, островной гигантизм встречается в большинстве крупных позвоночное животное группы и в беспозвоночные.
Территориализм может способствовать развитию островного гигантизма. Исследование на Остров Анахо в Неваде определили, что территориальные виды рептилий имеют тенденцию быть крупнее на острове по сравнению с материком, особенно среди более мелких видов. У территориальных видов больший размер позволяет особям лучше конкурировать за защиту своей территории. Это дает дополнительный импульс эволюции в сторону увеличения размера островной популяции.[7]
Еще одним средством установления островного гигантизма может быть эффект основателя действует, когда более крупные члены материкового населения превосходят в своих способностях колонизировать острова.[8]
Размер острова играет роль в определении степени гигантизма. Более мелкие острова, как правило, ускоряют скорость эволюции изменений размеров организмов, а организмы на этих островах развиваются в больших размерах.[9]
Примеры
Примеры островного гигантизма включают:
Млекопитающие
Многие грызуны вырастают на островах, тогда как плотоядные, хоботки и парнокопытные обычно становятся меньше.
Eulipotyphlans
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Корсиканская гигантская землеройка | Asoriculus corsicanus | Корсика | Вымерший (до 500 г. до н.э.) |  Краснозубые землеройки |
| Балеарская гигантская землеройка | Asoriculus hidalgo | Майорка и Менорка | Вымерший | |
| Сардинская гигантская землеройка | Asoriculus similis | Сардиния | Вымерший | |
 Дейногалерикс | Дейногалерикс виды | Остров Гаргано | Вымерший (Поздний миоцен ) |  Лунные крысы |
Грызунов
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник | Островной / материк соотношение длины или массы |
|---|---|---|---|---|---|
| Тупозубая гигантская хутия | Amblyrhiza inundata | Ангилья и Сен-Мартен | Вымерший (Плейстоцен ) |  Неотропические колючие крысы | |
| Большая ямайская гигантская хутия | Clidomys osborni | Ямайка | Вымерший (Поздний плейстоцен) | ||
| Гигантская хутия с пластинчатыми зубами | Косой эластомодонтомис | Пуэрто-Рико | Вымерший (ок. 1 н.э.) | ||
| Мышь с витыми зубьями | Quemisia Gravis | Hispaniola | Вымерший | ||
| Древесная гигантская хутия[10] | Тайнотерий валей | Пуэрто-Рико | Вымерший | ||
| Малая ямайская гигантская хутия | Xaymaca fulvopulvis | Ямайка | Вымерший | ||
| Майоркский гигант хомяки | Apocricetus darderi Tragomys macpheei | Майорка | Вымерший | Apocricetus alberti[11] Cricetus кормоси[12] | |
 Гигантский хомяк Гаргано | Hattomys gargantua | Остров Гаргано | Вымерший | ||
 Полевая мышь St Kilda | Apodemus sylvaticus hirtensis | Сент-Кильда | Наименьшие опасения |  Деревянная мышь | MR ≈ 2 [13] |
 Полевая мышь Хензеля | Rhagamys orthodon | Корсика и Сардиния | Вымерший (Поздний плейстоцен ) | ||
 Гигантская крыса Тенерифе | Canariomys bravoi | Тенерифе | Вымерший (Поздний плейстоцен) | Африканские рыжие крысы | |
 Гигантская крыса Гран-Канарии | Canariomys tamarani | Гран-Канария | Вымерший (до 1500 г.) | ||
| Чернохвостая садовая соня Форментера | Eliomys quercinus ophiusae | Форментера | Редкий[14] |  Садовая соня и Другой Leithiinae соня | |
| Миноркская гигантская соня | Гипномис махоненсис | Менорка | Вымерший | ||
 Майоркская гигантская соня | Гипномис морфеус | Майорка | Вымерший | ||
| Сицилийская гигантская соня | Leithia картей | Сицилия | Вымерший | ||
| Мальтийская гигантская соня | Leithia melitensis | Мальта | Вымерший | ||
 Оркнейская полевка | Microtus arvalis orcadensis | Оркнейские острова | Уязвимый |  Обыкновенная полевка и Другой луговые полевки | |
 Гигантские полевки Гаргано | Mikrotia magna М. maiuscula М. Парва | Остров Гаргано | Вымерший (Ранний плиоцен) | ||
 Домовая мышь St Kilda | Mus musculus muralis | Сент-Кильда | Вымерший (ок. 1930 г.) |  Домовая мышь | |
 Гигантская крыса Флореса | Папагомыс armandvillei | Флорес | Под угрозой |  Северная Африка черная крыса и другие настоящие крысы | |
| Сулавесская гигантская крыса | Паруромыс доминатор | Сулавеси | Наименьшие опасения | ||
| Адмиралтейство гигантская крыса | Rattus detentus | Остров Мануса | Неизвестно / Вероятная угроза[15] | ||
| Популяция черных крыс Congreso[16] | Раттус раттус | Исла-дель-Конгресо | Наименьшие опасения | ||
| Мыши-олени Нормандских островов | Peromyscus anyapahensis P. nesodytes | Северные Нормандские острова Калифорнии | Вымерший (ок. 6000 г. до н.э.) |  Североамериканская олень мышь | |
 Гигантская соня Гаргано | Stertomys laticrestatus[17] | Остров Гаргано | Вымерший |  Glirinae соня |
Зайцеобразные
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Миноркский гигантский лагоморф | Нуралаг Рекс | Менорка | Вымерший (Средний плиоцен ) | Алилепус (?) Тришизолагус (?) |
| Prolagus imperialis | Остров Гаргано | Вымерший |  Пикас | |
 Сардинская пищуха | Prolagus sardus | Корсика, Сардиния и Таволара | Вымерший (ок. 1800 г.) |
Приматы
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Испанская обезьяна | Antillothrix bernensis | Hispaniola | Вымерший (до 1600 г.) |  Cheracebus |
| Гаитянская обезьяна | Insulacebus toussaintiana | Юго-западный Гаити | Вымерший | |
 Кубинские обезьяны | Паралуатта марианская[18] P. varonai[18] | Куба | Вымерший (Плейстоцен) | |
| Ямайская обезьяна | Ксенотрикс макгрегори | Ямайка | Вымерший | |
 Горилла лемур | Archaeoindris fontoynontii | Центральная Мадагаскар | Вымерший (ок. 350 г. до н.э.) |  Лорисоиды |
 Лемуры бабуина | Археолемур виды Hadropithecus виды | Мадагаскар | Вымерший (до 1280 г.) | |
 Ленивые лемуры | Бабакотия виды Палеопропитек виды | Западный и Центральный Мадагаскар | Вымерший (ок. 1500 г.) | |
 Коала лемуры | Megaladapis edwardsi М. grandidieri М. madagascariensis | Мадагаскар | Вымерший (1280-1420 гг. Нашей эры) |
Плотоядные
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Сардинская гигантская выдра | Megalenhydris barbaricina | Сардиния | Вымерший (Поздний плейстоцен) |  Выдры |
 Ямка | Криптопрокта ферокс | Мадагаскар | Уязвимый |  Мангусты |
 Гигантская ямка | Cryptoprocta spelaea | Мадагаскар | Вымерший (до 1400 г.) |
Птицы
Стеблевые птицы
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Балаур | B. bondoc | Остров Хатег | Вымерший (Поздний мел) |  Jeholornis[19] |
 Гаргантюавис | Г. Филохинос | Иберо -Armorican Остров | Вымерший (Поздний мел ) |  Патагоптерикс (?) |
Ratites
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Киви | Аптерикс виды | Новая Зеландия | Переменная | Проаптерикс[b] |
 Больше слон птицы | Эпиорнис Хильдебрандти A. maximus Воромбе титан | Мадагаскар | Вымерший (ок. 1000 г. н.э.) | |
 Малые птицы-слоны | Mullerornis виды | Мадагаскар | Вымерший (ок. 1260 г.) | |
 Моа | Аномалоптерикс дидиформ Dinornis виды Эмеус Красс Эвриаптерикс виды Мегалаптерикс дидинус Пахиорнис виды | Новая Зеландия | Вымерший (до 1445 г.) |  Tinamous |
Водоплавающих птиц
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Новозеландская мускусная утка | Biziura delautouri | Новая Зеландия | Вымерший (после 1500 г.) |  Австралийский мускусная утка |
 Новозеландские гуси | Cnemiornis calcitrans C. gracilis | Новая Зеландия | Вымерший |  Мыс Бесплодный гусь |
 Гарганорнис | Г. баллманни | Гаргано и Scontrone острова | Вымерший (Поздний миоцен ) |  Гуси[21] |
 Моа-нало с черепаховой пастью | Челычилинеченский квас | Кауаи | Вымерший (ок. 1000 г. н.э.) |  Веселые утки |
 Мелкоклювый моа-нало | Ptaiochen pau | Мауи | Вымерший (ок. 1000 г. н.э.) | |
Моа-нало крупноклювый | Thambetochen chauliodous | Мауи Нуи | Вымерший (ок. 1000 г. н.э.) | |
 Оаху моа-нало | Thambetochen xanion | Оаху | Вымерший (ок. 1000 г. н.э.) | |
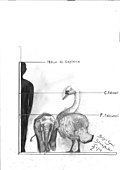 Гигантский лебедь | Cygnus falconeri | Сицилия и Мальта | Вымерший (Средний плейстоцен ) |  Лебедь-шипун |
| Утка Скарлетт | Malacorhynchus scarletti | Новая Зеландия | Вымерший (после 1500 г.) |  Утка с розовыми ушами |
Пангаллообразные
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Свайный мегапод | Мегаподиусный молиструктор | Новая Каледония и Тонга | Вымерший (ок. 1500 г. до н.э.) |  Птица |
| Благородный мегапод | Megavitiornis altirostris | Фиджи | Вымерший |  Galliformes |
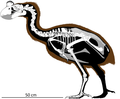 Гигантский мегапод Новой Каледонии | Sylviornis neocaledoniae | Новая Каледония и Остров Пайнс | Вымерший |
Gruiformes
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Красный рельс | Aphanapteryx bonasia | Маврикий | Вымерший (ок. 1700 г.) |  Рельсы |
 Рельс Хокинса | Diaphanapteryx hawkinsi | Острова Чатем | Вымерший (ок. 1900 г.) | |
 Антильские пещеры | Nesotrochis debooyi | Пуэрто-Рико и Виргинские острова | Вымерший | |
| Кубинская пещерная ограда | Несотрочис Picapicensis | Куба | Вымерший | |
| Пещерный перила Гаити | Nesotrochis steganinos | Hispaniola | Вымерший | |
 Южный остров такахэ | Porphyrio hochstetteri | Южный остров, Новая Зеландия | Находящихся под угрозой исчезновения | |
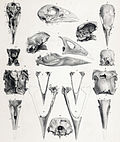 Такахэ Северного острова | Порфирио мантелли | Северный остров, Новая Зеландия | Вымерший (до 1900 г.) | |
 Adzebills | Дефоссор апторниса А. otidiformis | Новая Зеландия | Вымерший |  Мадагаскарский пушистый хвост[22] |
 Chatham проститутка | Fulica chathamensis | Острова Чатем | Вымерший (после 1500 г.) |  Краснобородая лысуха и другие лысухи |
 Маскаренская лысуха | Fulica newtonii | Маврикий и Реюньон | Вымерший (ок. 1700 г.) | |
| Новозеландская лысуха | Fulica Prisca | Новая Зеландия | Вымерший (после 1280 г.) | |
 Реюньон болото | Porphyrio coerulescens | Plaine des Cafres, Реюньон | Вымерший (ок. 1730 г.) |  Фиолетовые болота |
Голуби
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Вити-Леву гигантский голубь | Natunaornis gigoura | Вити Леву, Фиджи | Вымерший |  Венценосные голуби |
 Пасьянс Родригес | Pezophaps solitaria | Родригес | Вымерший (до 1778 г.) |  Никобарский голубь |
 Додо | Raphus cucullatus | Маврикий | Вымерший (ок. 1662 г.) |
Хищные птицы
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Пещера Лико беркут | Aquila chrysaetos simurgh | Крит | Вымерший (Поздний плейстоцен) |  Золотой орел |
| Гигантский краб-ястреб[23] | Buteogallus borrasi | Куба | Вымерший |  Большой черный ястреб и другие ястребы |
| Гигантский ястреб | Гигантохиеракс sp. | Куба | Вымерший | |
| Титан-ястреб | Titanohierax gloveralleni | Куба, Hispaniola и Багамы | Вымерший | |
| Ямайский каракара | Каракара теллустрис | Ямайка | Вымерший |  Каракарас |
| Лунь глазка | Цирк Эйлези | Новая Зеландия | Вымерший (ок. 1000 г.) |  Болотный лунь |
| Орлы острова Гаргано | Garganoaetus freudenthali G. murivorus | Остров Гаргано | Вымерший (Поздний миоцен) | Aquila дельфин |
 Орел Хааста | Hieraaetus moorei | Новая Зеландия | Вымерший (ок. 1400 г.) |  Маленький орел |
 Филиппинский орел | Pithecophaga jefferyi | Филиппины | Находящихся под угрозой исчезновения |  Bateleur[24] |
Попугаи
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Попугай геркулес | Геракл неизведанный | Новая Зеландия | Вымерший (Миоцен ) |  Другие попугаи |
 Какапо | Strigops habroptilus | Новая Зеландия | Находящихся под угрозой исчезновения | |
 Ширококлювый попугай | Lophopsittacus mauritianus | Маврикий | Вымерший (ок. 1680 г.) |  Пситтакулин попугаи |
Совы
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Критская сова | Афина кретенсис | Крит | Вымерший (Плейстоцен) |  Маленькая сова |
 Кубинские гигантские совы | Орнимегалоникс виды | Куба | Вымерший (Плейстоцен) |  Лесные совы |
 Большая гигантская сова Гаргано | Tyto gigantea | Остров Гаргано | Вымерший (Поздний миоцен ) |  Сипухи |
 Сипуха на острове Андрос | Тито пыльца | Остров Андрос, Багамы | Вымерший (до 1600 г.) | |
 Сипуха Риверо | Tyto riveroi | Куба | Вымерший | |
 Гигантская сова Малый Гаргано | Tyto robusta | Остров Гаргано | Вымерший (Ранний плиоцен ) |
Caprimulgiformes
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Новозеландский совенок-козодой | Aegotheles novazelandiae | Новая Зеландия | Вымерший (ок. 1200 г. н.э.) |  Австралийский совенок-козодой |
 Новокаледонский совенок-козодой | Эготелес Савеси | Новая Каледония | Находящихся под угрозой исчезновения |
Passeriforms
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Чатемский ворон | Corvus moriorum | Острова Чатем | Вымерший | Новозеландский ворон |
 Длинноногая овсянка | Эмбериза альковери | Тенерифе | Вымерший (после 1 г. н.э.) |  Овсянка кабаниса |
| Гигантская нукупу'у | Hemignathus vorpalis | Гавайи | Вымерший (после 1000 г. н.э.) |  Зяблики |
| Превосходный тасманский крапивник | Malurus cyaneus cyaneus | Тасмания | Наименьшие опасения |  Великолепный крапивник |
| Великолепные крапивницы с острова Кенгуру | Malurus cyaneus ashbyi | Остров Кенгуру | Наименьшие опасения | |
| Толстоногий крапивник | Pachyplichas yaldwyni | Южный остров из Новая Зеландия | Вымерший |  Другие прохожие |
 Козерог серебристый | Zosterops lateralis chlorocephalus | Козерог и Бункер Групп из Австралийский Большой Барьерный риф | Неизвестный |  Сильверай |
Рептилии
Птерозавры
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Hatzegopteryx | H. thambema | Остров Хатег | Вымерший (Поздний мел) |  Quetzalcoatlus |
Игуаниды
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник | Островной / материк соотношение длины или массы |
|---|---|---|---|---|---|
 Тонга гигантская игуана[25] | Brachylophus gibbonsi | Тонга | Вымерший (ок. 800 г. до н.э.) |  южноамериканец игуаны | |
 Фиджийская гигантская игуана [26] | Lapitiguana impensa | Фиджи | Вымерший (ок. 1000 г. до н.э.) | ||
 Остров ангелов чаквалла | Sauromalus hispidus | Исла Анхель де ла Гуарда, Нижняя Калифорния | Под угрозой | Полуостров Чаквалла | MR ≈ 5 [27] |
 Сан-Эстебан Чаквалла | Sauromalus varius | Остров Сан-Эстебан, Нижняя Калифорния | Находящихся под угрозой исчезновения | MR ≈ 5 [27] |
Гекконы
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник | Островной / материк соотношение длины или массы |
|---|---|---|---|---|---|
 Гигантское гекко Делькура | Hoplodactylus delcourti | Новая Зеландия | Вымерший (ок. 1870 г.) |  Диплодактилидные гекконы | LR ≈ 6,75 [c] |
 Новокаледонский гигантский геккон | Rhacodactylus leachianus | Новая Каледония | Наименьшие опасения | LR ≈ 4,4 [d] MR ≈ 60 [e] | |
 Родригес гигантский дневной геккон | Phelsuma gigas | Родригес | Вымерший (ок. 1850 г.) |  Дневные гекконы |
Сцинки
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Мабуя Вайланта | Chioninia vaillanti | Кабо-Верде | Находящихся под угрозой исчезновения |  Материк мабуйин сцинки |
 Кабо-Верде гигантский сцинк | Macroscincus coctei | Кабо-Верде | Вымерший (после 1900 г.) | |
 Гигантский сцинк Маврикия | Leiolopisma mauritiana | Маврикий | Вымерший (после 1600 г.) | Материк евгонгилин сцинки |
| Сцинк ужаса | Phoboscincus bocourti | Desle des Pins выключенный Новая Каледония | Находящихся под угрозой исчезновения |  Материк евгонгилин сцинки |
| Гигантский сцинк кишиноуэ | Плестиодон кишинуйей | Острова Мияко и Острова Яэяма, Япония | Уязвимый |  Азиатский Плестиодон виды |
Настенные ящерицы
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Гигантская ящерица Ла Пальмы | Gallotia auaritae | Ла Пальма | Находящихся под угрозой исчезновения |  Средиземноморские ящерицы-песчаники |
 Гигантская ящерица Ла Гомера | Галлотия бравоана | Гомера | Находящихся под угрозой исчезновения | |
 Гигантская ящерица Тенерифе[31] | Галлотия голиаф | Тенерифе | Вымерший (ок. 1500 г.) | |
 Гигантская ящерица Эль Йерро | Gallotia simonyi | Эль Йерро | Находящихся под угрозой исчезновения | |
 Гигантская ящерица Гран-Канарии | Gallotia stehlini | Гран-Канария | Наименьшие опасения |
Змеи
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
| Крапчатая гремучая змея на острове Анхель де ла Гуарда | Crotalus mitchellii angelensis | Исла Анхель де ла Гуарда выключенный Нижняя Калифорния | Наименьшие опасения |  Крапчатая гремучая змея |
| Популяция полосатых змей таданаэдзима[32] | Elaphe quadrivirgata | Остров Таданаэ-Джима Токио | Неизвестный |  Японская полосатая змея |
 Популяции островных тигровых змей | Notechis scutatus | Острова Mount Chappell, Уильямс, Хопкинс, и Нуйцкий архипелаг (Южная Австралия )[33] | Наименьшие опасения[34] |  Тигровая змея |
| Длинноносая змея Isla Cerralvo | Rhinocheilus lecontei etheridgei | Остров Жака Кусто выключенный Южная Нижняя Калифорния | Неизвестный |  Длинноносая змея |
Сомнительные примеры


- В Дракон Комодо из Флорес и близлежащие острова, самая большая из ныне живущих ящериц и аналогичный (вымерший) гигант варан ящерица из Тимор считались примерами гигантских островных хищников. Поскольку острова, как правило, предлагают ограниченную пищу и территорию, их хищные млекопитающие (если они есть) обычно меньше континентальных. Эти дела включают экзотермический плотоядные животные на островах слишком малы, чтобы содержать много млекопитающее конкуренция. Однако эти ящерицы не такие большие, как их вымершие австралийские родственники. мегалания, и было предложено на основе свидетельств окаменелостей, что предки этих варанид сначала развили свои большие размеры в Австралии, а затем расселились в Индонезии.[35] Если это правда, они будут рассматриваться не как островные гиганты, а как примеры филетический гигантизм. Эта интерпретация подтверждается свидетельствами существования ящерицы в плиоценовой Индии, Varanus sivalensis, по размеру сопоставимый с komodoensis.[35] Тем не менее, учитывая, что Австралия часто описывается как самый большой остров в мире и что связанная с этим мегалания, самая крупная наземная ящерица, известная в летописи окаменелостей, была ограничена Австралией, восприятие крупнейших австралийских / индонезийских ящериц как островных гигантов все еще может иметь определенное значение.
- Гигантские черепахи в Галапагосские острова и Сейшельские острова, самые большие из современных черепах, а также вымершие черепахи Маскарены и Канарские острова, часто считаются образцами островного гигантизма. Однако во время плейстоцена черепахи сопоставимого размера или большего размера присутствовали в Австралия (Мейолания ), южный Азия (Мегалохелис ), Мадагаскар (Алдабрачелис ), Северная Америка[36] (Hesperotestudo ) и Южная Америка[37] (Chelonoidis, тот же род, который сейчас встречается на Галапагосских островах[38]), а также на ряде других, более доступных островов.[36] В позднем плиоцене они также присутствовали в Африке.[39] ("Геохелоне " laetoliensis[39]). Нынешняя ситуация, когда крупные черепахи встречаются только на удаленных островах, по-видимому, отражает то, что эти острова были обнаружены людьми недавно и не были густонаселенными, что делает их черепах менее уязвимыми. чрезмерная эксплуатация.
Амфибии
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник | Островной / материк соотношение длины или массы |
|---|---|---|---|---|---|
| Гигантская древесная лягушка Сан-Томе | Hyperolius thomensis[40] | Остров Сан-Томе | Находящихся под угрозой исчезновения |  Африканские камышовые лягушки | |
| Лягушка из пальмового леса | Лептопелис пальматус[40] | Príncipe Остров | Уязвимый |  Красная квакша | LR ≈ 1,2 [f] |
| Гигантская наземная лягушка Фиджи | Platymantis megabotoniviti[43] | Вити Леву, Фиджи | Вымерший |  Азиатский платимантины | |
| Гигантская травяная лягушка Сан-Томе | Ptychadena newtoni[40] | Остров Сан-Томе | Находящихся под угрозой исчезновения |  Маскаренская травяная лягушка |
Членистоногие





















Брюхоногие моллюски
| Пример | Биномиальное имя | Родной диапазон | Текущее состояние | Континентальный родственник |
|---|---|---|---|---|
 Сухопутные улитки каури | Парифанта виды Powelliphanta виды | Новая Зеландия | Под угрозой |  Другие ритидиды |
Флора
Помимо увеличения размера, остров трава растения могут также демонстрировать "островную древесность". Наиболее яркими примерами являются мегатравы из Новая Зеландия с субантарктические острова.[47] Также сообщалось об увеличении размера листьев и семян у некоторых островных видов независимо от формы роста (травянистые, куст, или же дерево ).[48]




















Смотрите также
Примечания
- ^ Снижение хищничества на островах часто также приводит к более прирученному поведению островных видов добычи, тенденция, которая была проанализирована на ящерицах.[2][3]
- ^ Самый ранний из известных новозеландских предков киви, предположительно недавно прибывший из Австралии.[20]
- ^ На основе предполагаемой общей длины Х. делькурти, ~ 23,6 дюйма,[28] и средняя длина члена Диплодактиль, наиболее специфичный род австралийских диплодактилид гекконов, ~ 3.5 дюйма.[29]
- ^ Исходя из средней общей длины более крупных подвидов, Р. л. Leachianus, ~ 15,5 дюйма,[30] и средняя длина члена Диплодактиль, наиболее специфичный род австралийских диплодактилид гекконов, ~ 3.5 дюйма.[29]
- ^ Исходя из средней массы более крупных подвидов, Р. л. Leachianus, ~ 240 г,[30] со средним весом члена Диплодактиль, наиболее видный род австралийских диплодактилидных гекконов, ~ 4 г.[29]
- ^ На основе средней длины морды самки до ствола (SVL) L. palmatus, ~ 96 мм,[41] со средней женской СВЛ L. rufus, ~ 80 мм.[42]
Рекомендации
- ^ Herczeg, G. B .; Gonda, A. L .; Мериля, Дж. (16 июля 2009 г.). «Эволюция гигантизма у девятииглой колюшки». Эволюция. 63 (12): 3190–3200. Дои:10.1111 / j.1558-5646.2009.00781.x. PMID 19624722. S2CID 205782326.
- ^ Cooper, W. E .; Pyron, R.A .; Гарланд, Т. (2014-01-08). «Приручение острова: жизнь на островах сокращает расстояние начала полета». Труды Королевского общества B: биологические науки. 281 (1777): 20133019. Дои:10.1098 / rspb.2013.3019. ЧВК 3896029. PMID 24403345.
- ^ Йонг, Э. (2014-01-08). «Острова делают животных укротителями». Природа. Дои:10.1038 / природа.2014.14462. S2CID 183158746.
- ^ Jaffe, A. L .; Слейтер, Г. Дж .; Альфаро, М. Э. (26 января 2011 г.). «Эволюция островного гигантизма и изменение размеров тела у черепах и черепах». Письма о биологии. 7 (4): 558–561. Дои:10.1098 / рсбл.2010.1084. ЧВК 3130210. PMID 21270022.
- ^ Barahona, F .; Evans, S.E .; Mateo, J.A .; Garcia-Marquez, M .; Лопес-Хурадо, Л.Ф. (март 2000 г.). "Эндемизм, гигантизм и исчезновение островных ящериц: род Галлотия на Канарских островах ». Журнал зоологии. 250 (3): 373–388. Дои:10.1017 / s0952836900003101. HDL:10553/19918.
- ^ а б Raia, P .; Мейри, С. (август 2006 г.). «Островное правило у крупных млекопитающих: палеонтология встречается с экологией». Эволюция. 60 (8): 1731–1742. Дои:10.1111 / j.0014-3820.2006.tb00516.x. PMID 17017072. S2CID 26853128.
- ^ Keehn, J. E .; Nieto, N.C .; Tracy, C. R .; Gienger, C.M .; Фельдман, К. Р. (27 августа 2013 г.). «Эволюция на необитаемом острове: расхождение в размерах тела рептилий острова Анахо в Неваде и материка вокруг озера Пирамид». Журнал зоологии. 291 (4): 269–278. Дои:10.1111 / jzo.12066.
- ^ Ломолино, М. В. (05.09.2005). «Эволюция размеров тела у островных позвоночных: общее правило острова». Журнал биогеографии. 32 (10): 1683–1699. Дои:10.1111 / j.1365-2699.2005.01314.x. HDL:2027.42/146565.
- ^ Филин, И .; Зив, Ю. (2004). «Новая теория островной эволюции: объединение потери дисперсности и изменения массы тела» (PDF). Исследования эволюционной экологии. 6: 115–124.
- ^ Турви, С. Т. (2006). «Новый род и вид гигантской хутии (Тайнотерий валей) из четвертичного периода Пуэрто-Рико: вымершее древесное четвероногое животное? ". Журнал зоологии. 270 (4): 585–594. Дои:10.1111 / j.1469-7998.2006.00170.x.
- ^ Torres-Roig, E .; Agustí, J .; Bover, P .; Альковер, Дж. (2017). «Новый гигантский крикетин из базального плиоцена Майорки (Балеарские острова, западное Средиземноморье): биостратиграфическая связь с зонами континентальных млекопитающих». Историческая биология. 31 (5): 559–573. Дои:10.1080/08912963.2017.1377194. S2CID 135302585.
- ^ Фройденталь, М. (1985). Cricetidae (Rodentia) из неогена Гаргано (пров. Фоджа, Италия). Рейксмузеум ван геологии и минералогии.
- ^ «Изучены« сверхразмерные »полевые мыши Сент-Кильды». BBC. 2010-09-03. Получено 2020-03-02.
- ^ http://www.mapama.gob.es/es/biodiversidad/temas/conservacion-de-especies-amenazadas/LIRON_tcm7-20977.pdf
- ^ Timm, R.M .; Weijola, V .; Аплин, К. П .; Donnellan, S.C .; Flannery, T. F .; Thomson, V .; Пайн, Р. Х. (2016-04-12). "Новый вид Раттус (Rodentia: Muridae) с острова Манус, Папуа-Новая Гвинея ". Журнал маммологии. 97 (3): 861–878. Дои:10.1093 / jmammal / gyw034.
- ^ http://www.raco.cat/index.php/Orsis/article/viewFile/24434/24268
- ^ Daams, R .; Фройденталь, М. (1985). "Stertomys laticrestatus, новый глирид (сони, Rodentia) из островной фауны Гаргано (пров. Фоджа, Италия) ». Scripta Geologica. 77: 21–27.
- ^ а б MacPhee, R.D.E., Iturralde-Vinent, M.A., и Gaffney, E.S. (Февраль 2003 г.). «Домо де Заза, местонахождение позвоночных животных раннего миоцена на юге центральной части Кубы, с заметками о тектонической эволюции Пуэрто-Рико и проливе Мона». Американский музей Novitates. 3394 (1): 1–42. Дои:10.1206 / 0003-0082 (2003) 394 <0001: DDZAEM> 2.0.CO; 2. HDL:2246/2820.CS1 maint: несколько имен: список авторов (связь)
- ^ «Позднемеловые животные острова Хацег в Румынии - более сложный вид».
- ^ Уорти, Тревор Х .; и другие. (2013). Окаменелости миоцена показывают, что киви (Аптерикс, Apterygidae) вероятно не филетические карлики (PDF). Палеорнитологические исследования 2013, Труды 8-го Международного собрания Общества палеонтологии и эволюции птиц. Получено 16 сентября 2017.
- ^ Павия, М .; Meijer, H. J. M .; Росси, М. А .; Гёлих, У. Б. (11 января 2017 г.). "Крайняя островная адаптация Гарганорнис баллманни Meijer, 2014: гигантские Anseriformes неогена Средиземноморского бассейна ». Королевское общество открытой науки. 4 (1): 160722. Bibcode:2017RSOS .... 460722P. Дои:10.1098 / rsos.160722. ЧВК 5319340. PMID 28280574.
- ^ https://www.canterburymuseum.com/about-us/media-releases/african-origins-for-the-enigmatic-adzebill/
- ^ Нейш, Даррен (2008-01-28). «Титан-ястребы и другие супер-хищники». Блог о четвероногих зоологии. ScienceBlogs LLC. Получено 2011-03-02.
- ^ Lerner, Heather R.L .; Минделл, Дэвид П. (2005). «Филогения орлов, стервятников Старого Света и других Accipitridae на основе ядерной и митохондриальной ДНК». Молекулярная филогенетика и эволюция. 37 (2): 327–346. Дои:10.1016 / j.ympev.2005.04.010. PMID 15925523.
- ^ Pregill, G.K .; Стедман, Д. У. (март 2004 г.). «Игуаны южной части Тихого океана: воздействие человека и новый вид». Журнал герпетологии. 38 (1): 15–21. Дои:10.1670 / 73-03A. JSTOR 1566081. S2CID 85627049.
- ^ Pregill, G.K .; Уорти, Т. Х. (март 2003 г.). «Ящерица-игуанид (Squamata, Iguanidae) из четвертичного периода Лар на Фиджи, юго-западная часть Тихого океана». Herpetologica. 59 (1): 57–67. Дои:10.1655 / 0018-0831 (2003) 059 [0057: ANILSI] 2.0.CO; 2. ISSN 0018-0831.
- ^ а б Петрен, К .; Кейс, T.J. (1997). "Филогенетический анализ эволюции размеров тела и биогеография в Чаквалласе (Савромалус) и другие игуанины ». Эволюция. 51 (1): 206–219. Дои:10.1111 / j.1558-5646.1997.tb02402.x. PMID 28568786. S2CID 22032248.
- ^ Уилсон, К.-Дж. (2004). Полет гуйя: экология и сохранение новозеландских лягушек, рептилий, птиц и млекопитающих. Издательство Кентерберийского университета. ISBN 0-908812-52-3. OCLC 937349394.
- ^ а б c Стюарт, К. "Диплодактиль Гекконы Австралии ». reptilesmagazine.com/. Получено 2020-02-29.
- ^ а б Bergman, J .; Хэмпер, Р. (2016). «Лист по уходу за гигантским гекконом Новой Каледонии». reptilesmagazine.com/. Получено 2020-02-29.
- ^ Maca-Meyer, N .; Carranza, S .; Rando, J.C .; Arnold, E.N .; Кабрера, В. М. (01.12.2003). "Статус и взаимоотношения вымершей гигантской канарской ящерицы Галлотия голиаф (Reptilia: Lacertidae), оцененная с использованием древней мтДНК из его мумифицированных останков " (PDF). Биологический журнал Линнеевского общества. 80 (4): 659–670. Дои:10.1111 / j.1095-8312.2003.00265.x. Получено 2010-04-03.
- ^ https://www.jstage.jst.go.jp/article/hsj2000/21/1/21_1_43/_pdf
- ^ Keogh, J. S .; Scott, I. A. W .; Хейс, К. (январь 2005 г.). «Быстрое и повторяющееся происхождение островного гигантизма и карликовости у австралийских тигровых змей». Эволюция. 59 (1): 226–233. Дои:10.1111 / j.0014-3820.2005.tb00909.x. PMID 15792242. S2CID 58524.
- ^ Майкл, Д .; Clemann, N .; Робертсон, П. (2018). "Notechis scutatus". Красный список видов, находящихся под угрозой исчезновения МСОП. 2018: e.T169687A83767147. Получено 19 декабря 2019.
- ^ а б Hocknull, S.A .; Piper, P.J .; van den Bergh, G.D .; Due, R.A .; Морвуд, M.J .; Курниаван, И. (2009). «Потерянный рай для драконов: палеобиогеография, эволюция и исчезновение крупнейших наземных ящериц (Varanidae)». PLOS ONE. 4 (9): e7241. Bibcode:2009PLoSO ... 4,7241H. Дои:10.1371 / journal.pone.0007241. ЧВК 2748693. PMID 19789642.
- ^ а б Hansen, D. M .; Donlan, C.J .; Griffiths, C.J .; Кэмпбелл, К. Дж. (Апрель 2010 г.). «Экологическая история и скрытый потенциал сохранения: большие и гигантские черепахи как модель для замены таксонов» (PDF). Экография. 33 (2): 272–284. Дои:10.1111 / j.1600-0587.2010.06305.x. Получено 2012-03-02.
- ^ Cione, A. L .; Tonni, E. P .; Сойбелзон, Л. (2003). "Сломанный зигзаг: вымирание крупных млекопитающих и черепах в позднем кайнозое в Южной Америке" (PDF). Rev. Mus. Argentino Cienc. Нат. Н.С. 5 (1): 1–19. Дои:10.22179 / REVMACN.5.26. ISSN 1514-5158. Архивировано из оригинал (PDF) на 2011-07-06. Получено 2011-02-06.
- ^ Фаринья, Р.А., Вискайно, С.Ф. И Де Юлиис, Г. (2013) Мегафауна: гигантские звери Южной Америки. Издательство Индианского университета, 448 страниц.
- ^ а б Харрисон, Т. (2011). «Черепахи (Chelonii, Testudinidae)». Палеонтология и геология Лаэтоли: эволюция человека в контексте, Vol. 2: Ископаемые гоминины и связанная с ними фауна. Палеобиология и палеоантропология позвоночных. Springer Science + Business Media. С. 479–503. Дои:10.1007/978-90-481-9962-4_17. ISBN 978-90-481-9961-7.
- ^ а б c Measey, G.J .; Vences, M .; Drewes, R.C .; Chiari, Y .; Мело, М .; Бурлес, Б. (2006). «Пресноводные тропы через океан: молекулярная филогения лягушки. Ptychadena newtoni дает представление о колонизации океанических островов земноводными ". Журнал биогеографии. 34 (1): 7–20. Дои:10.1111 / j.1365-2699.2006.01589.x.
- ^ "Лептопелис пальматус". amphibiaweb.org. Калифорнийский университет в Беркли. 2008 г.. Получено 29 февраля 2020.
- ^ "Leptopelis rufus". amphibiaweb.org. Калифорнийский университет в Беркли. 2008 г.. Получено 29 февраля 2020.
- ^ Уорти, Т. (2001). "Новый вид Платимантида (Anura: Ranidae) из четвертичных отложений на Вити-Леву, Фиджи ". Палеонтология. 44 (4): 665–680. Дои:10.1111/1475-4983.00197.
- ^ Ни кокосовые крабы, ни их родственники не могут плавать дальше стадии личинки, поэтому взрослые особи практически высаживаются на землю. Кокосовые крабы могут весить более 4 кг (9 фунтов); самые большие раки-отшельники родственного рода Coenobita, C. brevimanus прибрежных районов Африки и Азии, всего 230 граммов (0,5 фунта).
- ^ «План действий по сохранению биоразнообразия острова Вознесения: Гарип титаниус план действий по видам " (PDF). Джорджтаун, остров Вознесения: Государственный департамент охраны природы острова Вознесения. 2015-02-26. Получено 2019-09-11.
- ^ Кеппель, Гуннар; Лоу, Эндрю Дж .; Поссингем, Хью П. (2009). «Меняющиеся взгляды на биогеографию тропической части южной части Тихого океана: влияние распространения, викариантности и исчезновения». Журнал биогеографии. 36 (6): 1035–1054. Дои:10.1111 / j.1365-2699.2009.02095.x. ISSN 0305-0270.
- ^ Боуэн, Лизабет; Вурен, Дирк Ван (1997). «Островные эндемичные растения не обладают защитой от травоядных животных». Биология сохранения. 11 (5): 1249–1254. Дои:10.1046 / j.1523-1739.1997.96368.x. ISSN 0888-8892.
- ^ а б c https://www.sciencelearn.org.nz/resources/2289-small-islands-breed-big-seeds
- ^ http://www.terrain.net.nz/friends-of-te-henui-group/table-1/corokia-macrocarpa-chatham-is-korokio.html
- ^ http://www.terrain.net.nz/friends-of-te-henui-group/table-1/korokio.html
- ^ а б Бернс, К. (Май 2019). Эволюция в изоляции: поиск островного синдрома у растений. Издательство Кембриджского университета. Дои:10.1017/9781108379953. ISBN 978-1108379953. OCLC 1105218367.
- ^ Проктор, Дж. (1984). «Растительность гранитных островов Сейшельских островов». В Стоддарте, Д. Р. (ред.). Биогеография и экология Сейшельских островов. W. Junk. ISBN 978-90-6193-881-1. OCLC 906429733.
внешняя ссылка
| Правила |
|  |
|---|---|---|
| Связанный |
| |