Transandinomys bolivaris - Transandinomys bolivaris
| Transandinomys bolivaris | |
|---|---|
 | |
| Череп с Серро Азул, Панама, вид сверху[1] | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Rodentia |
| Семья: | Cricetidae |
| Подсемейство: | Sigmodontinae |
| Род: | Трансандиномис |
| Разновидность: | Т. bolivaris |
| Биномиальное имя | |
| Transandinomys bolivaris (J.A. Аллен, 1901) | |
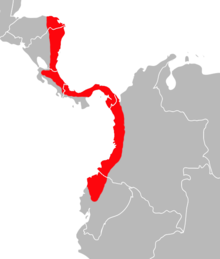 | |
| Распределение Transandinomys bolivaris в южной части Центральной и северо-западной части Южной Америки[3] | |
| Синонимы[11] | |
| |

Transandinomys bolivaris, также известный как рисовая крыса с длинными усами, это грызун в роду Трансандиномис. Он встречается во влажных лесах от северо-востока Гондураса до западного Эквадора, на высоте до 1800 м (5900 футов) над уровнем моря. С тех пор как он был впервые описан в 1901 году в Эквадоре, для него было введено шесть научных названий, но их общая идентичность не была задокументирована до 1998 года, и этот вид долгое время был известен под названием Oryzomys bombycinus, описан из Панамы в 1912 году. Название Оризомыс боливарис использовался до того, как его перевели в новый род Трансандиномис с Transandinomys talamancae (ранее Oryzomys talamancae) в 2006 году.
Это среднего размера рисовая крыса отличается очень длинным вибриссы (усы) - те, что над глазами, до 50 мм (2 дюйма) в длину. Мех мягкий и густой, обычно темно-коричневый сверху и светло-серый снизу; у молоди темнее. Ноги длинные, а длина хвоста равна длине головы и тела. Череп узкий и имеет широкий межглазничная область (между глазами). Вид обычно живет на земле. Хотя это редко, его статус сохранения считается безопасным.
Таксономия
В 1901 г. Джоэл Асаф Аллен описал четыре новых вида рисовых крыс этого рода Оризомыс: трое из Эквадора и перуанца Oryzomys perenensis. Три эквадорских вида -Оризомыс боливарис из Порвенира,[Примечание 1] Боливарская провинция; Oryzomys castaneus из Сан-Хавьера, Провинция Эсмеральдас; и Oryzomys rivularis из Рио Верде, Провинция Пичинча - каждый основан на одном экземпляре, собранном в 1899 или 1900 гг.[13] Он выделил троих по окраске, размеру и относительной длине хвоста.[14] Филип Гершковиц перечислил все три среди многих синонимы из "Оризомыс латицепс "(в настоящее время более узко определяется как Hylaeamys laticeps ) в статье 1960 года.[15]
Эдвард Альфонсо Гольдман описанный Oryzomys bombycinus в 1912 г. из четырех экземпляров из Панамы. Он сравнил это с Oryzomys talamancae и поместил его с "Оризомыс латицепс группа".[16] Три года спустя он описал Oryzomys nitidus alleni из Коста-Рики как подвид из Oryzomys nitidus, не говоря уже о бомбицин.[8] Он пересмотрел Оризомыс Северной Америки в 1918 г. Oryzomys bombycinus как единственный член своей группы, с Аллени как подвид, отличающийся пропорциями черепа. Он также упомянул, что группа произошла в Эквадоре, и указал, что O. bombycinus вероятно добрался до Колумбии. Голдман считал группу похожей на О. таламанка, но предположил, что бомбицин и Аллени может быть только подвидом О. nitidus.[17] В 1939 году Оливер Пирсон добавил третий подвид, О. Б. оринус, из восточной Панамы,[18] а в 1966 году этот вид был впервые зарегистрирован в Колумбии.[19] Рональд Пайн отзывался Oryzomys bombycinus в 1971 году, когда было известно 59 его экземпляров, и впервые был зарегистрирован вид из Никарагуа и Эквадора.[20] Он сохранил три описанных подвида -Аллени от Никарагуа до западной Панамы, бомбицин из центральной Панамы и оринус из восточной Панамы в Эквадор.[21]
Альфред Гарднер и Джеймс Паттон предположили в 1976 году, что О. rivularis может быть того же вида, что и O. bombycinus. Они считали О. боливарис так же вероятно, как О. nitidus и перечисленные Castaneus как синоним О. капито (эквивалент современного Hylaeamys megacephalus и близкородственные виды плюс Transandinomys talamancae).[22] В 1984 году Беншоф и его коллеги сообщили о первой записи Oryzomys bombycinus из Гондураса.[23] Гай Мюссер и Марина Уильямс рассмотрели О. таламанка в 1985 и включены О. castaneus как один из его синонимов, хотя и не изучив голотип.[24] Во втором издании 1993 г. Виды млекопитающих мира, Мюссер и Майкл Карлтон использовали имя Оризомыс боливарис для видов, ранее известных как O. bombycinus, а в 1998 году Мюссер и его коллеги полностью задокументировали присвоение имен Боливарис, Castaneus, rivularis, бомбицин, Аллени, и оринус к тому же виду, Оризомыс боливарис.[25] Они отметили его сходство с О. таламанка, но не пытался определить филогенетический отношения между видами, которые они обсуждали.[26] В своем ограниченном материале они обнаружили, что географические вариации внутри вида несущественны, и они не признали никаких подвидов.[27]
В 2006 году Марсело Векслер опубликовал филогенетический анализ Oryzomyini («рисовые крысы»), племя которому Оризомыс выделяется, используя морфологический и Последовательность ДНК данные. Его результаты показали виды Оризомыс рассредоточены по Oryzomyini и предположили, что большинство видов этого рода следует отнести к новым родам.[28] Позже в том же году он вместе с Александром Персекильо и Робертом Воссом назвал десять новых родов этих видов, включая Трансандиномис, у которого есть Oryzomys talamancae (сейчас же Transandinomys talamancae) как его типовой вид.[10] Они также включали Оризомыс боливарис в Трансандиномис, так что теперь он называется Transandinomys bolivaris, хотя он не был включен в филогенетическое исследование Векслера. Два вида морфологически похожи, но они смогли идентифицировать только один синапоморфия (общая-производная черта) для них: очень долго надбровные вибриссы (усы над глазами).[29] Трансандиномис является одним из примерно 30 родов Oryzomyini, разнообразного сообщества американских грызунов, насчитывающего более ста видов,[30] и на более высоких таксономических уровнях в подсемействе Sigmodontinae семьи Cricetidae, наряду с сотнями других видов, в основном мелких грызунов.[31]
Различные авторы использовали распространенное имя «длинноусая рисовая крыса» для этого вида,[32] но было предложено несколько других названий. В 1918 году Гольдман назвал О. bombycinus bombycinus «Длинношерстная рисовая крыса»[33] и О. Б. Аллени "Аллен Райс Рэт".[34] Массер и Карлтон в третьем издании 2005 г. Виды млекопитающих мира, б / у "Орызомыс длинноусый",[35] Красный список МСОП 2009 г. поставил "Рисовую Крысу Боливар",[2] и Томас Ли и его коллеги использовали «длинноусую трансандскую рисовую крысу» в 2010 году.[36]
Описание
| Область, край | п[Заметка 2] | Голова и тело | Хвост | Задняя лапа |
|---|---|---|---|---|
| Никарагуа | 5 | 108.2 (99–119) | 115.2 (99–126) | 28.8 (27–32) |
| Коста-Рика, западная Панама | 9–13 | 118.7 (103–134) | 113.2 (96–125) | 30.3 (27–33) |
| Центральная Панама | 13–15 | 120.7 (111–140) | 120.8 (111–130) | 30.0 (27–33) |
| Восточная Панама | 5 | 116.8 (107–125) | 117.6 (109–126) | 28.0 (25–29) |
| Колумбия | 8 | 127.4 (115–141) | 121.3 (100–135) | 30.5 (29–32) |
| Эквадор | 8–9 | 119.4 (111–133) | 114.8 (100–127) | 29.7 (28–32) |
| Размеры даны в миллиметрах и в форме «среднее (минимум – максимум)». | ||||
Transandinomys bolivaris Рисовая крыса среднего размера с очень длинными надбровными вибриссами.[Заметка 3] (более 50 мм (2,0 дюйма) в длину и выходит далеко за уши, когда кладется спиной к голове).[38] По словам Фионы Рид Млекопитающие Юго-Восточной Мексики и Центральной Америки, его можно отличить от любых рисовых крыс аналогичного размера по длине этих усов;[39] T. talamancae также имеет длинные надбровные вибриссы, но не такие длинные, как у Т. bolivaris.[10] У обоих видов мистические вибриссы (над ртом) также длинные и выходят за уши, когда они отведены назад, но они снова намного длиннее в Т. bolivaris.[10] Вибриссы в основном темные, но на концах полупрозрачные.[40] Handleyomys alfaroi, рисовая крыса, с которой молодые Т. bolivaris часто путают,[41] намного меньше.[42] В Т. bolivaris, щеки могут быть светло-серыми, бафф, или красноватый. Уши от темно-коричневого до черного, с редкой опушкой.[43]

Мех мягкий, плотный и густой,[40] от темно-коричневого до серого на верхней части тела, переходящего в черный в средней части спины и желтовато-коричневый по бокам.[38] Нижняя часть тела резко отличается по окраске. Здесь волосы темно-серые у основания и белые на кончиках, так что мех кажется серовато-белым.[40] Мех короче и темнее, чем у T. talamancae[44] и мягче и толще, чем в Х. альфарои.[42] У молодых животных шерсть темнее, тоньше и мягче.[45] Сосна разделила подвиды Аллени и оринус на основе их более темного меха,[46] но Муссер и его коллеги не смогли подтвердить эту закономерность и нашли более светлые и темные экземпляры в тех же географических регионах.[47]
Хвост кажется голым[39] и короче или примерно такой же длины, как голова и тело;[48] это дольше в T. talamancae.[49] Его окраска изменчива; он темно-коричневый сверху и по бокам и светло-коричневый и часто белый в большей или меньшей степени снизу, а у некоторых экземпляров хвост имеет одинаковый цвет сверху и снизу.[Примечание 4] Чешуя на хвосте мельче, чем у T. talamancae.[51]
Предплечья темно-серые. Передние лапы не пигментированы и ногтевые пучки белых волосков окружает столь же непигментированные когти.[45] У животного длинные узкие задние лапы,[39] дольше, чем в T. talamancae,[49] с обычно гладкой подошвой (отсутствует чешуйки, которые присутствуют в T. talamancae).[Примечание 5] Три средние цифры намного длиннее двух внешних. Шесть колодки присутствуют на подошве. Верхняя поверхность и бока задних лап белые и кажутся голыми, хотя присутствуют короткие белые волоски;[45] эти волосы длиннее в T. talamancae.[52] Когтевые пучки длинных, белых или седых волос вокруг когтей.[53] которые короткие и лишены пигмента.[45]
Длина головы и тела от 100 до 140 мм (от 3,9 до 5,5 дюйма), длина хвоста от 90 до 130 мм (от 3,5 до 5,1 дюйма), длина задней части стопы от 27 до 35 мм (от 1,1 до 1,4 дюйма), длина ушей от 16 до 21 мм (0,63 дюйма). до 0,83 дюйма), а масса тела от 39 до 75 г (от 1,4 до 2,6 унции).[38] У самок четыре пары молочные железы, как обычно у oryzomyines.[45] Как и большинство рисовых крыс, Т. bolivaris имеет двенадцать грудной (сундук) и семь поясничный позвонки.[54] Исследование, проведенное в Коста-Рике, показало, что существует 58 хромосомы, в том числе многие большие с двумя руками, а основное число рук - 80 (2n = 58, FN = 80), высокодифференцированный кариотип.[41] Кариотип T. talamancae вариабельна, но имеет меньше хромосом (от 34 до 54) и основных плеч (от 60 до 67).[55] Х. альфарои имеет больше хромосом (от 60 до 62) и основных плеч (от 100 до 104).[42]
Череп и зубы


Череп относительно длинный с длинным узким рострумом (передняя часть), широким межглазничная область (между глазами) и узкий черепная коробка с почти вертикально ориентированными стенами по бокам и сзади.[45] Он отличается от T. talamancae в различных пропорциях.[52] Хотя подвид, ранее признанный в "Oryzomys bombycinus«были разделены небольшими различиями в чертах черепа, Пайн отверг их на основании своих гораздо более крупных образцов.[56] Мюссер и его коллеги согласились, но отметили, что у колумбийских животных черепа были больше.[57] В скуловые пластины широкие и скуловые дуги (скулы) позади них почти параллельны друг другу. По краям межглазничной области видны выступающие бусинки, которые доходят до мозга в виде височные гребни;[45] они обычно менее развиты в T. talamancae.[58] В отличие от T. talamancae и Х. альфарои, то теменная кость обычно ограничивается крышей мозговой коробки и не распространяется в стороны.[59]
В резкое отверстие, которые перфорируют переднюю часть нёбо, не проходите между коренные зубы. Небо заканчивается за третьими коренными зубами и перфорировано заднебоковые небные ямки там. За ним крыша мезоптеригоидная ямка перфорирован плохо развитым клиновидно-небные пустоты. В слуховые буллы, в которых находится внутреннее ухо, большие.[45] Обычно сосцевидная кость отсутствуют отверстия (фенестры), которые присутствуют в T. talamancae.[10] Образец артерии в голове примитивен, на что указывает состояние различных отверстие (отверстия) и бороздки в черепе.[41]

В нижняя челюсть (нижняя челюсть) выглядит коренастой и имеет длинный кондиллоидный отросток за спиной;[41] что из T. talamancae более стройный.[61] В капсульный отросток, выступ в задней части челюсти, в котором находится корень нижнего резец, развит слабо.[41]
Резцы большие, без бороздок. Их эмаль оранжевый, но бледнее на нижних. Ориентация верхних резцов опистодонт, с режущей кромкой, обращенной назад. Моляры Brachydont (с низкой коронкой) и имеют два ряда главных выступов, разделенных глубокими впадинами и дополненных сетью гребней и меньших выступов.[41] Первый верхний моляр шире, чем у T. talamancae.[61] Как у этого вида, но в отличие от многих других рисовых крыс, в том числе Х. альфарои, то мезофлексия на втором верхнем моляре, который разделяет паракон (один из главных куспидов) от мезолоф (вспомогательный гребень), не разделяется эмалевым мостом надвое.[62] В гипофлексид на втором нижнем моляре основная впадина между бугорками очень длинная, простирается более чем на половину длины зуба; по этому признаку он похож на T. talamancae но в отличие от Х. альфарои.[42] У каждого из верхних коренных зубов по три корня (два на внешней стороне и один на внутренней), а на каждом из нижних - по два (один спереди, а другой сзади).[45]
Распространение, экология и поведение

Transandinomys bolivaris - необычный вид.[39] Его известное распространение простирается от северо-востока Гондураса, на побережье Карибского моря, через восточную часть Никарагуа, Коста-Рику и Панаму до прибрежных западных районов Колумбии и северо-западных и западно-центральных районов Эквадора. Он был обнаружен около уровня моря, а высотные рекорды находятся на высоте почти 1500 м (4900 футов) в Панаме и 1800 м (5900 футов) в Эквадоре. Это распределение совпадает с распределением влажных Трансандийский лес.[64] Обычно это происходит в районах со средней температурой выше 16 ° C (60,8 ° F) и годовым количеством осадков от 4000 до 6000 мм (от 157 до 236 дюймов),[65] предпочитает леса на средней высоте (от 600 до 900 м или от 1970 до 2950 футов) и часто встречается у воды.[39] Можно ожидать, что фактический ареал этого вида расширится дальше на север и запад, возможно, в Веракрус, южная Мексика и западная Венесуэла, где он еще не зарегистрирован.[66] Омар Линарес упомянул возможную запись из Озеро Маракайбо регион на северо-западе Венесуэлы в 1998 году.[67] Его диапазон аналогичен диапазону различных других животных тропических лесов, включая полупустой ястреб (Leucopternis semiplumbeus), рисовые крысы Sigmodontomys alfari и С. афрастус, колючие крысы Proechimys semispinosus и Hoplomys gymnurus,[68] и опоссум Мармоса зеледони.[69] T. talamancae и Х. альфарои часто встречаются в тех же местах, что и Т. bolivaris, но встречаются и в других областях.[41]
Мало что известно о его биологии.[70] Вид в основном обитает на земле, но некоторые молодые животные были взятый в растительности на высоте до 1,5 м (4,9 фута) над землей. Обычно его ловят «под бревнами, у корней больших деревьев или среди камней у ручьев».[39] Две самки с четырьмя эмбрионами каждая были пойманы в Панаме в июне, одна с двумя - в Никарагуа в сентябре.[71] и один с четырьмя в Коста-Рике в декабре.[72] В марте в Коста-Рике был пойман очень молодой экземпляр. Одна беременная самка все еще была в ювенильной шерсти.[71] Четыре вида клещи были найдены на Т. bolivaris в Панаме (Gigantolaelaps gilmorei, G. oudemansi, Лаэлапс пилифер, и Haemolaelaps glasgowi ),[73] два чиггеры (Leptotrombidium panamensis и Pseudoschoengastia bulbifera ),[74] и два блохи (Polygenis roberti и Polygenis klagesi ).[75]
Статус сохранения
2009 год Красный список МСОП списки Т. bolivaris в качестве "Наименьшие опасения ", поскольку это широко распространенный вид с предположительно большой популяцией, которая встречается во многих охраняемые территории. Тем не мение, разрушение среды обитания обезлесение может представлять угрозу.[2]
Примечания
- ^ В 1998 году Мюссер и его коллеги не смогли определить точное местонахождение этого места; см. там для обсуждения.[12]
- ^ Количество измеренных образцов. В некоторых образцах количество зависит от измерения.
- ^ См. Musser et al. (1998, рис. 53) и Рид (2009, табл. 23, рис. 3) для иллюстраций.
- ^ Это описание основано на подробном описании окраски хвоста Массером и его коллегами.[40] Другие описания меняются: Рид пишет, что хвост двухцветный,[39] но Векслер, Персекильо и Фосс утверждают, что он одноцветный[10] и Тирира, что он одноцветный или немного бледнее на нижней стороне у основания.[50]
- ^ Векслер и коллеги[10] и Тирира[50] пишут, что подошва гладкая, но Мюссер и его коллеги упоминают экземпляры, у которых действительно есть рудиментарные чешуи между пальцами.[45]
Рекомендации
- ^ а б c Гольдман, 1918 г., тарелка II
- ^ а б c Гомес-Лаверде и др., 2008 г.
- ^ Musser et al., 1998, рис. 50
- ^ Аллен, 1901, стр. 405
- ^ Аллен, 1901, стр. 406
- ^ Аллен, 1901, стр. 407
- ^ Гольдман, 1912, с. 6
- ^ а б Гольдман, 1912, с. 129
- ^ Пирсон, 1939, стр. 2
- ^ а б c d е ж грамм Weksler et al., 2006, стр. 25
- ^ Musser et al., 1998, стр. 271
- ^ Musser et al., 1998, стр. 272
- ^ Аллен, 1901, стр. 405–407; Musser et al., 1998, стр. 271
- ^ Musser et al., 1998, стр. 273
- ^ Гершковиц, 1960, стр. 544
- ^ Гольдман, 1912, стр. 6–7.
- ^ Гольдман, 1918, с. 75–78.
- ^ Пирсон, 1939, стр. 2–3.
- ^ Сосна, 1971, с. 592
- ^ Сосна, 1971, с. 590
- ^ Сосна, 1971 г., рис. 1
- ^ Гарднер и Паттон, 1976, стр. 39–41.
- ^ Benshoof et al., 1984, стр. 512
- ^ Массер и Уильямс, 1985, стр. 14
- ^ Musser et al., 1998, стр. 110, 273
- ^ Musser et al., 1998, стр. 323
- ^ Musser et al., 1998, стр. 148–149.
- ^ Weksler, 2006, стр. 75–77.
- ^ Weksler et al., 2006, стр. 26
- ^ Векслер, 2006, с. 3
- ^ Мюссер и Карлтон, 2005
- ^ Сосна, 1971, с. 570; Рид, 2009, стр. 208
- ^ Гольдман, 1918, с. 77
- ^ Гольдман, 1918, с. 78
- ^ Массер и Карлтон, 2005, стр. 1146
- ^ Ли и др., 2010, стр. 10
- ^ Musser et al., 1998, таблица 25
- ^ а б c Тирира, 2007, с. 199; Рид, 2009, стр. 208
- ^ а б c d е ж грамм Рид, 2009, стр. 208
- ^ а б c d Musser et al., 1998, стр. 123
- ^ а б c d е ж грамм Musser et al., 1998, стр. 125
- ^ а б c d Musser et al., 1998, стр. 143
- ^ Musser et al., 1998, стр. 121
- ^ Musser et al., 1998, стр. 128
- ^ а б c d е ж грамм час я j Musser et al., 1998, стр. 124
- ^ Musser et al., 1998, стр. 145–146.
- ^ Musser et al., 1998, стр. 148
- ^ Musser et al., 1998, стр. 121; Тирира, 2007, с. 199
- ^ а б Musser et al., 1998, стр. 427
- ^ а б Тирира, 2007, с. 199
- ^ Musser et al., 1998, стр. 129
- ^ а б Musser et al., 1998, стр. 131
- ^ Musser et al., 1998, стр. 124; Рид, 2009, стр. 208
- ^ Степпан, 1995 г., стол 5
- ^ Musser et al., 1998, таблица 13
- ^ Musser et al., 1998, стр. 145
- ^ Musser et al., 1998, стр. 146
- ^ Musser et al., 1998, стр. 135
- ^ Musser et al., 1998, стр. 135, 143; Weksler et al., 2006, стр. 25
- ^ Гольдман, 1918 г., тарелка V
- ^ а б Musser et al., 1998, стр. 140
- ^ Musser et al., 1998, с. 140, 143
- ^ Гольдман, 1918 г., пластина VI
- ^ Musser et al., 1998, стр. 113
- ^ Musser et al., 1998, стр. 116
- ^ Musser et al., 1998, стр. 119
- ^ Линарес, 1998, стр. 284
- ^ Musser et al., 1998, стр. 119, 121.
- ^ Росси и др., 2010, с. 65
- ^ Тирира, 2007, с. 200
- ^ а б Сосна, 1971, с. 595
- ^ Рид и Лэнгтимм, 1993, стр. 93
- ^ Типтон и др., 1966, стр. 42
- ^ Бреннан и Юнкер, 1966, стр. 266
- ^ Типтон и Мендес, 1966, стр. 323
Цитированная литература
- Аллен, Дж. 1901 г. Новые южноамериканские мюриды и новый Метахирус. Бюллетень Американского музея естественной истории 14: 405–412.
- Беншоф, Л., Йейтс, Т. и Froehlich, J.W. 1984 г. Заслуживающие внимания записи о млекопитающих из восточного Гондураса (требуется подписка). Юго-западный натуралист 29 (4): 511–514.
- Бреннан, Дж. М., Юнкер, К. Э. 1966. Чиггеры Панамы (Acarina: Trombiculidae). Стр. 221–266 в Wenzel, R.L. and Tipton, V.J. (ред.). Эктопаразиты Панамы. Чикаго: Полевой музей естественной истории.
- Дафф, А. и Лоусон, А. 2004. Млекопитающие мира: Контрольный список. Нью-Хейвен: A&C Black. ISBN 0-7136-6021-X.
- Гарднер А.Л. и Паттон Дж. Л. 1976. Кариотипические вариации у оризоминых грызунов (Cricetinae) с комментариями по хромосомной эволюции в неотропическом крикетиновом комплексе. Случайные статьи из Зоологического музея, Университет штата Луизиана 49: 1–48.
- Гольдман, Э.А. 1912 г. Описание двенадцати новых видов и подвидов млекопитающих Панамы.. Разные коллекции Смитсоновского института 56 (№ 36): 1–11.
- Гольдман, Э.А. 1915 г. Пять новых рисовых крыс этого рода Оризомыс из Средней Америки. Труды Биологического общества Вашингтона 28: 127–130.
- Гольдман, Э.А. 1918 г. Рисовые крысы Северной Америки. Фауна Северной Америки 43: 1–100.
- Гомес-Лаверде, М., Ривас, Б., Тимм, Р. и Пино, Дж. 2008. Transandinomys bolivaris. В МСОП. Красный список исчезающих видов МСОП. Версия 2009.2. <www.iucnredlist.org >. Загружено 4 мая 2010 г.
- Гершковиц, П. 1960 г. Млекопитающие северной Колумбии, предварительный отчет нет. 8. Древесные рисовые крысы, систематический пересмотр подрода. Oecomys, род Оризомыс. Труды Национального музея США 110: 513–568.
- Ли, Т.Э., младший, Бурнео, С.Ф., Кокран, Т.Дж. и Чавес, Д. 2010. Мелкие млекопитающие Санта-Росы, юго-западная провинция Имбабура, Эквадор. Случайные статьи, Музей Техасского технического университета 290: 1–14.
- Линарес, О. 1998. Mamíferos de Venezuela. Каракас: Sociedad Conservacionista Audubon de Venezuela и British Petroleum, 691 стр. (На испанском языке). ISBN 980-6326-16-4
- Musser, G.G. и Карлтон, доктор медицины, 2005. Надсемейство Muroidea. Стр. 894–1531 в Wilson, D.E. и Ридер, Д. (ред.). Виды млекопитающих мира: таксономический и географический справочник. 3-е изд. Балтимор: Издательство Университета Джона Хопкинса, 2 тома, 2142 стр.ISBN 978-0-8018-8221-0
- Musser, G.G. и Уильямс, М. 1985 г. Систематические исследования грызунов оризомиинов (Muridae): определения Oryzomys villosus и Oryzomys talamancae. Американский музей Novitates 2810: 1–22.
- Массер, Г.Г., Карлтон, доктор медицины, братья Э.М. и Гарднер, А.Л. 1998. Систематические исследования грызунов оризомиинов (Muridae: Sigmodontinae): диагностика и распространение видов, ранее отнесенных к Оризомыс "капито". Бюллетень Американского музея естественной истории 236: 1–376.
- Пирсон, О. П. 1939. Три новых мелких млекопитающих из восточной Панамы. Notulae Naturae, Академия естественных наук (Филадельфия) 6: 1–5.
- Пайн Р.Х. 1971. Обзор рисовой крысы с длинными усами, Oryzomys bombycinus Goldman (требуется подписка). Journal of Mammalogy 52: 590–596.
- Рейд, Ф.А. (2009). Полевой справочник по млекопитающим Центральной Америки и Юго-Восточной Мексики (2-е изд.). Oxford University Press. ISBN 978-0-19-534322-9. OCLC 312626014.
- Рейд Ф.А. и Лангтимм К.А. 1993 г. Заметки о распространении и естественной истории избранных млекопитающих из Коста-Рики (требуется подписка). Юго-западный натуралист 38: 299–302.
- Росси Р.В., Восс Р.С. и Лунде, Д. 2010 г. Ревизия рода сумчатых дидельфид Мармоса. Часть 1. Виды в разделах Тейта «mexicana» и «mitis» и другие близкородственные формы.. Бюллетень Американского музея естественной истории 334: 1–83.
- Степпан, С.Дж. 1995 г. Ревизия трибы Phyllotini (Rodentia: Sigmodontinae) с филогенетической гипотезой для Sigmodontinae. Fieldiana Zoology 80: 1–112.
- Типтон, В.Дж. and Méndez, E. 1966. Блохи (Siphonaptera) Панамы. Стр. 289–385 в Wenzel, R.L. and Tipton, V.J. (ред.). Эктопаразиты Панамы. Чикаго: Полевой музей естественной истории.
- Типтон, В.Дж., Альтман, Р.М. и Кинан, К. 1966 г. Клещи подсемейства Laelaptinae в Панаме (Acarina: Laelaptidae). Стр. 23–82 в Wenzel, R.L. and Tipton, V.J. (ред.). Эктопаразиты Панамы. Чикаго: Полевой музей естественной истории.
- Тирира, Д. 2007. Guia de campo de los mamíferos del Ecuador. Кито: Ediciones Murciélago Blanco, publicación especial sobre los mamíferos del Ecuador 6, 576 стр. (На испанском языке). ISBN 9978-44-651-6
- Векслер М. 2006. Филогенетические взаимоотношения грызунов оризомиинов (Muroidea: Sigmodontinae): отдельный и комбинированный анализ морфологических и молекулярных данных. Бюллетень Американского музея естественной истории 296: 1–149.
- Векслер М., Персекильо А. и Восс, Р. 2006 г. Десять новых родов грызунов оризомиинов (Cricetidae: Sigmodontinae). Американский музей Novitates 3537: 1–29.