Цитотоксические Т-клетки - Cytotoxic T cell

А цитотоксические Т-клетки (также известен как ТC, цитотоксический Т-лимфоцит, CTL, Т-киллерная клетка, цитолитическая Т-клетка, CD8 + Т-клетки или же убийца Т-клетки) это Т-лимфоцит (тип лейкоцит ) что убивает рак клетки, клетки, которые инфицированы (особенно с вирусы ) или клетки, поврежденные иным образом.[1]
Большинство цитотоксических Т-клеток экспрессируют Рецепторы Т-клеток (TCR), которые могут распознать конкретный антиген. Антиген - это молекула, способная стимулировать иммунная реакция и часто производится раковые клетки или вирусы. Антигены внутри клетки связаны с класс I MHC молекулы MHC класса I и переносятся на поверхность клетки, где они могут быть распознаны Т-клеткой. Если TCR специфичен для этого антигена, он связывается с комплексом молекулы MHC класса I и антигена, и Т-клетка разрушает клетку.
Чтобы TCR мог связываться с молекулой MHC класса I, первая должна сопровождаться гликопротеин называется CD8, который связывается с постоянной частью молекулы MHC класса I. Поэтому эти Т-клетки называются CD8 + Т-клетки.
В близость между CD8 и молекулой MHC сохраняет TC клетка и клетка-мишень тесно связаны вместе во время антиген-специфической активации. CD8 + Т-клетки распознаются как ТC клетки, как только они активируются, и обычно классифицируются как имеющие заранее определенную цитотоксическую роль в иммунной системе. Однако CD8 + Т-клетки также обладают способностью производить некоторые цитокины.
Разработка
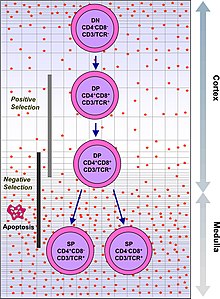
Иммунная система должна распознавать миллионы потенциальных антигенов. В организме человека менее 30 000 генов, поэтому невозможно иметь один ген для каждого антигена. Вместо этого ДНК в миллионах лейкоцитов в костном мозге перетасовывается, чтобы создать клетки с уникальными рецепторами, каждый из которых может связываться с разными антигенами. Некоторые рецепторы связываются с тканями в самом организме человека, поэтому, чтобы организм не атаковал сам себя, эти самореактивные лейкоциты разрушаются во время дальнейшего развития в организме человека. вилочковая железа, в котором йод необходимо для его развития и деятельности.[2]
TCR состоит из двух частей, обычно альфа и бета цепи. (У некоторых TCR есть гамма- и дельта-цепочка.) Гематопоэтические стволовые клетки в Костный мозг мигрировать в вилочковая железа, где они проходят V (D) J рекомбинация своей бета-цепочки TCR ДНК для формирования развивающейся формы белка TCR, известного как пре-TCR. Если эта перестройка успешна, клетки затем перестраивают свою ДНК TCR альфа-цепи, чтобы создать функциональный комплекс альфа-бета TCR. Этот крайне вариабельный продукт генетической перестройки генов TCR помогает создавать миллионы различных Т-клеток с разными TCR, помогая иммунной системе организма реагировать практически на любые белок захватчика. Подавляющее большинство Т-клетки экспрессируют альфа-бета TCR (αβ Т-клетки), но некоторые Т-клетки в эпителиальных тканях (например, кишечник) экспрессируют гамма-дельта TCR (гамма-дельта Т-клетки ), которые распознают небелковые антигены.
Т-клетки с функционально стабильными TCR экспрессируют как CD4 и CD8 корецепторы и поэтому называются «дважды положительными» (DP) Т-клетками (CD4 + CD8 +). Двойные положительные Т-клетки подвергаются воздействию широкого спектра аутоантигенов в тимусе и проходят два критерия отбора:
- положительный выбор, в котором те дважды положительные Т-клетки, которые связываются с чужеродным антигеном в присутствии собственного MHC. Они будут дифференцироваться на CD4 + или CD8 + в зависимости от того, какой MHC связан с представленным антигеном (MHC1 для CD8, MHC2 для CD4). В этом случае в клетках был бы представлен антиген в контексте MHC1. Положительный отбор означает отбор тех TCR, способных распознавать собственные молекулы MHC.
- отрицательный выбор, в котором те дважды положительные Т-клетки, которые связывают слишком сильно к MHC -представлено себя антигены пройти апоптоз потому что в противном случае они могут стать самореактивными, что приведет к аутоиммунитет.
Положительно отбираются только те Т-клетки, которые слабо связываются с комплексами МНС-аутоантиген. Те клетки, которые выживают при положительном и отрицательном отборе, дифференцируются в одноположительные Т-клетки (CD4 + или CD8 +), в зависимости от того, распознает ли их TCR антиген, представленный MHC класса I (CD8), или MHC класс II -представленный антиген (CD4). Именно CD8 + Т-клетки созревают и становятся цитотоксические Т-клетки после их активации антигеном, ограниченным классом I.
Активация

За исключением некоторых типов клеток, таких как не-зародился клетки (в том числе эритроциты ), MHC класса I выражается всеми хозяин клетки. Когда эти клетки заражены вирус (или другой внутриклеточный возбудитель ) клетки разрушают чужеродные белки через обработка антигена. В результате образуются пептидные фрагменты, некоторые из которых представлены MHC Class I для Рецептор антигена Т-клеток (TCR) на CD8 + Т-клетках.
Активация цитотоксических Т-клеток зависит от нескольких одновременных взаимодействий между молекулами, экспрессируемыми на поверхности Т-клеток, и молекулами на поверхности антигенпрезентирующая клетка (БТР). Например, рассмотрим двухсигнальная модель для TC активация клеток.
| Сигнал | Т-клетка | APC | Описание |
| Первый сигнал | TCR | связанный с пептидом MHC класс I молекула | Есть второе взаимодействие между CD8 корецептор и молекула MHC класса I для стабилизации этого сигнала. |
| Второй сигнал | CD28 молекула на Т-клетке | либо CD80 или же CD86 (также называемые B7-1 и B7-2) | CD80 и CD86 известны как костимуляторы для активации Т-клеток. Этому второму сигналу можно помочь (или заменить) стимулированием TC клетка с цитокинами, высвобождаемыми из Т-хелперные клетки. |
Простая активация наивного CD8+ Т-клетки требуют взаимодействия с профессиональными антигенпрезентирующими клетками, в основном со зрелыми дендритные клетки. Для создания долговечных Т-клетки памяти и чтобы позволить повторяющуюся стимуляцию цитотоксических Т-клеток, дендритные клетки должны взаимодействовать с обоими активированными CD4+ хелперные Т-клетки и CD8+ Т-клетки.[3][4] Во время этого процесса CD4+ хелперные Т-клетки «лицензируют» дендритные клетки, чтобы дать мощный активирующий сигнал наивному CD8+ Т-клетки.[5]
Кроме того, созревание CD8 + Т-клеток опосредуется CD40 сигнализация.[6] Как только наивная CD8 + Т-клетка связывается с инфицированной клеткой, инфицированная клетка запускается для высвобождения CD40.[6] Это высвобождение CD40 с помощью хелперных Т-клеток будет запускать дифференцировку наивных CD8 + Т-клеток в зрелые CD8 + Т-клетки.[6]
Хотя в большинстве случаев активация зависит от распознавания антигена TCR, описаны альтернативные пути активации. Например, было показано, что цитотоксические Т-клетки активируются под действием других Т-лимфоцитов CD8, что приводит к толеризации последних.[7]
После активации TC клетка подвергается клональной экспансии с помощью цитокина Интерлейкин-2 (ИЛ-2), который представляет собой рост и дифференциация фактор для Т-клеток. Это увеличивает количество клеток, специфичных для антигена-мишени, которые затем могут перемещаться по телу в поисках антиген-положительных соматические клетки.
Эффекторные функции
При контакте с инфицированными / дисфункциональными соматическими клетками TC клетки выделяют цитотоксины перфорин, гранзимы, и гранулизин. Благодаря действию перфорина гранзимы попадают в цитоплазму клетки-мишени и их сериновая протеаза функция запускает каспаза каскад, который представляет собой серию цистеиновых протеаз, которые в конечном итоге приводят к апоптоз (запрограммированная гибель клеток). Из-за высокого порядка липидов и отрицательно заряженного фосфатидилсерина, присутствующего в их плазматической мембране, TC клетки устойчивы к действию их цитотоксинов перфорина и гранзима.[8]
Второй способ вызвать апоптоз - это взаимодействие клеточной поверхности между ТC и инфицированная клетка. Когда TC активируется, он начинает экспрессировать поверхностный белок Лиганд FAS (FasL) (Apo1L) (CD95L), который может связываться с Фас Молекулы (Apo1) (CD95) экспрессируются на клетке-мишени. Однако считается, что это взаимодействие Fas-Fas-лиганда более важно для удаления нежелательных Т-лимфоциты во время их развития или литической активности определенных TЧАС клеток, чем цитолитическая активность ТC эффекторные клетки. Взаимодействие Fas с FasL позволяет рекрутировать сигнальный комплекс, индуцированный смертью (DISC).[9]Fas-связанный домен смерти (FADD) перемещается вместе с DISC, позволяя рекрутировать прокаспазы 8 и 10.[9] Эти каспазы затем активируют эффекторные каспазы 3, 6 и 7, что приводит к расщеплению субстратов смерти, таких как ламин А, ламин B1, ламин B2, PARP (поли-АДФ-рибоза-полимераза ), и ДНК-PKcs (ДНК-активированная протеинкиназа). Конечный результат - апоптоз клетки, экспрессирующей Fas.
Фактор транскрипции Эомесодермин предполагается, что он играет ключевую роль в функции CD8 + Т-клеток, действуя как регуляторный ген в адаптивном иммунном ответе.[10] Исследования, изучающие эффект потери функции эомезодермина, показали, что снижение экспрессии этого фактора транскрипции приводит к снижению количества перфорина, продуцируемого CD8 + Т-клетками.[10]
Роль в патогенезе болезни
В течение вирус гепатита В (HBV) цитотоксические Т-клетки убивают инфицированные клетки и продуцируют противовирусные цитокины, способные очищать HBV от жизнеспособных гепатоцитов. Они также играют важную патогенную роль, внося свой вклад почти во все повреждения печени, связанные с инфекцией HBV.[11] Тромбоциты было показано, что они способствуют накоплению вирус-специфических цитотоксических Т-клеток в инфицированной печени.[12]
Цитотоксические Т-клетки участвуют в прогрессировании артрит: истощение коленного сустава хрящ макромолекулы, такие как гликозаминогликаны цитотоксическими Т-клетками и макрофаги наблюдалась на крысиной модели болезни.[13]
Было обнаружено, что CD8 + Т-клетки играют роль в ВИЧ инфекционное заболевание. Со временем ВИЧ разработал множество стратегий, позволяющих уклоняться от иммунной системы клетки-хозяина. Например, ВИЧ имеет очень высокую частоту мутаций, позволяющую избежать распознавания CD8 + Т-клетками.[14] Они также способны подавлять экспрессию поверхностных белков MHC класса I инфицированных ими клеток, чтобы избежать разрушения CD8 + Т-клетками.[14] Если CD8 + T-клетки не могут найти, распознать и связаться с инфицированными клетками, вирус не будет уничтожен и продолжит расти.
Кроме того, недавно было обнаружено, что CD8 + Т-клетки играют решающую роль в Диабет 1 типа.[15] Ранее считалось, что это аутоиммунное заболевание контролируется исключительно клетками CD4 +, но недавние исследования на модели мышей с диабетом показали, что CD8 + Т-клетки также участвуют в разрушении продуцирующих инсулин клеток поджелудочной железы.[15]
Интересно, что для разрешения проблемы могут потребоваться CD8 + Т-клетки. периферическая невропатия, вызванная химиотерапией (CIPN).[16][17] У мышей без CD8 + Т-клеток наблюдается более продолжительный CIPN по сравнению с нормальными мышами, а инъекция образованных CD8 + T-клеток разрешает или предотвращает CIPN.
Смотрите также
Рекомендации
- ^ Аль-Шура АН (2020). «Лимфоциты». Продвинутая гематология в интегрированной сердечно-сосудистой китайской медицине. Эльзевир. С. 41–46. Дои:10.1016 / b978-0-12-817572-9.00007-0. ISBN 978-0-12-817572-9.
Хелперные Т-клетки / CD4 + • экспрессируют гликопротеины CD4 на своей клеточной поверхности, которые активируются в присутствии пептидных антигенов на поверхности вторгающихся патогенов; • немедленно реагировать, чтобы защитить иммунную систему; • секретируют различные цитокиновые белки в соответствии с иммунным ответом.
- ^ Вентури С., Вентури М. (сентябрь 2009 г.). «Йод, тимус и иммунитет». Питание. 25 (9): 977–9. Дои:10.1016 / j.nut.2009.06.002. PMID 19647627.
- ^ Hivroz C, Chemin K, Tourret M, Bohineust A (2012). «Перекрестные помехи между Т-лимфоцитами и дендритными клетками». Критические обзоры в иммунологии. 32 (2): 139–55. Дои:10.1615 / CritRevImmunol.v32.i2.30. PMID 23216612.
- ^ Хойер С., Проммерсбергер С., Пфайфер И.А., Шулер-Турнер Б., Шулер Г., Дёрри Дж., Шафт Н. (декабрь 2014 г.). «Одновременное взаимодействие DC с CD4 (+) и CD8 (+) T-клетками улучшает вторичную экспансию CTL: для танго нужны три». Европейский журнал иммунологии. 44 (12): 3543–59. Дои:10.1002 / eji.201444477. PMID 25211552.
- ^ Ланзавеккья А (1998). «Иммунология. Лицензия на убийство». Природа. 393 (6684): 413–414. Дои:10.1038/30845. PMID 9623994.
- ^ а б c Беннет С.Р., Карбон Ф.Р., Карамалис Ф., Флавелл Р.А., Миллер Дж.Ф., Хит В.Р. (июнь 1998 г.). «Помощь в ответах цитотоксических Т-клеток опосредуется передачей сигналов CD40». Природа. 393 (6684): 478–80. Bibcode:1998Натура.393..478Б. Дои:10.1038/30996. PMID 9624004.
- ^ Milstein O, Hagin D, Lask A, Reich-Zeliger S, Shezen E, Ophir E, Eidelstein Y, Afik R, Antebi YE, Dustin ML, Reisner Y (январь 2011 г.). «CTL реагируют активацией и секрецией гранул, когда служат мишенями для распознавания Т-клеток». Кровь. 117 (3): 1042–52. Дои:10.1182 / кровь-2010-05-283770. ЧВК 3035066. PMID 21045195.
- ^ Rudd-Schmidt JA, Hodel AW, Noori T., Lopez JA, Cho HJ, Verschoor S и др. (Ноябрь 2019 г.). «Порядок липидов и заряд защищают Т-киллеры от случайной смерти». Nature Communications. 10 (1): 5396. Дои:10.1038 / с41467-019-13385-х. ЧВК 6881447. PMID 31776337.
- ^ а б Бакши Р.К., Кокс М.А., Заяц А.Дж. (2014). «Цитотоксические Т-лимфоциты». Энциклопедия медицинской иммунологии. С. 332–342. Дои:10.1007/978-0-387-84828-0_36. ISBN 978-0-387-84827-3.
- ^ а б Пирс Е.Л., Маллен А.С., Мартинс Г.А., Кравчик С.М., Хатчинс А.С., Зедиак В.П., Баница М., ДиЧоччио С.Б., Гросс Д.А., Мао Калифорния, Шен Х., Цереб Н., Ян С.Ю., Линдстен Т., Россант Дж., Хантер К.А., Райнер С.Л. (Ноябрь 2003 г.). «Контроль функции эффекторных CD8 + Т-клеток с помощью фактора транскрипции эомезодермина». Наука. 302 (5647): 1041–3. Дои:10.1126 / science.1090148. PMID 14605368.
- ^ Iannacone M, Sitia G, Guidotti LG (2006). «Патогенетический и противовирусный иммунный ответ против вируса гепатита В». Будущая вирусология. 1 (2): 189–96. Дои:10.2217/17460794.1.2.189.
- ^ Iannacone M, Sitia G, Isogawa M, Marchese P, Castro MG, Lowenstein PR, Chisari FV, Ruggeri ZM, Guidotti LG (ноябрь 2005 г.). «Тромбоциты опосредуют поражение печени, вызванное цитотоксическими Т-лимфоцитами». Природа Медицина. 11 (11): 1167–9. Дои:10,1038 / нм 1317. ЧВК 2908083. PMID 16258538.
- ^ Субраманиан С., Рамалингам К. (2005). «Электронно-микроскопические доказательства участия цитотоксических Т-лимфоцитов и макрофагов в адъюванте Mtb, вызванном воспалением соединительной ткани и артритогенезом у Rattus norvegicus». Азиатский журнал микробиологии, биотехнологии и наук об окружающей среде. 7 (2): 227–233. ISSN 0972-3005.
- ^ а б Гульзар Н., Коупленд К.Ф. (январь 2004 г.). «CD8 + Т-клетки: функция и ответ на ВИЧ-инфекцию». Текущие исследования ВИЧ. 2 (1): 23–37. Дои:10.2174/1570162043485077. PMID 15053338.
- ^ а б Цай С., Шамели А., Сантамария П. (2008). «CD8 + Т-клетки при диабете 1 типа». Достижения в иммунологии. 100: 79–124. Дои:10.1016 / S0065-2776 (08) 00804-3. ISBN 9780123743268. PMID 19111164.
- ^ Лаумет Дж., Эдралин Дж. Д., Данцер Р., Хейнен С. Дж., Кавелаарс А. (июнь 2019 г.). «Цисплатин обучает CD8 + Т-клетки предотвращать и устранять вызванную химиотерапией периферическую нейропатию у мышей». Боль. 160 (6): 1459–1468. Дои:10.1097 / j.pain.0000000000001512. ЧВК 6527475. PMID 30720585.
- ^ Krukowski K, Eijkelkamp N, Laumet G, Hack CE, Li Y, Dougherty PM и др. (Октябрь 2016 г.). «CD8 + Т-клетки и эндогенный IL-10 необходимы для купирования нейропатической боли, вызванной химиотерапией». Журнал неврологии. 36 (43): 11074–11083. Дои:10.1523 / JNEUROSCI.3708-15.2016. PMID 27798187.
внешняя ссылка
 СМИ, связанные с Цитотоксические Т-клетки в Wikimedia Commons
СМИ, связанные с Цитотоксические Т-клетки в Wikimedia Commons- Группа Т-клеток - Кардиффский университет