Оленекян - Olenekian
| Система / Период | Серии / Эпоха | Этап / Возраст | Возраст (Ма ) | |
|---|---|---|---|---|
| Юрский | Ниже/ Рано | Геттангский | моложе | |
| Триасовый | Верхний / Поздно | Ретиан | 201.3 | ~208.5 |
| Norian | ~208.5 | ~227 | ||
| Карнийский | ~227 | ~237 | ||
| Середина | Ладинский | ~237 | ~242 | |
| Анисианский | ~242 | 247.2 | ||
| Ниже/ Рано | Оленекян | 247.2 | 251.2 | |
| Индуанский | 251.2 | 251.902 | ||
| Пермский период | Лопингиан | Changhsingian | старшая | |
| Подразделение триасовой системы согласно ICS, по состоянию на 2020 год.[1] | ||||
в геологическая шкала времени, то Оленекян является возраст в Ранний триас эпоха; в хроностратиграфия, это сцена в Нижний триас серии. Он охватывает время между 251,2 Ма и 247,2 млн лет назад.[2] Оленекян иногда делят на Смитиан и Спатианский подэтапы или подэтапы.[3] Оленекян следует за Индуанский и следует Анисианский (Средний триас ).[4]
В оленекяне отложение значительной части Buntsandstein в Европе. Оленекский ярус примерно ровесник региональному юннинчженьскому ярусу, использовавшемуся в Китай.

Стратиграфические определения
Оленекский ярус был введен в научную литературу русскими стратиграфами в 1956 г.[5] Сцена названа в честь Оленек в Сибирь. До того, как произошло разделение на оленекский и индский, оба этапа сформировали скифский этап, который с тех пор исчез из официальной шкалы времени.
Основание оленекского яруса находится в самом низком проявлении отложения. аммоноиды Геденстроемия или же Meekoceras gracilitatis, и конодонта Neospathodus waageni. Он определяется как оканчивающийся около самых низких вхождений роды Япониты, Параданубиты, и Paracrochordiceras; и из Conodont Chiosella timorensis. А GSSP (глобальный справочный профиль для базы) в 2009 г. еще не установлен.
Оленекянская жизнь


Жизнь все еще оправлялась от тяжелого массовое вымирание в конце перми. В оленекский период Флора изменено с ликопод преобладают (например, Плевромея ) к голосеменное растение и птеридофит преобладают.[6][7] Эти изменения растительности связаны с глобальными изменениями температуры и осадки. Хвойные (голосеменные ) были доминирующими растениями на протяжении большей части Мезозойский. Среди наземных позвоночных архозавры - группа диапсид рептилии, охватывающие крокодилы, птерозавры, динозавры, и в конечном итоге птицы - впервые возникла из архозаврический предки во времена оленекского. В эту группу входят свирепые хищники, такие как Erythrosuchus.
В океанах, Микробные рифы были обычным явлением в раннем триасе, возможно, из-за отсутствия конкуренции с многоклеточный строители рифов в результате исчезновения.[8] Тем не менее, временные рифы многоклеточных животных появлялись во время оленекского яруса везде, где это позволяли условия окружающей среды.[9] Аммоноиды и конодонты диверсифицированы, но оба понесли убытки во время Вымирание на границе Смита и Спата[10] в конце Смитианского подэтапа.
Рыбы со скатными плавниками в значительной степени осталась незатронутой событием пермско-триасового вымирания.[11][12] Многие роды демонстрируют космополитическое (всемирное) распространение во время Индуанский и Оленекян (например, Австралосомус, Birgeria, Parasemionotidae, Птеронискул, Ptycholepidae, Saurichthys ). Это хорошо видно на примере Грисбахян (рано Индуанский ) состарившиеся рыбные сообщества Формация Уорди Крик (Восток Гренландия ), Диенерианский (поздно Индуанский ) состаренные комплексы Формация Сакамена (Мадагаскар ), Формация Канделария (Невада, Соединенные Штаты ), и Микинская свита (Химачал-Прадеш, Индия ) и смитианские комплексы Формация Викингхогда (Шпицберген, Норвегия ), Формация Тэйнс (западный Соединенные Штаты ), и Формация Хелуншань (Аньхой, Китай ).
морской темноспондил амфибии, Такие как Афанерамма или же Wantzosaurus показать широкий географический диапазон во время Индуанский и оленекский возраст. Их окаменелости найдены в Гренландия, Шпицберген, Пакистан и Мадагаскар.[13] Первые морские рептилии появились в оленекский век.[13] Hupehsuchia, Ихтиоптеригия и Зауроптеригия являются одними из первых морских рептилий, вышедших на сцену (например, Cartorhynchus, Chaohusaurus, Utatsusaurus, Hupehsuchus, Гриппия, Омфалозавр, Корозавр ). Зауроптериги и ихтиозавры правили океанами во времена Мезозойский Эра.
В оленекском ярусе произошло крупное вымирание: Вымирание на границе Смита и Спата.[14] Вероятно, это событие было вызвано поздними извержениями вулкана. Сибирские ловушки и привели к вымиранию нескольких групп, особенно нектонический и пелагический таксоны Такие как аммоноиды и конодонты. Один из нескольких исключительно разнообразных комплексов раннего триаса, Парижская биота[15] (от близкого Париж, юго-восток Айдахо ), образовался после исчезновения на границе Смита и Спата. В нем не менее 7 тип и 20 различных многоклеточный заказы, в том числе лептомитид протомонаксонид губки (ранее известно только из Палеозой ), тилакоцефаланы, ракообразные, наутилоиды, аммоноиды, колеоиды, офиуроиды, морские лилии, и позвоночные.[16]

Хрящевые рыбы
| Chondrichthyes оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Лопингиан к Поздний мел | Свальбард | А гибодонт эластожаберный | ||
| Каменноугольный к Ранний триас | Канада | Хондрихтиан с неопределенным родством, известный прежде всего по перьевидным зубчикам. | ||
| Лопингиан к Поздний триас | Соединенные Штаты | А синеходонтиформный эластожаберный | ||
| Триасовый | Свальбард | А гибодонт эластожаберный | ||
| Ранний триас | Соединенные Штаты | |||


Рыбы со скатными плавниками
| Актиноптеригии оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Триасовый | США, Шпицберген | Неоптеригиан | ||
| Лопингиан к Средний триас | Свальбард | Неоптеригиан | ||
| Ранний триас | Гренландия, Мадагаскар | Неоптеригиан | ||
| Ранний триас | Свальбард | Платисиагид | ||
| Ранний триас к Средний триас | Свальбард | Неоптеригиан | ||
| Триасовый | США, Свальбард, Китай | Неоптеригиан | ||



Целаканты
| Актинистия оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас | Свальбард | Латимерия | ||
| Ранний триас | Свальбард | Латимерия | ||
| Ранний триас | Канада | Целакант с вилкохвостым хвостом | ||
| Ранний триас | Свальбард | Латимерия | ||
| Ранний триас | Свальбард | Латимерия | ||
| Ранний триас | Свальбард | Латимерия | ||

Двудомные рыбы
| Двоякодышащие оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас к эоцен | Польша, Россия, Антарктида, Германия | Ceratodontid dipnoan, близкий родственник современных австралийских двоякодышащих рыб. Neoceratodus. | ||
| Пермский период к Ранний триас | Польша, Россия | А гнаторхизид дипноан. | ||
| Ранний триас | Австралия | Маленький двуногий, вероятно, гнаторхизид. | ||
| Ранний триас | P. donensis: Россия ?П. sp.: Южная Африка | Род производных цератодонтоид двоякодышащая рыба. | |


† Темноспондилы
| Темноспондили оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас | Свальбард | А трематозаврид стереоспондил амфибия | ||
| Ранний спатский | Польша; Россия | А брахиопид стереоспондильные амфибии. | ||
| Индуанский в Смитиан | Мадагаскар | А капитозавр стереоспондил, тесно связанный с Watsonisuchus. | ||
| Ранний триас | Германия | А капитозавр стереоспондиловые амфибии | ||
| Спатианский | Германия | А капитозавр стереоспондиловые амфибии | ||
| Ранний триас | Европа; Центральная Азия | Широко распространенный и разнообразный род капитозавр стереоспондильные амфибии. | ||
| Поздний спатианский - Ранний анисианский | Формация Терригал, Австралия | А брахиопид стереоспондильные амфибии. | ||
| Средний триас | Африка; Северная Америка | А капитозавр стереоспондиловые амфибии | ||
| Германия | А капитозавр стереоспондиловые амфибии | |||
| Ранний триас | Германия; Россия | А трематозаврид стереоспондиловые амфибии | ||
| Поздний ветлугиан | Россия | А капитозавр стереоспондиловые амфибии | ||
| W. angustifrons: Слудкян Горизонт В. Малахови: Устмилианский Горизонт | Россия и Гренландия | А Capitosaurid стереоспондиловые амфибии, используемые как индекс окаменелости. | |






† Хрониозухианы
| Хрониозухия оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Россия | ||||
| Лопингиан к Ранний триас | Россия, Китай | |||
| Ранний триас | Россия | ||


Лиссамфибия
| Лиссамфибии индуса | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Польша | Чаткобатрахус - вымерший род лягушачьих амфибий. Это с Триадобатрахус, один из двух старейших известных лиссамфибий. Точнее, это член Salientia; это связано, но вне Анура, который включает всех существующих лягушек. | |||
† Проколофонономорфы
| Проколофонономорфа оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас | Германия, Швейцария | Род ящерицы парарептильный | ||

Архозавроморфы
| Архозавроморфы оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Волгоградская область, Россия | Самый старый из известных танистрофеид. | |||
| Остров Колгуев, арктическая Россия | Базальная архозавроморфы рептилия спорной классификации, различные исследования считали его близким родственником Пролацерта, танистрофеиды, оба или ни один. | |||
| Липовская свита, Россия | А гребневидный сучок. | |||
| Южная Африка, Китай | А протерозухид это одна из самых ранних известных архозавриформ. Его длина превышала 2 метра (6 футов), и считается, что он вел себя как современный крокодил. У его рта были две отличительные особенности: верхняя часть его челюсти загнута вниз, чтобы помочь удерживать добычу, и верхнее небо было выстлано рядом зубов (примитивная особенность, утраченная у более поздних архозавров). | |||
| Формация Соллинг, Германия | А гребневидный сучок. Ктенозаврикус один из старейших архозавры известно на сегодняшний день. | |||
| Ранний триас | Польша | |||
| 250-237,2 млн лет, Оленекян до Ладинский | Формация Омингонд, Намибия Южная Африка | Самый большой эритросухид. Одной из немногих отличительных черт, помимо размера, является гладкость края чешуйчатой кости. У других эритрозухид край этой кости выступает назад от черепа, придавая ему вид крючка. В Erythrosuchus, край выпуклый, без крючка. | ||
| 247,3-245 млн лет, Оленекян к Анисианский | Россия | Потенциально сомнительный архозавроморф из неопределенное родство. Первоначально считалось протерозухид архозаврический, анализ 2016 года показал, что он также имеет общие функции с танистрофеиды. | ||
| Россия | Эритросухида длиной примерно 1,50–2 метра (5–6 футов 8 дюймов). | |||
| ~ 252–247 млн лет, Индуанский Оленекяну | Катбергская формация, Ист-Кейп, Южная Африка | Первоначально считавшиеся предками современных ящериц, более позднее исследование вместо этого показало, что они имеют больше общего с родословной, которая приведет к архозаврам. | ||
| 252-250 млн лет, от Инда до Оленекяна | Южная Африка Китай | Протерозухид, который был одним из крупнейших наземных рептилий в раннем триасе. Он был чем-то похож на примитивный крокодил, и имел многие из их современных черт, таких как длинные челюсти, мощные мышцы шеи, короткие ноги и длинный хвост, обладая при этом некоторыми особенностями, уникальными для протерозухид, такими как крючковидный рот и длинные ряды простых конусовидных зубов. Эта челюсть могла быть приспособлением для ловли добычи, например Lystrosaurus. | ||
| Россия, | Возможно Rauisuchid. Скифозух был от 2 до 3 метров в длину и относительно крепко сложен. | |||
| Индуан в Оленекян | Ноклофти Формирование, West Hobart, Тасмания | Когда-то этот таксон считался протерозухидом, а теперь считается, что он был промежуточным звеном между продвинутыми неархозавровидными архозавроморфами, такими как Пролацерта, и базальные архозавриды, такие как Протерозух. Этот род также известен как одна из наиболее полных известных австралийских рептилий триаса. | ||
| Индуан в Оленекян | Южный Урал, Россия | Первоначально классифицировался как Rauisuchid, Цилмозух совсем недавно был интерпретирован как неопределенный архозавриформ. Цилмосухус происходили на протяжении всего оленекского века. Некоторые из слоев, из которых был найден Tsylmosuchus, имеют индийский возраст, что делает его одним из самых ранних архозавров. | ||
| последний Оленекян | Яренскян Горизонт, Республика Коми, Россия | Род паракрокодиломорфов архозавров, возможно, Rauisuchid, и старейший из известных архозавров из России. | ||
| Формация Хешангоу, Китай | А гребневидный сучок. Xilousuchus один из старейших архозавры известно на сегодняшний день. | |||






Лепидозавроморфы
| Лепидозавроморфа оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас | Польша | |||
† Зауроптеригия
| Зауроптериги оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Поздний оленекян | США, Вайоминг | А пистозавр. | ||
| Оленекян | Китай | Возможный пахиплеврозавр. | ||
| Оленекян | Китай | Возможный пахиплеврозавр. | ||

† Ихтиозавроморфы
† Hupehsuchians
| †Hupehsuchia оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас | Китай | An ихтиозавроморф |  Безымянный полидактильный hupehsuchian | |
| Ранний триас | Китай | An ихтиозавроморф | ||
| Ранний триас | Китай | An ихтиозавроморф | ||
| Ранний триас | Китай | An ихтиозавроморф | ||
| Ранний триас | Китай | An ихтиозавроморф | ||

† Ихтиозавриформы
| †Ихтиозаврообразные оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас | Китай | Рано ихтиозавриформ | ||
| Ранний триас | Китай | Рано ихтиозавриформ | ||


† Ихтиоптериги
| †Ихтиоптеригия оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Ранний триас | Китай | |||
| Ранний триас | Канада, Япония, Гренландия, Китай | |||
| Ранний триас к Средний триас | Свальбард | Рано ихтиозавриформ с зубцами пуговицами | ||
| Ранний триас | Свальбард | |||
| Ранний триас | Префектура Мияги, Япония; британская Колумбия, Канада | Рано ихтиозавриформ | ||




Терапсиды
| Терапсиды оленекского | ||||
|---|---|---|---|---|
| Таксоны | Присутствие | Место расположения | Описание | Изображений |
| Каннемейерия | Формация Бургерсдорп, Южная Африка Формация Нтавере, Замбия Формация Омингонд, Намибия Формация Манда, Танзания | А Kannemeyeriid это был один из первых представителей семейства, а значит, и один из первых крупных травоядных животных триаса. Хотя у него была большая голова, он был легким из-за размера глазниц и носовой полости. У него также были пояса конечностей, которые образовывали массивные костяные пластинки, которые помогали поддерживать его крепкое тело. |  | |
| Lystrosaurus | Формация Фремоу, Антарктида | 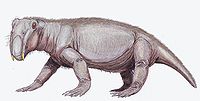 | ||
| Trirachodon | Формация Дрикоппен, Свободный штат, Южная Африка Формация Омингонд, Намибия |  | ||
Рекомендации
Примечания
- ^ «Международная хроностратиграфическая карта» (PDF). Международная комиссия по стратиграфии. 2020.
- ^ По Градштейну (2004). Brack и другие. (2005) дают от 251 до 248 млн лет
- ^ Тозер Э. Т. (1965): Этапы нижнего триаса и зоны аммоноидей Арктической Канады: Документ Геологической службы Канады 65: 1–14.
- ^ См. Подробную геологическую шкалу времени Gradstein и другие. (2004)
- ^ Кипарисова и Попов (1956)
- ^ Шнебели-Германн, Эльке; Kürschner, Wolfram M .; Керп, Ганс; Бомфлер, Бенджамин; Хочули, Петр А .; Бухер, Хьюго; Уэр, Дэвид; Рухи, Газала (апрель 2015 г.). «История растительности на границе перми и триаса в Пакистане (разрез Амб, Соляной хребет)». Исследования Гондваны. 27 (3): 911–924. Bibcode:2015GondR..27..911S. Дои:10.1016 / j.gr.2013.11.007.
- ^ Гудеманд, Николас; Романо, Карло; Лей, Марк; Бухер, Хьюго; Троттер, Джули А .; Уильямс, Ян С. (август 2019 г.). «Динамическое взаимодействие между климатом и потрясениями морского биоразнообразия во время раннего триасового Смит-Спатского биотического кризиса». Обзоры наук о Земле. 195: 169–178. Bibcode:2019ESRv..195..169G. Дои:10.1016 / j.earscirev.2019.01.013.
- ^ Фостер, Уильям Дж .; Хендель, Катрин; Ричоз, Сильвен; Глива, Яна; Lehrmann, Daniel J .; Бауд, Аймон; Колар ‐ Юрковшек, Чай; Алинович, Дунья; Юрковшек, Богдан; Корн, Дитер; Martindale, Rowan C .; Пекманн, Йорн (20 ноября 2019 г.). «Подавление конкурентного исключения позволило размножить микробиалиты на границе между перми и триасом». Отложение записи. 6 (1): 62–74. Дои:10.1002 / dep2.97. ЧВК 7043383. PMID 32140241.
- ^ Брайярд, Арно; Веннин, Эммануэль; Оливье, Николя; Bylund, Кевин Дж .; Дженкс, Джим; Стивен, Дэниел А .; Бухер, Хьюго; Хофманн, Ричард; Гудеманд, Николас; Эскаргуэль, Жиль (18 сентября 2011 г.). «Временные рифы многоклеточных животных после массового вымирания в конце перми». Природа Геонауки. 4 (10): 693–697. Bibcode:2011НатГе ... 4..693B. Дои:10.1038 / ngeo1264.
- ^ Галфетти, Томас; Хочули, Петр А .; Брайярд, Арно; Бухер, Хьюго; Вайссерт, Гельмут; Вигран, Йорун Ос (2007). «Смитско-спатийское пограничное событие: свидетельство глобального изменения климата после биотического кризиса в конце пермского периода». Геология. 35 (4): 291. Bibcode:2007Гео .... 35..291Г. Дои:10.1130 / G23117A.1.
- ^ Романо, Карло; Koot, Martha B .; Коган, Илья; Брайярд, Арно; Миних, Алла В .; Бринкманн, Винанд; Бухер, Хьюго; Кривет, Юрген (февраль 2016 г.). «Пермско-триасовые Osteichthyes (костистые рыбы): динамика разнообразия и эволюция размеров тела». Биологические обзоры. 91 (1): 106–147. Дои:10.1111 / brv.12161. PMID 25431138. S2CID 5332637.
- ^ Smithwick, Fiann M .; Стаббс, Томас Л. (2 февраля 2018 г.). «Выжившие в фанерозое: эволюция актиноптерисов в периоды массового вымирания в пермо-триасовом и триас-юрском периодах». Эволюция. 72 (2): 348–362. Дои:10.1111 / evo.13421. ЧВК 5817399. PMID 29315531.
- ^ а б Scheyer, Torsten M .; Романо, Карло; Дженкс, Джим; Бухер, Хьюго; Фарке, Эндрю А. (19 марта 2014 г.). «Восстановление морской биотики в раннем триасе: взгляд на хищников». PLOS ONE. 9 (3): e88987. Bibcode:2014PLoSO ... 988987S. Дои:10.1371 / journal.pone.0088987. ЧВК 3960099. PMID 24647136.
- ^ Галфетти, Томас; Хочули, Петр А .; Брайярд, Арно; Бухер, Хьюго; Вайссерт, Гельмут; Вигран, Йорун Ос (2007). «Смитско-спатийское пограничное событие: свидетельство глобального изменения климата после биотического кризиса в конце пермского периода». Геология. 35 (4): 291. Bibcode:2007Гео .... 35..291Г. Дои:10.1130 / G23117A.1.
- ^ Брайярд, Арно; Krumenacker, L.J .; Боттинг, Джозеф П .; Дженкс, Джеймс Ф .; Bylund, Кевин Дж .; Фара, Эммануэль; Веннин, Эммануэль; Оливье, Николя; Гудеманд, Николас; Сосед, Томас; Шарбонье, Сильвен; Романо, Карло; Догужаева, Лариса; Туи, Бен; Хаутманн, Михаэль; Стивен, Дэниел А .; Томазо, Кристоф; Эскаргель, Жиль (15 февраля 2017 г.). «Неожиданная морская экосистема раннего триаса и подъем современной эволюционной фауны». Достижения науки. 3 (2): e1602159. Bibcode:2017SciA .... 3E2159B. Дои:10.1126 / sciadv.1602159. ЧВК 5310825. PMID 28246643.
- ^ Специальный выпуск о Paris Biota: https://www.sciencedirect.com/journal/geobios/vol/54
Литература
- Брэк, Питер; Рибер, Ганс; Никора, Альда; Мундил, Роланд (1 декабря 2005 г.). «Глобальный пограничный стратотипический разрез и точка (GSSP) ладинского яруса (средний триас) в Баголино (Южные Альпы, Северная Италия) и его значение для временной шкалы триаса». Эпизоды. 28 (4): 233–244. Дои:10.18814 / epiiugs / 2005 / v28i4 / 001.
- Gradstein, F.M .; Огг, Дж. И Смит, А.; 2004: Шкала геологического времени 2004, Издательство Кембриджского университета.
- Кипарисова, Л. И Попов, Ю.; 1956: Расчленение нижнего отдела триасовой системы на ярусы (Подразделение нижнего ряда триасовой системы на этапы), Доклады Академии Наук СССР 109(4), стр. 842–845. (на русском).
внешняя ссылка
- База данных GeoWhen - Оленекян
- Шкала времени нижнего триаса на сайте подкомиссии по стратиграфической информации ИКС
- Шкала времени нижнего триаса на веб-сайте Норвежской сети морских записей геологии и стратиграфии.
Координаты: 31 ° 57′55 ″ с.ш. 78 ° 01′29 ″ в.д. / 31,9653 ° с. Ш. 78,0247 ° в.