Формация Тяоцзишань - Tiaojishan Formation
| Формация Тяоцзишань Стратиграфический диапазон: Батонский -Оксфордский ~165–153 Ма | |
|---|---|
| Тип | Геологическое образование |
| Подразделения | Кровать Daohugou |
| Лежит в основе | Формация Тучэнцзы, Формация Хоучэн |
| Перекрывает | Формация Хайфангоу |
| Толщина | 2,420 м (7,940 футов) |
| Литология | |
| Начальный | Андезит |
| Другой | Песчаник, сланец, туф, каменный уголь |
| Место расположения | |
| Координаты | 41 ° 18′N 119 ° 12'E / 41,3 ° с. Ш. 119,2 ° в.Координаты: 41 ° 18′N 119 ° 12'E / 41,3 ° с. Ш. 119,2 ° в. |
| Приблизительные палеокоординаты | 43 ° 00′N 123 ° 06′E / 43,0 ° с. Ш. 123,1 ° в. |
| Область, край | Хэбэй, Внутренняя Монголия, & Ляонин |
| Страна | |
| Степень | Пояс Яньшань |
 Формация Тяоцзишань (Китай) | |
В Формация Тяоцзишань это геологическая формация в Хэбэй и Ляонин, Китайская Народная Республика, относящиеся к средне-позднеюрский период (Батонский -Оксфордский этапы). Он известен своими исключительно сохранившийся окаменелости, в том числе растения, насекомые и позвоночные. Он состоит в основном из пирокластическая порода с вкраплениями основных вулканических и осадочные породы. Ранее формация Тяоцзишань группировалась вместе с нижележащими слоями. Формация Хайфангоу (также известная как формация Цзюлуншань) как единое целое "Формация Ланьци."[1] Большинство исследователей согласны с тем, что Кровать Daohugou, ранее спорная датировка, является частью формации Тяоцзишань.[2] Формация Тяоцзишань является ключевой частью Янляо Биота сборка.
Геология
Геология пласта Даохугоу сбивает с толку, потому что она сложна и не соответствует; Это означает, что элементы и слои горных пород разного возраста смешаны вместе в результате складчатости, эрозии и вулканической активности. Лю и др. (2006) пришли к выводу, что породы, несущие Даохугоу Биота также включают свиты Тяоцзишань и Ланьци. Они продемонстрировали, что Формация Цзюлуншань старше (Средняя юра ), и что Формация Тучэнцзы моложе (Позднеюрский период ). Однако многие другие исследователи считают Даохугоу частью самой формации Цзюлуншань.[3]
Полевые исследования, опубликованные в 2006 году, также показали, что грядки одинаковы на большой площади; с запада Ляонин в Нинчэн графство Внутренняя Монголия (Ней Монгол).[4]
Возраст
С помощью Аргон-аргоновое датирование, Ван и его коллеги в 2005 году датировали часть формации Тяоцзишань примерно 160 миллионами лет назад, началом Оксфордский этап, первый этап Верхняя юра эпоха.[5] В 2006 году в исследовании Лю и его коллег U-Pb датировка циркона сделать вывод, что формация Тяоджишань коррелирует со слоями Даохугоу, а полный хронологический диапазон этой общей биоты датируется 168–164/152 млн лет назад.[6] Последующее исследование, опубликованное в 2008 году, дополнительно уточнило возрастной диапазон формации, обнаружив, что нижняя граница Тяоджишань была сформирована 165 млн лет назад, а верхняя граница - где-то между 156-153 млн лет назад.[7]
Кровать Daohugou
Возраст кровати Даохугоу обсуждался, и ряд исследований с использованием различных методологий пришли к противоречивым выводам. Различные документы помещают окаменелости сюда как находящиеся где-то вдали от Средняя юра период (169 миллионов лет назад) до Раннемеловой период период (122 мА).[8] Одно из первых исследований возраста пластов Даохугоу, опубликованное в 2004 г. и другие., обнаружил, что они относятся к раннему меловому периоду, всего на несколько миллионов лет старше вышележащих пластов Джехол Формация Исянь.[9] В исследовании 2004 г. в основном использовались Аргон-аргоновое датирование из туф в слоях Даохугоу, чтобы определить его возраст.
Однако последующие исследования ставят под сомнение этот относительно недавний возраст. В исследовании 2006 года Гао и Рен критиковали Хэ и другие. за то, что они не включили в свою статью достаточно конкретики и деталей, а также не согласились с их радиометрическим датированием туфа Даохугоу. Туф, как утверждали Гао и Рен, содержит кристаллы с множеством различных радиометрических возрастов, некоторым из которых до миллиарда лет, поэтому использование дат только нескольких из этих кристаллов не может определить общий возраст отложений. Гао и Рен продолжали защищать средний юрский период для пластов на основе биостратиграфия (использование индекс окаменелости ), а также отношения пласта к слою, который, как известно, отмечает границу средней и поздней юры.[10]
Другое исследование, опубликованное Вангом в 2006 г. и другие., утверждал, что формация Тяоцзишань, возраст которой 159–164 миллиона лет, лежит в основе, а не перекрывает пласты Даохугоу. В отличие от более раннего исследования Гао и Рен, Ван и другие. обнаружили общее сходство между ископаемыми животными, найденными в слоях Даохугоу, и животными из Формация Исянь. Авторы заявили, что
"окаменелости позвоночных, такие как Ляокситритон, Jeholopterus и пернатые манирапторанс показывают большое сходство с таковыми из формации Исянь. Другими словами, несмотря на отсутствие Lycopteraгруппа позвоночных даохуго, типичная рыба биоты Джехол, ближе к таковой из раннемеловой биоты Джехол, чем к любой другой биоте ».
Ван и другие. пришел к выводу, что Даохуго, вероятно, представляет собой самые ранние эволюционные стадии Джехол Биота и что она «принадлежит к тому же циклу вулканизма и седиментации, что и формация Исянь группы Джехол».[4] Однако более позднее исследование Джи и другие. утверждал, что ключевым индикатором биоты Джехола являются индексные окаменелости Пейпяостей и Lycoptera. Согласно этому определению, самый ранний этап эволюции биоты Джехол представлен Формация Хуацзин, а формация Даохугоу исключена из-за отсутствия Lycoptera окаменелости.[11] Позже в 2006 году Лю и другие. опубликовали собственное исследование возраста пластов Даохугоу, на этот раз с использованием Циркона. Уран-свинцовые датировки на вулканических породах, перекрывающих и подстилающих слои, содержащие саламандр (саламандры часто используются как индексные окаменелости). Лю и другие. обнаружили, что пласты сформировались между 164-158 миллионами лет назад, в средней и поздней юре.[6][12] Исследование 2012 года, проведенное Гао и Шубином, согласилось с этой оценкой и сообщило, что аргон-аргонная дата для горизонта Даохуго составила 164 плюс-минус 4 миллиона лет назад.[13]
Климат
Основываясь на растительности, присутствующей в формации Тяоджишань, Ван Юндун и его коллеги определили, что климат в Ляонине в середине юрского периода был субтропический к умеренный, тепло и влажный.[1]
Фауна
Красиво сохранившиеся окаменелости динозавры, птерозавры, саламандры, насекомые, паукообразные[14] и другие беспозвоночные, хвойные породы, гинкго, саговники, хвощи, и папоротники, и даже самое раннее из известных планирующих млекопитающих (Волатикотерий ) и водное животное (Castorocauda ) были обнаружены в этих породах. Эти организмы были частью Даохугоу Биота, которые сформировали локальную экосистему того юрского времени. Туфогенный состав некоторых слоев горных пород показывает, что это была вулканическая область, которая иногда подвергалась сильным пеплопадам в результате извержений. В пейзаже тогда преобладали горные ручьи и глубокие озера, окруженные лесами. голосеменное растение деревья.[15] Некоторые авторы[ВОЗ? ] пришли к выводу, что биота Даохуго является ранней стадией Джехол Биота, а недавняя работа[нужна цитата ] продемонстрировал, что эти два отличия.
Леса биоты Даохугоу росли во влажном умеренно-теплом климате, и в них преобладали голосеменное растение деревья. Существовал гинкгопсид подобно Гинкоиты, Гинкго, Байера, Чекановская, и Феникопсис. Были также хвойные породы подобно Питиофиллум, Рипидиоклад, Элатоклад, Шизолепис, и Подозамитесь. Также, Ликопсиды подобно Lycopodites и Продажа, хвощ (Sphenopsida ) подобно Equisetum, саговники подобно Аномозамиты, и папоротники (Filicopsida ) подобно Todites и Coniopteris.[16]
Саламандры
| Саламандры формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
Б. jianpingensis[13] | Ляонин | А саламандроид известен по почти полному и сочлененному скелету, открытому с вентральной стороны. | ||||
С. tianyiensis | А криптобранхоидный длиной 18 сантиметров. | |||||
Дж. Парадокс | Криптобранхоид со странной морфологией черепа, сначала предполагалось, что он произошел от Раннемеловой период. | |||||
L. daohugouensis | Малоизвестный криптобранхоид. | |||||
P. sinensis | Криптобранхоид, характеризующийся коротким туловищем (всего 14 пресакральных звеньев) и короткой и широкой головой, придает форму толстому телу, от которого и произошло название рода («Пан» на китайском означает жир). | |||||

Птерозавры
| Птерозавры формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
А. linglongtaensis[17] | Ляонин | А монофенестратан известен по неполному скелету с частичным черепом и нижней челюстью. | ||||
К. пани[18] | А птеродактилиформ известна только по единственному экземпляру молодой особи с размахом крыльев 475 миллиметров (18,7 дюйма). | |||||
D. delicatus[19] | Один экземпляр | Относительно базальный птерозавр, известный по частичному скелету с отпечатками мягких тканей. | ||||
D. modularis[20] | Ляонин | А Wukongopterid названный в честь Чарльз Дарвин. Типовой вид, D. modularis был первым известным птерозавром, у которого были черты длиннохвостого рамфоринхоиды и короткохвостый птеродактилоиды, и был описан как переходное ископаемое между двумя группами. Дарвиноптер Сообщалось также, что экземпляры демонстрируют несколько различий между самцами и самками, причем самцы имеют характерные гребни на головах. Известно, что они откладывали яйца на землю и, возможно, также не проявляли особой заботы о своих родителях. | ||||
D. linglongtaensis | ||||||
D. robustodens | ||||||
D. mutoudengensis[21] | Хэбэй | Один экземпляр | А батрахогнатин анурогнатид Первоначально считалось, что это животное из раннего мелового периода, с размахом крыльев около 40 сантиметров, что делает его одним из самых маленьких известных птерозавров. | |||
F. lii[22] | Ляонин | А скафогнатин рамфоринхид похож на других скафогнатинов своим коротким тупым черепом с большим анторбитальным отверстием и широко расставленными вертикально ориентированными зубами (в отличие от горизонтально ориентированных зубов других рамфоринхид). | ||||
Дж. Ninchengensis | Внутренняя Монголия | Батрахогнатин анурогнатид, сохранившийся с пикнофибрами и остатками кожи. | ||||
J. robustus[25] | Ляонин | Скафогнатин рамфоринхид, известный по единственному ископаемому скелету. | ||||
J. zhaoianus[26] | Ляонин | Скафогнатий рамфоринхид известен по почти полному скелету с сохранившимся черепом. | ||||
К. sinensis | Ляонин | Кровать Daohugou | Один почти полный экземпляр | Вуконгоптерид с удлиненной головой, длиной 106,9 миллиметра. | ||
П. wellnhoferi | Внутренняя Монголия | Кровать Daohugou | Один экземпляр[23] | А дарвиноптер с высоким гребнем на голове и удлиненным черепом длиной 11,8 сантиметра (4,65 дюйма), длинным хвостом и размахом крыльев около 85 сантиметров (33,46 дюйма). Первоначально считалось, что это рамфоринхид. Единственный известный экземпляр состоит из сочлененного, почти полного скелета с остатками покровов. Они включали крыловую мембрану, волосовидные пикнофибры, длинную версию лопасти, находящуюся на конце «рамфоринхоидных» хвостов, и гребень головы с низким костным основанием и большим кератиновым удлинением. | ||
Q. guoi[27] | Ляонин | А рамфориншин рамфоринхид известен только по одному экземпляру, включающему скелет с черепом. | ||||
W. lii | Ляонин | Кровать Daohugou | Один экземпляр[28] | Вуконгоптерид необычен тем, что имеет удлиненную шею и длинный хвост. Его размах крыльев оценивается в 730 миллиметров (29 дюймов). | ||



Динозавры
| Динозавры формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
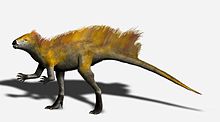
А. Хаксли[29] | Несколько экземпляров[30] | An анхиорнитид сначала считалось Troodont. Учитывая изысканную сохранность окаменелостей одного из первых экземпляров, Анхиорнис стал первым видом динозавров, для которого можно было определить почти всю окраску жизни. Большинство перьев тела Анхиорнис были серые и черные. Макушка головных перьев была преимущественно рыжей с серым основанием и передней частью, а на лице были рыжеватые крапинки среди преимущественно черных перьев головы. Перья на крыльях и задних конечностях белые с черными кончиками. Покровы были серыми, контрастируя с белыми основными крыльями. Более крупные кроющие крыла также были белыми с серыми или черными кончиками, образующими ряды более темных точек вдоль середины крыла. Они имели форму темных полос или даже рядов точек на внешнем крыле (первичные кроющие перья), но более неравномерный ряд пятен на внутреннем крыле (вторичные кроющие). Голени были серыми, за исключением длинных перьев ног, а ступни и пальцы ног были черными. Он был 13 дюймов в длину и весил всего 110 граммов (3,9 унции). | ||||
А. Сюй | Ляонин | Один экземпляр | Анхиорнитид размером примерно с современного фазан, длиной 20 дюймов. Кости его ноги были похожи на кости Археоптерикс, но в целом его анатомия была более примитивной. | |||
К. джуджи | Янляо Биота | Один экземпляр | Анхиорнитид, известный по взрослому экземпляру, длина тела которого составляет 400 мм. Его окаменелые перья обладают наноструктурами, которые были проанализированы и интерпретированы как меланосомы, показывающие сходство с органеллами, которые дают черный переливающийся цвет у некоторых видов современных животных. птицы. Другие перья, обнаруженные на голове, груди и основании хвоста, сохраняют сплющенные пластинки пластинчатых меланосом, очень похожие по форме на те, которые создают яркие переливающиеся оттенки в перьях современного человека. колибри. Однако эти структуры кажутся твердыми и в них отсутствуют пузырьки воздуха, и поэтому внутренне они больше похожи на меланосомы в трубачи чем колибри. Caihong представляет собой старейшее известное свидетельство наличия тромбоцитов меланосом. Он назван в честь большого гребня на слезной кости черепа.[31] | |||
E. brevipenna | Ляонин | Один экземпляр | Поначалу считалось, что анхиорнитид - троодонт, известный по единственному ископаемому образцу, представляющему почти полный скелет малолетнего или взрослого человека. Образец очень маленький, около 12 дюймов в длину. | |||
E. hui | Кровати Daohugou | Один экземпляр | А скансориоптеригид известен по хорошо сохранившемуся частичному скелету длиной 10 дюймов (17,5 дюймов, включая неполные рулевые перья), который включает четыре длинных пера на хвосте, состоящих из центрального стержня и лопастей. Однако, в отличие от прямоугольников в современном стиле, лопатки не разветвлялись на отдельные нити, а состояли из одного лентообразного листа. Эпидексиптерикс также сохранился покров из более простых перьев тела, состоящих из параллельных зубцов, как у более примитивных пернатых динозавров. Однако перья тела Эпидексиптерикс уникальны тем, что некоторые из них возникают из «перепончатой структуры» у основания каждого пера. Было высказано предположение, что это может представлять собой этап в эволюции пера. Эпидексиптерикс и его родственники представляют собой самые ранние известные образцы декоративных перьев в летописи окаменелостей. | |||
П. daohugouensis | Внутренняя Монголия | Кровати Daohugou | Один экземпляр | Анхиорнитид, который, вероятно, имел длину 1 метр (3 фута) или меньше, но поскольку этот вид известен только по задним ногам, реальную длину трудно оценить. Помимо очень птичьего скелета на ногах, Педопенна был примечателен наличием длинных перистых перьев на плюсне (стопе). Некоторые другие паравианцы также известны эти «задние крылья», но у Педопенна отличаются от животных, подобных Микрораптор. Педопенна задние крылья были меньше и более округлой формы. Самые длинные перья были немного короче плюсны, около 55 мм (2 дюйма) в длину. Кроме того, перья Педопенна были симметричными, в отличие от асимметричных перьев некоторых других нептичьих паравиан и птиц. Поскольку асимметричные перья типичны для животных, приспособленных к полету, вполне вероятно, что Педопенна представляет собой раннюю стадию развития этих структур. Хотя многие отпечатки перьев в окаменелостях слабы, ясно, что у каждого из них были рахис и зазубрины, и хотя точное количество перьев на лапах не известно, их больше, чем на задних крыльях Микрораптор. Педопенна также показывает свидетельство более коротких перьев, покрывающих перья на длинных лапах, свидетельство присутствия кроющих, наблюдаемых у современных птиц. Поскольку у перьев меньше аэродинамических приспособлений, чем у аналогичных задних крыльев Микрораптор, и кажутся менее жесткими, предполагает, что если у них и была какая-то аэродинамическая функция, то она была намного слабее, чем у других нептичьих паравиан и птиц. | ||
С. хейлманни | Ляонин | Точное происхождение типового образца неизвестно, скорее всего, из слоев Даохугоу.[4] | Один или два экземпляра | А воробей -размерный скансориоптеригид, известный по молодому экземпляру. | ||
S. sungei | Ляонин | Один экземпляр[32] | Анхиорнитид с плюмовидными перьями. Отпечатки перьев включают тонкие пучки на шее, короткие и симметричные лопаточные перья на руках, а также пушистые и длинные перистые перья на задних конечностях (поразительно напоминающие тонкие волокна задних конечностей с современными. Silkie порода домашняя курица. В то время как его анатомия и покровы имеют общие черты с птицами, а также производные дромеозавры Такие как МикрорапторКладистический анализ помещает этот род в группу пернатых нептичьих динозавров, близкую к происхождению птичьих. Вряд ли это был летчик. | |||
T. confuciusi | Ляонин | А гетеродонтозавр первоначально сообщалось, что он принадлежит к группе Джехол раннемелового периода. Окаменелость была собрана в местности, транслитерированной как Лингленгта или Линглонгта. Lu et al., 2010 сообщили, что эти пласты на самом деле были частью формации Тяоцзишань, датируемой поздним юрским периодом. Тяньюлонг имеет ряд длинных нитчатых покровных структур на спине, хвосте и шее особи, подобных перьям, обнаруживаемым у некоторых теропод (это предполагает, что все гетеродонтозавры могли иметь эти волокна). Голотип принадлежит малолетнему взрослому человеку, длина которого, вероятно, составляла 70 сантиметров, исходя из пропорций родственного ему человека. Южноафриканский разновидность Гетеродонтозавр. | ||||
X. zhengi[33] | Ляонин | Один экземпляр | Первоначально считалось, что анхиорнитид - это либо дромеозавр, либо троодонт. | |||
Ю. ци[34] | Кровати Daohugou | Один экземпляр | Планирующий скансориоптеригид весом около 380 граммов (0,84 фунта), который, как и другие скансориоптеригиды, обладал необычным удлиненным третьим пальцем, который (в случае Йи) помогал поддерживать пленочную скользящую плоскость из кожи. Самолеты Йи также поддерживались длинной костлявой стойкой, прикрепленной к запястью. Эта модифицированная кость запястья и самолет на основе мембраны уникальны среди всех известных динозавров и могли привести к появлению крыльев, похожих на крылья летучие мыши. Это также приводит к гипотезе о том, что два других рода скансориоптеригид также имели скользящие мембраны, но это еще не подтверждено официально. | |||









Ящерицы
Цветовой ключ
| Примечания Неопределенные или предположительные таксоны входят в маленький текст; |
| Лепидозавры (ящерицы и родственники) слоев Даохугоу | |||||
|---|---|---|---|---|---|
| Род | Разновидность | Состояние | Избыток | Примечания | Изображений |
Безымянная ящерица[35] | Внутренняя Монголия | Один экземпляр | Новая ящерица с относительно короткими передними конечностями | ||
Безымянная ящерица[35] | Внутренняя Монголия | Один экземпляр | Ящерица с длинными задними конечностями и узким телом | ||
Цинодонты
| Цинодонты свиты Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
A. scansorius | Кровать Daohugou | А землеройка -размерный, древесный докодонтид это известно как одно из первых восхождений на деревья млекопитающие.[36] Он был размером примерно 13 сантиметров от головы до хвоста и весил около 27 граммов. Его внешний вид был похож на белка с длинной мордой, изогнутыми рогатыми когтями и гибкими суставами голеностопного и запястья, типичными для современных древесных пород. млекопитающие. Передние зубы были лопатообразными, что свидетельствовало о том, что Агилодокодон мог прогрызть кору дерева и съесть жевательную резинку или сок. | ||||
А. Дженкинси | Один экземпляр | Древесный, цепкохвостый эухарамиид Харамиидан Это был самый большой из известных харамиидан, который, по оценкам, весил около 354 граммов.[37] Арборохарамия отличается от любого современного млекопитающего тем, что у него нижняя челюсть, которая может двигаться вверх, вниз и назад, но не вперед. Оно имеет грызун зубной ряд с увеличенными резцами и коренными зубами, без клыков. | ||||
С. lutrasimilis | Кровать Daohugou | Один экземпляр[38] | Полуводный докодонтид, который был высокоспециализированным, с адаптациями, которые развивались конвергентно с таковыми у современных полуводных млекопитающих, таких как бобры, выдры, и утконосы. Животное, вероятно, весило около 500-800 граммов (от 1 фунта до почти 2 фунтов) и вырастало как минимум до 42,5 сантиметров (17 дюймов) в длину. Это делает его самым крупным млекопитающим (включая настоящих млекопитающих) юрского периода (предыдущий рекордсмен был Sinoconodon ). | |||
D. brachydactylus | Один экземпляр[39] | Докодонтид специализировался на подземном роющем образе жизни. Строение скелета и пропорции тела поразительно похожи на золотая родинка. Он был не менее 9 сантиметров в длину, не считая хвоста, и весил не менее 9 граммов, а возможно, и 16 граммов. | ||||
Дж. Синенсис[40] | Ляонин | Один экземпляр | Маленький, похожий на землеройку евтерский с длиной тела примерно 70-100 миллиметров. Открытие Джурамая дает новое представление об эволюции плацентарные млекопитающие показав, что их родословная расходится с родословной метатеры На 35 миллионов лет раньше, чем считалось ранее. Кроме того, его открытие заполняет пробелы в летописи окаменелостей и помогает откалибровать современные методы датирования эволюции на основе ДНК. Основываясь на приспособлениях для лазания, обнаруженных в костях передних конечностей, было высказано предположение, что базальный стебель эвтериан был древесным. | |||
| Майопатагиум | М. furculiferum | Планирующий эухарамиидан | ||||
М. mammaliaformis | Внутренняя Монголия | Кровать Daohugou | Один экземпляр | An элеутеродонтид Харамиидан считался травоядным животным, жившим на земле и имевшим осанку, аналогичную современной броненосцы и каменные даманы.[2] Мегаконус по оценкам, весил около 250 граммов (8,8 унции). | ||
| Микродокодон | M. gracilis | Кровать Daohugou | А тегериид докодонтан известен по образцу с сохранившейся подъязычной костью, которая почти не упоминается в летописи окаменелостей ранних млекопитающих. | |||
| Qishou | Q. jizantang | Эухарамиида | ||||
R. eurasiaticus | Ляонин | Даксишанский сайт | Один экземпляр | Всеядный павловоидный многотуберкулезный это самый старый из описанных на данный момент случаев в мульти-туберкулезе. Он сильно напоминал маленького грызуна (как крыса или бурундук ).[41] По оценкам, он весил 65-80 грамм, что примерно соответствует среднему весу бурундука. | ||
V. antiquum | Внутренняя Монголия | Кровать Daohugou | Один экземпляр | Скольжение, белка-летяга -подобно волатикотериан эвтриконодонт со специальной скользящей мембраной. Зубы Волатикотерий были в высшей степени специализированы на поедании насекомых, а его конечности были приспособлены к жизни на деревьях. Скользящая мембрана была изолирована толстым мехом и поддерживалась конечностями и хвостом. Открытие Волатикотерий предоставил самую раннюю известную запись о планирующем млекопитающем и предоставил дополнительные доказательства разнообразия млекопитающих во время Мезозойская эра. | ||
| Вилеволодон | В. дипломатилос | Планирующий элеутеродонтид-харамиидан с травоядной диетой. | ||||




Членистоногие (без учета ракообразные )
В формировании представлены следующие порядки; Ephemeroptera, Одоната, Plecoptera, Blattodea, Прямокрылые, Hemiptera, Neuroptera, Жесткокрылые, Перепончатокрылые, и Двукрылые.
Цветовой ключ
| Примечания Неопределенные или предположительные таксоны входят в маленький текст; |
| Членистоногие пластов Даохугоу | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Состояние | Избыток | Примечания | Изображений | |
А. neimengguensis[42] | ||||||
A. striatus[43] | Арчисаргид мухи | |||||
А. чжанги[44] | Внутренняя Монголия | |||||
А. спуривениус[43] | Арчисаргид летает | |||||
A. strigatus[43] | ||||||
С. (Калосаргус) антиквус[43] | Арчисаргид летает | |||||
С. (С.) Bellus[43] | ||||||
С. (С.) daohugouensis[43] | ||||||
С. (С.) хани[43] | ||||||
С. (С.) tenuicellulatus[43] | ||||||
С. (С.) валидус[43] | ||||||
С. (Птеросарг) синус[43] | Внутренняя Монголия | |||||
D. vulcanica[43] | А лодочник | |||||
F. стадный[43] | А поденок | |||||
Э. Герчи[45] | 1 образец | А plectreurid паук | ||||
Х. Люи[46] | А шизофорид летающий водяной жук | |||||
J. orientalis[43] | Внутренняя Монголия | А Неместриноид летать | ||||
М. объясняющий[46] | Шизофориды летающие водяные жуки | |||||
М. oxycerus[46] | ||||||
M. grammicus[46] | ||||||
М. daohugouensis[43] | Муха архисаргида | |||||
M. portentosus[43] | Внутренняя Монголия | Арчисаргид летает | ||||
M. signatus[43] | Внутренняя Монголия | |||||
M. jurassica[3] | 2 образца | An аранеоморф паук изначально считался разновидностью золотой шелкопрядчик. У самки панцирь составляет 9,31 на 6,83 мм (0,367 на 0,269 дюйма), а опистосома - 15,36 на 9,5 мм (0,605 на 0,374 дюйма). Общая длина тела составляет примерно 24,6 миллиметра (0,97 дюйма), в то время как передние лапы достигают примерно 56,5 миллиметра (2,22 дюйма) в длину. Самец имеет длину тела 16,54 миллиметра (0,651 дюйма) с удлиненными педипальпами. | ||||
S. lacustris[43] | Поденка | |||||
С. дарани[46] | А шизофорид летающий водяной жук | |||||
Другие беспозвоночные
Неопределенный вид эшноидальных насекомых известен из Ляонина.[29]
| Род | Разновидность | Провинция | Стратиграфическое положение | Избыток | Примечания |
|---|---|---|---|---|---|
D. impudica[29] | Ляонин | An остракод | |||
D. magna[29] | Ляонин | Остракод | |||
D. sarytirmenensis[29] | Ляонин | Остракод | |||
С. cliovata[29] | Ляонин |
Флора
Опрос на основе Ванга и другие. 2006 г., если не указано иное.[1]
Bennettitales
Саговидные растения, наиболее многочисленная группа растений в свите. 27 видов в 11 родах.
| Bennettitales формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
Гинкгоалес
Обычные доисторические деревья гинкго, 11 видов, принадлежащих к 6 родам.
| Гинкоалес формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
Pinophyta
Хвойные, 5 видов представлены 4 родами.
| Пинофитаны формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
Pteridophyta
Лептоспорангиатные папоротники, представленные 17 видами в 8 родах, являются вторым по численности типом растений в формации.
| Птеридофитаны формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
Cladophlebis виды | Папоротники | |||||
D. changeyingziensis | ||||||
Д. чариэлса | ||||||
H. shebudaiensis | Нечасто. | |||||
М. hoerenensis | Нечасто. | А мараттиопсид папоротник. | ||||
R. stricta | Папоротник. | |||||
T. denticulata | ||||||
T. williamsonii | ||||||
Другие растения
Cycads, довольно разнообразный, с 10 видами, представленными в 2 родах.
| Cycads формации Тяоцзишань | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Избыток | Примечания | Изображений |
H. shebudaiensis | Нечасто. | А мохообразный. | ||||
Taeniopteris sp. | Нечасто. | |||||
Рекомендации
- ^ а б c Wang, Y .; Ken, S .; Zhang, W .; Чжэн, С. (2006). «Биоразнообразие и палеоклимат среднеюрских флор формации Тяоцзишань в западном Ляонине, Китай». Прогресс естествознания. 16 (1): 222–230. Дои:10.1080/10020070612330087.
- ^ а б Zhou, C. F .; Wu, S .; Martin, T .; Луо, З. X. (2013). «Юрская форма млекопитающих и самые ранние эволюционные адаптации млекопитающих». Природа. 500 (7461): 163–167. Bibcode:2013Натура.500..163Z. Дои:10.1038 / природа12429. PMID 23925238. S2CID 4346751.
- ^ а б Селден, П. А .; Ши, C .; Рен, Д. (2011). "Золотой паук-ткач кругов (Araneae: Nephilidae: Nephila) из средней юры Китая". Письма о биологии. 7 (5): 775–778. Дои:10.1098 / рсбл.2011.0228. ЧВК 3169061. PMID 21508021.
- ^ а б c Ван, Y; и другие. (2006). «Биоразнообразие и палеоклимат среднеюрских флор формации Тяоцзишань в западном Ляонине, Китай». Прогресс естествознания. 16 (S1): 222–30. Дои:10.1080 / 10020070612330087A (неактивно 10.11.2020).CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- ^ Сяолинь Ван; Чжунхэ Чжоу; Хуайюй Хэ; Фан Джин; Юаньцин Ван; Цзянгён Чжан; Юань Ван; Син Сюй; Фучэн Чжан (2005). «Стратиграфия и возраст пласта Даохугоу в Нинчэн, Внутренняя Монголия». Китайский научный бюллетень. 50 (20): 2369–2376. Bibcode:2005ЧСБУ..50.2369Вт. Дои:10.1007 / BF03183749. S2CID 198142479.
- ^ а б Liu, Y .; Liu, Y .; Ji, S .; Ян, З. (2006). «Возраст циркона U-Pb для биоты Даохугоу в Нинчэне Внутренней Монголии и комментарии по связанным вопросам». Китайский научный бюллетень. 51 (21): 2634–2644. Bibcode:2006ЧСБУ..51.2634Л. Дои:10.1007 / s11434-006-2165-2. S2CID 96442710.
- ^ Zhang, H .; Wang, M .; Лю, X. (2008). «Ограничения на возраст верхней границы вулканических пород формации Тяоцзишань в Западном Ляонин-Северном Хэбэе по данным датирования LA-ICP-MS». Китайский научный бюллетень. 53 (22): 3574–3584. Bibcode:2008SciBu..53.3574Z. Дои:10.1007 / s11434-008-0287-4.
- ^ Ren, D .; и другие. (2002). «О биостратиграфии ископаемых пластов юрского периода в Даохугоу около Нинчэна, Внутренняя Монголия». Геол. Бык. Китай. 21: 584–591.
- ^ Он, H .; Ван, X .; Чжоу, З .; Zhu, R .; Jin, F .; Wang, F .; Дин, X .; Бовен, А. (2004). «(^ 40) Ar / (^ 39) Ar датирование игнимбритов из Внутренней Монголии, северо-восток Китая, указывает на постсредний юрский возраст вышележащих слоев Даохугоу». Письма о геофизических исследованиях. 31 (20): L20609. Bibcode:2004GeoRL..3120609H. Дои:10.1029 / 2004gl020792.
- ^ Gao, K .; Рен, Д. (2006). «Радиометрическое датирование игнимбритов из Внутренней Монголии не дает никаких указаний на пост-среднеюрский возраст слоев Даохугоу». Acta Geologica Sinica - английское издание. 80 (1): 42–45. Дои:10.1111 / j.1755-6724.2006.tb00793.x.
- ^ Jin, F .; Zhang, F.C .; Li, Z.H .; Zhang, J.Y .; Li, C .; Чжоу, З.Х. (2008). "На горизонте Протоптерикс и группы окаменелостей ранних позвоночных в биоте Джехол ". Китайский научный бюллетень. 53 (18): 2820–2827. Bibcode:2008SciBu..53.2820J. Дои:10.1007 / s11434-008-0209-5. S2CID 133507763.
- ^ Liu, Y .; Liu, Y .; Чжан, Х. (2006). «LA-ICPMS датирование U-Pb циркона в юрских слоях Даохугоу и соответствующих слоях в Нинчэн во Внутренней Монголии». Acta Geologica Sinica (английское издание). 80 (5): 733–742. Дои:10.1111 / j.1755-6724.2006.tb00296.x.
- ^ а б Gao, K. -Q .; Шубин, Н. Х. (2012). «Позднеюрский саламандроид из западного Ляонина, Китай». Труды Национальной академии наук. 109 (15): 5767–72. Bibcode:2012PNAS..109.5767G. Дои:10.1073 / pnas.1009828109. ЧВК 3326464. PMID 22411790.
- ^ Примечательно Mongolarachne jurassica.
- ^ Тан, Цзинцзин; Рен, Донг; Ши, Чунгкун (2006). «Новые купедиды из средней юры Внутренней Монголии, Китай (Coleoptera: Archostemata)». Annales Zoologici. 56 (1): 1–6.
- ^ Чжан, Куйян; Ян, Дин; Рен, Донг (2006). «Первый бекас (Diptera: Rhagionidae) из средней юры Внутренней Монголии, Китай». Zootaxa. 1134: 51–57. Дои:10.11646 / zootaxa.1134.1.3. S2CID 83904231.
- ^ Люй Цзюньчан и Фуча Сяохуэй (2011). «Новый птерозавр (Pterosauria) из среднеюрской формации Тяоджишань в западном Ляонине, Китай». Глобальная геология Z1: 113–118.
- ^ Лю, Дж. (2009). «Новый нептеродактилоидный птерозавр из округа Цинлун провинции Хэбэй Китая». Acta Geologica Sinica (английское издание). 83 (2): 189–199. Дои:10.1111 / j.1755-6724.2009.00062.x.
- ^ Ченг, Синь; Ван, Сяолинь; Цзян, Шуньсин; Келлнер, Александр В.А. (2014). «Краткая заметка о нептеродактилоидном птерозавре из верхнеюрских отложений Внутренней Монголии, Китай». Историческая биология: международный журнал палеобиологии. 27 (6): 1–6. Дои:10.1080/08912963.2014.974038. S2CID 128454476.
- ^ Lü, J .; Unwin, D.M .; Джин, X .; Liu, Y .; Джи, К. (2010). «Доказательства модульной эволюции длиннохвостого птерозавра с птеродактилоидным черепом». Труды Королевского общества B. 277 (1680): 383–389. Дои:10.1098 / rspb.2009.1603. ЧВК 2842655. PMID 19828548.
- ^ Люй Цзюньчан и Дэвид В.Э. Отточить (2012). "Новый китайский птерозавр анурогнатид и эволюция длины хвоста птерозавров". Acta Geologica Sinica. 86 (6): 1317–1325. Дои:10.1111/1755-6724.12002.
- ^ Lu, J .; Fucha, X .; Чен, Дж. (2010). «Новый скафогнатовый птерозавр из средней юры западного Ляонина, Китай». Acta Geoscientica Sinica. 31 (2): 263–266.
- ^ а б Ван, X .; Чжоу, З. (2006). «Сообщества птерозавров биоты Jehol и их значение для излучения птерозавров раннего мелового периода». Геологический журнал. 41 (3–4): 405–418. Дои:10.1002 / gj.1046.
- ^ Lü, J .; Джи, К. (2006). «Предварительные результаты филогенетического анализа птерозавров из западного Ляонина и окрестностей». Дж. Палеонт. Soc. Корея. 22 (1): 239–261.
- ^ Ченг, Синь; Ван, Сяолинь; Цзян, Шуньсин; Келлнер, Александр В.А. (2012). «Новый скафогнатидный птерозавр из западного Ляонина, Китай». Историческая биология. 24: 101–111. Дои:10.1080/08912963.2011.635423. S2CID 128539358.
- ^ Лю Цзюньчан и Бо Сюэ (2011). «Новый птерозавр рамфоринхид (птерозавр) из среднеюрской формации Тяоджишань в Западном Ляонине, Китай». Acta Geologica Sinica. 85 (5): 977–983. Дои:10.1111 / j.1755-6724.2011.00531.x.
- ^ Лю Дж, Унвин Д.М., Чжао Б., Гао Ц., Шэнь Ц. (2012). «Новый рамфоринхид (Pterosauria: Rhamphorhynchidae) из средней / верхней юры Цинлуна, провинция Хэбэй, Китай» (PDF). Zootaxa. 3158: 1–19. Дои:10.11646 / zootaxa.3158.1.1.
- ^ Ван, X .; Kellner, A.W.A .; Jiang, S .; Мэн, X. (2009). «Необычный длиннохвостый птерозавр с удлиненной шеей из западного Ляонина в Китае». Летопись Бразильской академии наук. 81 (4): 793–812. Дои:10.1590 / с0001-37652009000400016. PMID 19893903.
- ^ а б c d е ж Hu, D .; Hou, L .; Чжан, Л. и Сюй, X. (2009). "ПредварительноАрхеоптерикс Troodontid теропод из Китая с длинными перьями на плюсне ". Природа. 461 (7264): 640–643. Bibcode:2009Натура 461..640H. Дои:10.1038 / природа08322. PMID 19794491. S2CID 205218015.
- ^ Ли Кью, Гао К.К., Винтер Дж. И др. (Март 2010 г.). "Цветовые узоры оперения вымершего динозавра" (PDF). Наука. 327 (5971): 1369–72. Bibcode:2010Sci ... 327.1369L. Дои:10.1126 / science.1186290. PMID 20133521. S2CID 206525132.
- ^ Ху, Дунъюй; Кларк, Джулия А .; Элиасон, Чад М .; Цю, Руи; Ли, Куаньго; Шоуки, Мэтью Д.; Чжао, Цуйлинь; Д'Альба, Лилиана; Цзян, Цзинкай; Сюй, Син (2018). «Юрский динозавр с костистым гребнем и свидетельством радужного оперения подчеркивает сложность ранней паравианской эволюции». Nature Communications. 9 (1): 217. Bibcode:2018НатКо ... 9..217ч. Дои:10.1038 / s41467-017-02515-у. ЧВК 5768872. PMID 29335537.
- ^ Лефевр, Улисс; Кау, Андреа; Cincotta, Aude; Ху, Дунъюй; Чинсами, Анусуя; Эскилье, Франсуа; Годфруа, Паскаль (2017). «Новый теропод юрского периода из Китая документирует переходный этап в макроструктуре перьев». Наука о природе. 104 (9–10): 74. Bibcode:2017SciNa.104 ... 74L. Дои:10.1007 / s00114-017-1496-у. PMID 28831510. S2CID 32780661.
- ^ Сюй, Син; Ты, Хайлу; Ду, Кай; Хан, Фэнлу (28 июля 2011 г.). "An Археоптерикс-подобный теропод из Китая и происхождение Avialae ». Природа. 475 (7357): 465–470. Дои:10.1038 / природа10288. PMID 21796204. S2CID 205225790.
- ^ Сюй, X .; Чжэн, X .; Салливан, К .; Ван, X .; Xing, L .; Wang, Y .; Чжан, X .; о'Коннор, Дж. К .; Zhang, F .; Пан Ю. (2015). «Необычный юрский теропод манирапторана с сохранившимися признаками перепончатых крыльев». Природа. 521 (7550): 70–3. Bibcode:2015Натура.521 ... 70X. Дои:10.1038 / природа14423. PMID 25924069. S2CID 205243599.
- ^ а б Evans, S.E .; Ван, Ю. (2010). «Новая ящерица (Reptilia: Squamata) с превосходной сохранностью мягких тканей из нижнего мела Внутренней Монголии, Китай». Журнал систематической палеонтологии. 8 (1): 81–95. Дои:10.1080/14772010903537773. S2CID 56046960.
- ^ Мэн, Цин-Цзинь; Цзи, Цян; Чжан, Ю-Гуан; Лю, Ди; Гроссникль, Дэвид М .; Ло, Чжэ-Си (2015). «Древесный докодонт юрского периода и экологической диверсификации млекопитающих». Наука. 347 (6223): 764–768. Bibcode:2015Научный ... 347..764M. Дои:10.1126 / science.1260879. PMID 25678661. S2CID 206562565.
- ^ Чжэн, X .; Bi, S .; Ван, X .; Мэн, Дж. (2013). «Новый древесный харамиид показывает разнообразие кроновых млекопитающих юрского периода». Природа. 500 (7461): 199–202. Bibcode:2013Натура.500..199Z. Дои:10.1038 / природа12353. PMID 23925244. S2CID 2164378.
- ^ Meng, J .; Hu, Y .; Li, C .; Ван, Ю. (2006). «Фауна млекопитающих в биоте Джехола раннего мела: значение для разнообразия и биологии мезозойских млекопитающих». Геологический журнал. 41 (3–4): 439–463. Дои:10.1002 / gj.1054.
- ^ Чжэ-Си Ло; Цин-Цзинь Мэн; Цян Цзи; Ди Лю; Ю-Гуан Чжан; Апрель И. Неандер (2015). «Эволюционное развитие базальных форм млекопитающих, выявленное докодонтаном». Наука. 347 (6223): 760–764. Bibcode:2015Sci ... 347..760L. Дои:10.1126 / science.1260880. PMID 25678660. S2CID 206562572.
- ^ Luo, Z .; Юань, C .; Meng, Q .; Джи, К. (2011). «Юрское эвтериальное млекопитающее и расхождение сумчатых и плацентарных». Природа. 476 (7361): 442–445. Bibcode:2011Натура.476..442L. Дои:10.1038 / природа10291. PMID 21866158. S2CID 205225806.
- ^ Yuan, C. -X .; Ji, Q .; Meng, Q. -J .; Табрум, А. Р .; Луо, З. -Х. (2013). «Самая ранняя эволюция многотуберкулезных млекопитающих, обнаруженная по ископаемым останкам нового юрского периода». Наука. 341 (6147): 779–783. Bibcode:2013Научный ... 341..779Y. Дои:10.1126 / science.1237970. PMID 23950536. S2CID 25885140.
- ^ Zhang, K .; Ян, Д .; Ren, D .; Ге, Ф. (2008). «Новые среднеюрские мухи с запутанными жилками из Внутренней Монголии, Китай». Acta Palaeontologica Polonica. 53 (1): 161–164. Дои:10.4202 / приложение.2008.0112.
- ^ а б c d е ж грамм час я j k л м п о п q Чжан, Дж. Ф. (2010). «Записи о необычных юрских брахицерах в биоте Даохугоу, Китай (Diptera, Brachycera, Archisargidae и Rhagionemestriidae)». Палеонтология. 53 (2): 307–317. Дои:10.1111 / j.1475-4983.2010.00934.x.
- ^ Zhang, K .; Ли; Ян, Д .; Рен, Д. (2009). «Новый вид Archirhagio Rohdendorf, 1938 из средней юры Внутренней Монголии Китая (Diptera: Archisargidae)». Zootaxa. 1984: 61–65. Дои:10.11646 / zootaxa.1984.1.4. S2CID 82881259.
- ^ Selden, P.A .; Хуанг, Д. (2010). «Самый старый паук-гаплогин (Araneae: Plectreuridae) из средней юры Китая». Naturwissenschaften. 97 (5): 449–59. Bibcode:2010NW ..... 97..449S. Дои:10.1007 / s00114-010-0649-z. PMID 20140419. S2CID 24576304.
- ^ а б c d е Jarzembowski, E. A .; Ян, Э. В .; Ван, Б .; Чжан, Х.С. (2012). «Новый летающий водяной жук (Coleoptera: Schizophoridae) из юрского лагеря Daohugou lagerstätte». Палеомир. 21 (3–4): 160–166. Дои:10.1016 / j.palwor.2012.09.002.