Птеригот - Pterygotus
| Птеригот | |
|---|---|
 | |
| Окаменелости P. anglicus | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Членистоногие |
| Подтип: | Хелицерата |
| Порядок: | †Eurypterida |
| Надсемейство: | †Pterygotioidea |
| Семья: | †Pterygotidae |
| Род: | †Птеригот Агассис, 1839 г. |
| Типовой вид | |
| †Pterygotus anglicus Агассис, 1849 | |
| Виды | |
17 допустимых видов
6 сомнительных видов
| |
| Синонимы | |
Синонимия рода
Синонимы слова P. anglicus
Синонимы слова P. barrandei
| |
Птеригот это род гигантских хищных эвриптерида, группа вымерших водных членистоногие. Окаменелости Птеригот были обнаружены в отложениях возрастом от Поздний силурийский период к Поздний девон, и были отнесены к нескольким различным видам. Окаменелости были обнаружены с четырех континентов; Австралия, Европа, Северная Америка и Южная Америка, что означает, что Птеригот мог бы иметь почти космополитичный (по всему миру) распространение. Типовой вид, P. anglicus, был описан Швейцарский натуралист Луи Агассис в 1839 году. Агассис ошибочно полагал, что останки принадлежали гигантской рыбе по имени Птеригот переводится как «крылатая рыба». Он осознал ошибку только пять лет спустя, в 1844 году.
Птеригот был одним из самых крупных эвриптерид. Отдельные ископаемые останки большого хелицера (лобный придаток) предполагает, что самый крупный из известных видов, P. grandidentatus, достиг в длину 1,75 метра (5,7 футов). Несколько других видов, особенно P. impacatus на высоте 1,65 м (5,4 фута) и P. anglicus на 1,6 метра (5,2 фута) были столь же гигантскими. Птеригот была превзойдена по размерам другими гигантскими эвриптеридами. Акутирам смог превысить 2 метра (6,6 футов), и Jaekelopterus может достигать 2,6 метра (8,5 футов). Многие виды были значительно меньше самых крупных, например P. kopaninensis на 50 сантиметров (20 дюймов).
Как его близкий родственник Jaekelopterus, Птеригот был крупным и активным хищником, известным своими крепкими и увеличенными хелицерными когтями, которые позволяли ему колоть и хватать добычу, а также Острота зрения (четкость зрения) сопоставима с таковой у современных хищных членистоногих.
Описание
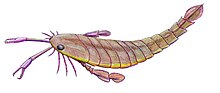
С самыми крупными видами, P. grandidentatus, достигая длины тела 1,75 метра (5,7 футов), Птеригот был одним из самых известных эвриптериды существовать, хотя некоторые из его близких родственников (например, Акутирам и Jaekelopterus ) превзошла его по длине.[1] Хотя было несколько видов-гигантов, многие виды были значительно меньше по размеру. Самый мелкий вид, P. kopaninensisдлиной всего 50 сантиметров (20 дюймов).[2]
Птеригот классифицируется как часть птериготид семейство эвриптерид, которому оно и дало свое название, группа высокообразованных эвриптерид Силурийский к Девонский периоды, которые отличаются от других групп рядом особенностей, возможно, наиболее заметно в хелицеры (первая пара конечностей) и Тельсон. Хелицеры Pterygotidae были увеличены и крепкий, четко адаптированный для использования для активного отлова добычи и более похожий на когти некоторых современных ракообразные, с хорошо развитыми зубцами на коготках, чем хелицеры других групп эвриптерид.[3] В отличие от большей части остальной части тела, которая была покрыта чешуйчатым орнаментом, как у других крылатых эвриптерид, когти лишены какого-либо орнамента.[4] Кроме того, концы когтей были круглыми и изогнутыми, в отличие от острых концов, присутствующих на концах когтей соответствующих Erettopterus.[5] Тельсоны pterygotid были сплющены и расширены, вероятно, использовались в качестве рулей при плавании.[6] Их ходячие ноги были маленькими и тонкими, без шипов, и они, вероятно, не могли ходить по суше.[7][1]
Птеригот отличается от других птериготидов изогнутым дистальный маржа Chelae (когти). В просома (голова) субтрапециевидная (трапеция со скругленными углами), с сложные глаза расположены у края передних углов. Тельсон (самый задний сегмент тела) имеет ярко выраженный спинной киль (или киль), спускающийся по его центру, заканчивающийся коротким шипом.[7]
Размер

Pterygotidae включает в себя самых крупных известных членистоногих, которые когда-либо жили, при этом некоторые виды превышают два метра в длину (например, Jaekelopterus rhenaniae на высоте 2,5 метра (8,2 фута) и Acutiramus bohemicus на высоте 2,1 метра (6,9 футов)).[1] Хотя Птеригот не был самым большим из крылатых, некоторые виды были крупными, превышая 1 метр (3,3 фута) в длину.[2]
Самый крупный из известных видов был P. grandidentatus, с самыми крупными из известных изолированных фрагментов хелицер, предполагающими длину 1,75 метра (5,7 футов). В эстонский P. impacatus является вторым по величине известным видом, самые крупные фрагменты предполагают длину 1,65 метра (5,4 фута).[2] P. anglicus, то типовой вид, выросла до 1,6 метра (5,2 фута) в длину, основываясь на большом тергит обнаружен Генри Вудворд в какой-то момент между 1866 и 1878 годами. Имея размеры чуть более 40 см (16 дюймов) в длину и 10,5 см (4,1 дюйма) в ширину, тергит предполагает наличие эвриптериды с полной длиной 1,6 метра (5,2 фута) от начала панцирь до конца тельсона, если считать расширенные хелицеры (обычно это не так), общая длина превысит 2 метра (6,6 фута).[8] P. carmani, из девона Огайо, вероятно, достигали длины более 1,5 метра (4,9 фута).[4]
Виды П. кобби (1,4 метра (4,6 фута)), P. barrandei (1,26 м (4,1 фута)) и P. denticulatus (1,2 метра (3,9 фута)) также превышал 1 метр в длину. Более мелкие виды включают P. floridanus на 90 см (35 дюймов), П. лайтбоди на 75 см (30 дюймов), P. arcuatus на 60 см (24 дюйма), P. bolivianus 55 сантиметров (22 дюйма) и самый маленький из известных видов, P. kopaninensisдлиной 50 сантиметров (20 дюймов).[2]
История исследования
Первоначальные находки


Первые окаменелости Птеригот были найдены в месторождениях Лохковский -Пражский (Раннедевонский) возраст, установленный карьерами в Шотландии и западной Англии, которые назвали большие ископаемые останки "Серафимы ". Луи Агассис, швейцарско-американский биолог и геолог, описал окаменелости в 1839 году и назвал их Птеригот в убеждении, что это были ископаемые останки большой рыбы; "Птеригот"в переводе" крылатая рыба ".[9] Образцы, описанные Агассисом из Англии, были отнесены к виду, который он назвал Pterygotus problematicus. Агассис впервые узнал истинную природу окаменелостей, поскольку пять лет спустя, в 1844 году, останки членистоногих, после изучения более полных окаменелостей, обнаруженных в Старый красный песчаник Шотландии.[10][11] Признавая окаменелости Птеригот поскольку членистоногие было ближе к своему современному филогенетическому положению, Агассис рассматривал Птеригот представлять ракообразный из Энтомострака подкласс. Несмотря на то что Фредерик Маккой заметил, что Птеригот напоминал Limulidae и ранее обнаруженная эвриптерида Евриптер в 1849 году он классифицировал как Евриптер и Птеригот как ракообразные.[12]
Новые шотландские окаменелости были названы как виды P. anglicus в 1849 г., который остается наиболее широко известным видом Птеригот, отличается от впоследствии обнаруженных видов наличием изогнутых конечных зубов, а также незначительным наклоном молочных и промежуточных зубов назад.[13]
P. problematicus также использовалось в качестве обозначения неполной хелицеры, обнаруженной в Уэльсе на границе западной Англии. Джон Уильям Солтер в 1852 году, но в наше время считается номен ванум («безосновательное название») как вид определить невозможно. Ископаемый материал, с помощью которого он был описан, не является диагностическим и недостаточным для установления каких-либо значимых характеристик, и поэтому многие фрагментарные окаменелости птериготид были отнесены к нему на протяжении всей его долгой истории, что делает его фактически синонимом семейства Pterygotidae. Чаще всего эти фрагменты состоят из пятен покровных покровов крылатых, сохраняющих чешуйчатый орнамент, характерный для группы, которая, как ошибочно полагали исследователи, была характерна только для них. Птеригот или P. problematicus. Поскольку такой орнамент известен у каждого рода pterygotid, он не может использоваться в качестве диагностического признака одного вида. Хотя P. problematicus это самое раннее название вида Птеригот, он не считается типовым видом, так как название больше не используется. Вместо P. anglicus, основанный на ряде диагностических признаков и должным образом проиллюстрированный в своем описании Агассисом в 1844 г., считается типовым видом Птеригот.[11]
Еще два вида, которые до сих пор относятся к этому роду, будут описаны в Англии во время 19 век; P. ludensis из Придоли (Позднесилурийский) возраст и P. arcuatus из Ludlow (Позднесилурийский) возраст, наряду с североамериканским видом, П. кобби, из Придоли США и Канады. P. arcuatus был первоначально описан из синтипический серия (серия экземпляров, из которых голотип не обозначен) ископаемых останков Джона Уильяма Солтера, содержащих пять отдельных ископаемых образцов. В 1961 году, через 102 года после его описания, Эрик Н. Кьеллесвиг-Веринг заметил, что только один из этих экземпляров (номер 89587 в Геологической службе и музее Лондона, свободная ветвь хелицеры) действительно может быть отнесен к этому особому виду как другие окаменелости (тергиты, тазики и неопределимые фрагменты) могут фактически представлять собой окаменелости других видов из-за недостаточной диагностики.[11] P. ludensis, описанный Солтером в 1859 г., отличается от других видов более развитым и удлиненным килем (или гребнем), проходящим по центру тельсона от его начала до хвостового шипа.[14]
Редкий вид П. кобби, описанный Джеймсом Холлом в 1859 году на основе окаменелостей, обнаруженных в Нью-Йорк, США, был первым видом Птеригот должны быть описаны за пределами Шотландии и значительно расширили известный диапазон этого рода. Холл описал этот новый вид вместе с двумя другими североамериканскими видами; P. macrophthalmus (теперь упоминается Акутирам) и P. osborni (позже синонимизированный с участием P. macrophthalmus). В дистальный зуб свободной ветви (движущейся части когтя) был менее заметен, чем у других видов, который был отмечен как сходный с дистальным зубом свободной ветви Акутирам камминси. Несмотря на то что П. кобби основан на бедном ископаемом материале, известном только по свободной ветви, он остается признанным отдельным видом из-за того, что он более похож на определенные виды, обнаруженные в Чешской Республике (например, P. barrandei), чем с другими видами крыловидных, обнаруженными в П. кобби местность (например, Acutiramus macrophthalmus).[5]
Создание подродов и Pterygotidae

К 1859 году к этому роду было отнесено 10 видов,[15] и Джон Уильям Солтер признали, что эти виды можно разделить на подроды на основе морфологии тельсонов. Салтер возвел подрод Птеригот (Erettopterus ) для видов с двулопастным тельсоном.[16] Дальнейшие подроды будут названы по мере того, как было отмечено больше различий между видами, такие подроды включают Птеригот (Курвирам) и Птеригот (Акутирам ), названный в 1935 г. на основании особенностей зубчиков (зубов) хелицер.[16] Птеригот (Курвирам) позже был признан синонимом Птеригот (Птеригот) Лейфом Стёрмером в том же году,[10] и Erettopterus и Акутирам будут признаны отдельными, но близкородственными родами (Erettopterus Эрик Н. Кьеллесвиг-Веринг в 1961 году,[10] и Акутирам Стёрмером в 1974 г.).[16]
В 1912 году семья Pterygotidae был возведен Джон Мейсон Кларк & Рудольф Рюдеманн в 1912 г. включить роды эвриптерид Птеригот, Slimonia, Hughmilleria и Hastimima. Три последних рода будут реклассифицированы как члены Hughmilleriidae Эрик Н. Кьеллесвиг-Веринг в 1951 году, оставив Птеригот и его бывшие подроды как единственные птеригидные эвриптериды.[8]
Дальнейшие открытия

Хотя ранние открытия Птеригот были ограничены Англией и Северной Америкой, находки окаменелостей на протяжении ХХ века помогут установить, что Птеригот как род достиг почти космополитическое распространение.[17] Первая эвриптерида, обнаруженная в Австралия был Pterygotus australis, окаменелости которых были найдены в Мельбурнской группе эпохи Ладлоу Формация Даргайл. Окаменелости, упомянутые P. australis, состоящий из четырех обломков, составляющих примерно половину отрезка, обнаруженных в процессе раскопок под Мельбурн во время строительства новых дренажных сооружений для города в 1899 году. Фрагментарные окаменелости очень напоминают окаменелости Erettopterus bilobus (классифицируется как вид Птеригот в то время), которые могли бы назначить Птеригот под вопросом.[18] В 2020 году вид был отмечен как номен дубиум (сомнительный вид) из-за отсутствия достаточного диагностического материала для разделения P. australis от остальных птериготидов.[19]
Кьеллесвиг-Веринг назвал вид P. bolivianus в 1964 г. по ископаемым останкам, извлеченным из отложений Эмсский -Эйфельян (Ранний-средний девон) возраст в Боливии. Этот вид был первым птериготидом, обнаруженным в Южная Америка, первый девонский птеригитд, обнаруженный в отложениях в Южное полушарие а также представляет собой одного из последних известных ныне живущих членов семейства птеригид. Образец (PE6173, размещенный в Чикагский музей естественной истории ) включает хорошо сохранившуюся переднюю половину хелицеры и ветвь. Зубцы ветви короткие, широкие и конические, все слегка загнуты назад. Конечный зуб больше, но лишь немного больше, чем следующий за ним зуб, и изгиб когтя внутрь предполагает, что может присутствовать другой зуб, создавая своего рода «комбинацию двойных зубов», которая также присутствует у некоторых других видов, таких как П. лайтбоди, P. impacatus и Erettopterus Brodiei. Общая длина окаменевшей ветви составляет 2,35 см (1 дюйм), но, вероятно, она составляет только половину полной ветви. Как и у других видов (и у Pterygotidae в целом), зубы тонко ребристые. Его можно отличить от всех других видов Птеригот широкими, но короткими и одинаковыми по размеру зубами, а также тем, что последний зуб не имеет особого развития.[8] В 2019 г.в области обнаружена новая фрагментарная ветвь хелицеры. Формация Куч из Колумбия. Образец (SGC-MGJRG.2018.I.5), отнесенный с неопределенностью к P. bolivianus из-за сходства со своим голотипом представляет собой первую эвриптериду Колумбии и четвертую в Южной Америке. Окаменелость была датирована Франский (Поздний девон), показывая, что Птеригот не вымерли во время среднего девона, как считалось ранее.[20]
После тщательного изучения и открытия новых свидетельств окаменелостей дальнейшие роды будут отделены от Птеригот. P. rhenaniae был классифицирован как часть своего рода, Jaekelopterus, Чарльз Д. Уотерстон в 1964 году. Он считал этот вид достаточно отличным от других Птеригот вида из-за его предположительно сегментированного генитального придатка (что позже было признано ошибочным), его узких и длинных хелицер и его молочных зубов, слегка наклоненных вперед.[21] Другой вид, П. вентрикоз, был выделен в отдельный род Ciurcopterus в 2007 году О. Эриком Тетли и Дереком Э. Г. Бриггсом, отличавшимся в первую очередь наличием общих черт с более базальными птеритигоидными эвриптеридами, такими как его придатки, похожие на таковые Slimonia.[22]
Открытия в Европе

Новые находки окаменелостей также выявили присутствие Птеригот в нескольких европейских странах, где он ранее был неизвестен, и установили его как высоко таксономически разнообразный род. P. barrandei был назван в 1898 году и имеет ископаемое представление в возрастных отложениях Придоли Чехия. P. barrandei отмечается, что очень похож на П. кобби, и предполагается тесная связь между двумя видами. Несмотря на многие сходства, у этих двух видов есть некоторые различия, наиболее заметно в хелицеральных зубах свободных ветвей. Самый крупный зуб свободной ветви P. barrandei значительно длиннее, чем соответствующий зуб в П. кобби и зубы свободной ветви P. barrandei в целом направлены вперед более заметно. Окаменелости P. barrandei редки, ископаемые находки приурочены к нескольким образованиям возраста Придоли в Богемия. Известные окаменелости включают некоторые неполные хелицеры и метастомы. К этому виду были отнесены некоторые дополнительные ископаемые останки, состоящие из тазиков и генитального придатка, но их отнесение к этому виду сомнительно.[23]
Виды P. nobilis, описанный в 1872 году, основан на небольшой фрагментарной хелицере, обнаруженной в том, что сегодня является Чехия. Расположение зубов, которое можно увидеть на этой клешне, хотя большинство зубов не сохранилось, было отмечено исследователем Максом Семпером в 1897 году как имеющее мало или совсем не сходное с тем, что наблюдается у типовых видов животных. Птеригот, P. anglicus, и было отмечено, что на самом деле он больше похож на то, что видно на P. buffaloensis и P. bohemicus. сегодня P. buffaloensis считается младшим синонимом P. bohemicus, который был реклассифицирован как часть близкородственного рода Акутирам.[16] Фрагментарные останки P. nobilis затрудняет дальнейшие исследования его точной идентичности, Семпер предположил, что это может быть синонимом Acutiramus bohemicus, но отметил, что на «вопросы [о его личности] невозможно ответить из доступных мне материалов».[24]
Другой вид, P. kopaninensis, также названный в 1872 г., известен по единичной и неполной фиксированной ветви хелицеральной ветви (номер экземпляра L1396), извлеченной из Копанинская свита вокруг села Задняя Копанина, находится в Прага. Образец имеет длину 4,3 см (1,7 дюйма) и в одной точке был отнесен к Erettopterus из-за «своеобразного крючковидного окончания ветви», особенности, которая, как теперь известно, присутствует из-за остатков свободной ветви, присутствующих в ископаемом остатке. Рисунок зубов и форма когтя подсказывают, что назначение на Птеригот скорее всего.[23]
П. семирадский, описанный Эмбрик Стрэнд в 1926 г. на основе ископаемого материала из западных Украина, основан на очень фрагментарном материале, имеющем небольшую диагностическую ценность. Единственный образец, обозначенный как П. семирадский, тельсон, имеет необычные и ярко выраженные гребни, которых нет ни у одного известного вида Птеригот, и ни в каком другом роде pterygotid eurypterids, что делает его отнесение к Птеригот под вопросом.[25]
В 1964 году два вида, описанные Кьеллесвиг-Верингом, увеличили известный ареал Птеригот к Шотландия (P. lanarkensis) и Эстония (P. impacatus), оба по возрасту Ладлоу.[2][8] P. lanarkensis был вторым птеригидом, обнаруженным в хорошо известной фауне эвриптерид Лесмахагоу в Ланаркшир, Шотландия. Поскольку птериготиды обычно встречаются вместе с несколькими родственными родами, считалось необычным, что существует только один вид, Erettopterus bilobus, присутствует в Лесмахагове. Ископаемые останки P. lanarkensis были известны с 1868 г. (впервые собраны Робертом Слимоном в 1855-1860 гг.), но впервые признаны таковыми Кьеллесвиг-Верингом в 1964 г. Представлен большой хелицерой (номер экземпляра 53890 в Британский музей естественной истории ) изначально сомнительно относился к Erettopterus bilobus, P. lanarkensis больше напоминает P. anglicus чем это делает виды Erettopterus. Конечный зуб (на этом экземпляре сломан) имеет длину 0,9 см, а центральный зуб необычно короткий. Сочетание необычно длинного конечного зуба и недоразвитого центрального зуба отличает P. lanarkensis из других видов Птеригот. Несколько особенностей отличают его от Erettopterus с которыми он был обнаружен, в том числе с более прочными хелицерами P. lanarkensis.[8]
P. impacatus, извлечены из отложений возраста Ладлоу на Кильконд в Сааремаа, Эстония, представлен голотипом, состоящим из фрагментарной свободной ветви хелицеры, сохраняющей некоторые диагностические и хорошо сохранившиеся детали (экземпляр № 7059/7, помещенный в Музей сравнительной зоологии ). Помимо этого экземпляра, важные экземпляры, относящиеся к этому виду, включают паратип (№ 7059/3, фрагментарные хелицеры). Этот образец включает несколько функций, которые позволяют диагностировать P. impacatus, например, прямые зубы, следующие за толстыми и длинными зубцами конечной части когтя. Особенно важное диагностическое значение имеет то, что зубы присутствуют в том месте, где первые зубы начинаются. Центральный зуб свободной ветви у этого вида необычно большой. Конечный зуб имеет длину 2,3 см (1 дюйм). Рами P. impacatus украшены большими заостренными пустулами (возвышениями на коже), и это помогает различать экземпляры P. impacatus из других птериготидов в ископаемых местах, где были найдены его останки.[8] В 2007 году О. Эрик Тетли поставил под сомнение назначение P. impacatus к Птеригот поскольку форма глаз и панциря была похожа на форму этих частей тела в Erettopterus.[16]
Англия

Англия, место первого открытия P. problematicus, предоставил окаменелости еще нескольким видам. Кьеллесвиг-Веринг назвал три новых вида из Англии в 1961 году; P. denticulatus, П. лайтбоди (оба в возрасте позднего Ладлоу) и P. grandidentatus (Венлок, Позднесилурийский по возрасту).[2] P. denticulatus Известен по единственному экземпляру (№ 48393 Британского музея естественной истории), включает около половины переднего конца того, что предположительно является неподвижной ветвью хелицеры. Коготь крепкий, с необычно короткими зубцами, слегка ребристыми. Эта частичная ветвь имеет длину 1,4 см (0,5 дюйма) и была обнаружена в Whitcliffe в Шропшир, Англия связана с окаменелостями брахиопод и головоногих моллюсков. Хотя иногда это считается синонимом П. лайтбоди, P. denticulatus можно отличить по маленьким, толстым и изогнутым зубчикам когтей, отличающимся не только от П. лайтбоди в этом отношении, но практически все другие виды Птеригот также.[11]
П. лайтбоди назван в честь Роберт Лайтбоди, британский геолог-любитель, внесший ценный вклад в палеонтологические исследования раннего палеозоя Валлийский пограничье, включая открытие важных окаменелостей силурия (таких как эвриптериды) в 1800-х годах. Этот вид был одним из самых распространенных эвриптерид в Англии в эпоху Ладлоу, был довольно большим и четко отличался (хотя и напоминал P. barrandei, P. floridanus и П. кобби по своей хелицеральной морфологии) от других видов рода, известных по множеству экземпляров. Наиболее важные окаменелости П. лайтбоди включают голотип (состоящий из большей части хелицер) и два паратипа (включая большую часть свободной ветви). Когти П. лайтбоди все они оснащены вертикально расположенными и очень длинными зубцами, большинство из которых слегка загнуты назад. Конечный зуб необычайно тонкий и длинный в П. лайтбоди, и, как и в случае с другими зубами, слегка загнуты назад. Среди наиболее важных диагностических признаков этого вида - сочетание большого конечного зуба и большого вертикального зуба рядом с ним.[11]
P. grandidentatus известен по единственному экземпляру, передняя половина свободной ветви хелицеры, обнаруженная в Венлок кровати на Дадли в Вустершир, Англия (экз. № I. 3163 в г. Британский музей естественной истории ). Он примечателен крепким стержнем и необычно длинной (1,75 см, 0,7 дюйма) конечного зуба и необычной толщиной его основания. P. grandidentatus Его легко отличить от других видов не только по необычному концевому зубу, но и по дезориентированным зубам вдоль когтя, изогнутым в разных направлениях. Терминальная часть ветви заканчивается расположением нескольких зубов, иначе наблюдаемых только у видов. П. waylandsmithi.[11] Поскольку эта специфическая часть ветвления мало известна от других видов Птеригот и П. waylandsmithi был реклассифицирован как вид Erettopterus в 2007 году назначение P. grandidentatus к Птеригот сомнительно.[16]
Англия также даст сомнительный вид, P. taurinus, из месторождений Придоли[2] или девонский[11] возраст. Названный Солтером в 1868 году, P. taurinus рассматривается как сомнительный вид по той причине, что он фактически представляет собой смесь, состоящую из окаменелостей трех различных эвриптерид. Эти окаменелости состоят из типового образца хелицер (который сейчас утерян, что усложняет любые возможные сравнения), большого панциря и хелицер (вероятно, фактически относящихся к Птеригот из-за того, что он похож на P. anglicus), нога (несомненно, представляющая собой карцинозоматид эвриптерида, потенциально Карциносома ) и тельсон (который гораздо больше похож на Erettopterus чем Птеригот). Большинство этих образцов было потеряно с 1870-х годов, последняя запись гласит, что все они находились в кабинете доктора Маккалоу из Абергавенни.[11]
Открытия в Северной Америке

В 20-м веке появятся описания дополнительных видов Птеригот в Северной Америке, включая силурийский P. marylandicus (1964, Мэриленд, США) и P. monroensis (1902, Нью-Йорк, США), ранний девон P. carmani (1961, США) и P. floridanus (1950, Флорида, США) и средний девон P. gaspesiensis (1953, Квебек, Канада ).[2][26]
Ископаемые останки птеригидных эвриптерид с отчетливым чешуйчатым орнаментом, известным из этой группы, были обнаружены в восточной части Канады еще в 1846 году, когда исследователь Уильям Эдмонд Логан сообщил о появлении животного, "очень похожего на животное Мерчисона. Pterygotus problematicus"в силурийско-девонских отложениях Гаспе, Квебек. Окаменелости, в конечном итоге идентифицированные как исключительно девонского возраста, сначала были предварительно отнесены к P. atlanticus (теперь синоним P. anglicus), которые были обнаружены в относительно непосредственной близости от окаменелостей Гаспе, благодаря P. atlanticus материал был настолько фрагментарен, что было невозможно сказать, представляют ли они один и тот же вид. Они были описаны Лорисом С. Расселом как принадлежащие к новому виду. P. gaspesiensis в 1953 г.[27]
Джон Уильям Доусон в 1861 г. назвал новый вид ликопод завод, Selaginites formosusна основании предполагаемых останков стволов и ветвей, найденных в Гаспе. Солтер убедил Доусона, что окаменелости S. formosus на самом деле были фрагментарными останками эвриптериды, что подтверждается дальнейшими находками.[28][29] Доусон реклассифицировал его как эвриптериду в 1871 году. Кьеллесвиг-Веринг в 1964 году определил его как сомнительный вид Птеригот.[10]
В 1921 году Рюдеманн описал фауну эвриптерид из Формация Вернон из Питтсфорд, Нью-Йорк. Среди них виды P. vernonensis возводился на основе двух небольших коротких панцирей. Контур и положение глаз позволяют предположить принадлежность к роду. Птеригот, в отличие от P. monroensis в почти прямоугольной форме и с прямым поперечный лобный край. Он предложил отношения с Slimonia, но он не назначил его из-за отсутствия дополнительных материалов, свидетельствующих о последнем.[30] Хотя позже он был отнесен к роду Waeringopterus, Сэмюэл Дж. Чурка-младший и О. Эрик Тетли пришли к выводу в 2007 году, что у голотипа на самом деле нет глаз и он представляет собой не что иное, как неполный сегмент тела. Поэтому они рассматривали этот вид как номен дубиум.[16]
P. floridanus, извлеченные из отложений лохковского возраста во Флориде, расширили известный ареал эвриптерид на континенте более чем на 800 км (500 миль) к югу. До открытия эвриптериды в Северной Америке были известны только из северных частей континента, а штат Нью-Йорк представлял собой штат, наиболее богатый ископаемыми. Остатки P. floridanus были впервые обнаружены Г. Артур Купер в Округ Суванни, Флорида, и окаменелости состоят из фиксированной ветви хелицер, а также фрагментов брюшных пластин и тергитов и, как было установлено, представляют новый вид Птеригот Эрик Н. Кьеллесвиг-Варинг в 1950 году. Он больше всего напоминает П. кобби и P. barrandei, в отличие от П. кобби в его более развитом центральном зубе и расположении более мелких зубцов когтей и от P. barrandei в этом P. floridanus имеет более тонкую ветвь.[31]
P. carmani наиболее часто встречающаяся эвриптерида в нижнедевонских отложениях Лукас Каунти, Огайо. Описан Эриком Н. Кьеллесвиг-Верингом в 1961 году и назван в честь доктора Дж. Эрнеста Кармана из Университета штата Огайо, первым обнаружившим эвриптериды в местности, где находились P. carmani останки были найдены, P. carmani в первую очередь известен по неполным хелицерам и гнатобазам тазиков. Известные ископаемые останки, помимо двух тазиков и трех хелицер в первоначальном описании, также включают метастому и претельсона. Все эти оригинальные ископаемые образцы были обозначены Кьеллесвиг-Верингом как паратип экземпляры по первоначальному описанию вида. Обозначенный типовой образец - неполная хелицера, PE5105, которая остается хранящейся в Чикагском музее естественной истории вместе с паратипными образцами. Вид можно отличить от других видов Птеригот в первую очередь особенностями его хелицеральных зубов, отличных от P. barrandei и П. кобби эти зубы менее развиты и толще P. carmani а также зубы, имеющие заметно иное расположение на когтях.
P. marylandicus, от депозитов Лудфордский (Позднесилурийский) возраст, известен по фрагментарному и небольшому телсону из Формация Маккензи, Штат Мэриленд, впервые описанный Кьеллесвиг-Верингом в 1964 году, который распознал его как тельсон Птеригот. Образец (№ 140901 в г. Национальный музей США ) очень широкая, 0,75 см, с почти прямым основанием с сходящимися кпереди краями. В отличие от некоторых видов, на телсоне нет зазубрин, а позвоночник тупой. Этот вид очень отличается от всех других силурийских видов Птеригот по форме тельсона. Вид Jaekelopterus, Дж. Хауэлли из раннего девона похож на широкую и усеченную форму тельсона, но его легко отличить по зубцам и гораздо большему конечному шипу.[8]
Виды P. monroensis, известный по месторождениям от позднего Венлока до Ладлоу в штате Нью-Йорк, США, был предложен как синоним Erettopterus osiliensis Сэмюэл Дж. Чурка-младший и О. Эрик Тетли в 2007 году, основываясь на схожей форме глаз и панциря. Такое изменение будет иметь последствия для других видов Птеригот а также с P. impacatus потенциально также представляет собой синоним E. osiliensis.[16] Последующие исследования и списки видов эвриптерид продолжали лечить P. monroensis и P. impacatus как отдельные виды Птеригот.[15]
Классификация


Птеригот относится к семье Pterygotidae в надсемействе Pterygotioidea,[15] давая свое имя как своей семье, так и надсемейству. Три наиболее производных птеригидных эвриптериды, Акутирам, Jaekelopterus и Птеригот, очень похожи друг на друга. Птеригот особенно похож на Jaekelopterus, от которого он фактически отличается только чертами генитального придатка и, возможно, тельсона.[32]
Сходство генитального придатка может означать, что все три рода являются синонимами друг друга, поскольку они были классифицированы в прошлом (как виды Птеригот). Некоторые различия между ними также были отмечены в хелицерах, хотя хелицеры были подвергнуты сомнению как основа родового различия эвриптерид, поскольку их морфология зависит от образа жизни и, как было замечено, варьировалась на протяжении всей жизни. онтогенез. Морфология Тельсона иногда используется в качестве отличительного признака, хотя все тельсоны трех производных родов птериготид имеют лопастную форму (тельсон из Jaekelopterus имеет треугольную форму, но может попадать в морфологический диапазон других родов).[33] Инклюзивный филогенетический анализ с несколькими видами Акутирам, Птеригот и Jaekelopterus требуется, чтобы решить, являются ли роды синонимами друг друга.[33]
Приведенная ниже кладограмма основана на девяти наиболее известных видах птеригид и двух таксонах внешней группы (Slimonia acuminata и Hughmilleria socialis ). Кладограмма также содержит максимальные размеры, достигаемые рассматриваемым видом, что, как предполагалось, могло быть эволюционной чертой данной группы на Правило Коупа («филетический гигантизм»).[1][34]
| Pterygotioidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология

Морфология хелицер и Острота зрения птеригидных эвриптерид разделяет их на отдельные экологические группы. Основным методом определения остроты зрения у членистоногих является определение количества линз на их теле. сложные глаза и межоматидиальный угол (сокращенный как IOA и относящийся к углу между оптическими осями соседних линз). IOA особенно важен, поскольку его можно использовать для различения различных экологических ролей у членистоногих, поскольку он невелик у современных активных хищников членистоногих.[35]
И то и другое Pterygotus anglicus и Jaekelopterus rhenaniae обладали очень высокой остротой зрения, которую исследователи могли определить, наблюдая низкий IOA и большое количество линз в их сложных глазах. В хелицеры из Птеригот были увеличены, прочны и обладали изогнутой свободной ветвью и зубчиками разной длины и размера, все приспособления, которые соответствуют сильным прокалывающим и захватывающим способностям в существующих скорпионы и ракообразные. Значения IOA для обоих Птеригот и Jaekelopterus совпадают с таковыми у современных активных хищных членистоногих высокого уровня, указывая на то, что они представляли визуальных и активных хищников.[35]
Все известные птериготиды (хотя они пока неизвестны в Ciurcopterus[22]) обладал хелицерными когтями. Первый сустав хелицер, где он соединяется с эпистомой (пластинка, расположенная на просома, или «голова»), была бы способна поворачивать весь придаток скручивающимся образом, что привело исследователей к выводу, что функция хелицер заключалась не только или даже в первую очередь в защите, а скорее в захвате и переносить пищу в рот. Когда добычу поймают, ее нужно будет разбить на более мелкие части, чтобы она могла уместиться в пасти; Рты эвриптерид были даже менее приспособлены к пожиранию больших кусков, чем рты современных крабы находятся. Ходячие придатки эвриптерид не могли ничего разрезать, транспортировать или захватывать, поэтому эти задачи, скорее всего, также выпадут на хелицеры. У крабов клешни разрывают пищу, а затем переносят более мелкие кусочки в рот. Основываясь на процессе питания, наблюдаемом у современных членистоногих с хелицерами, один из когтей будет удерживать добычу, а другой отрезать кусочки и переносить ее ко рту непрерывными и простыми движениями.[8]
Палеоэкология

подобно Акутирам, Птеригот жили в полностью морской среде, и сегодня обнаружены его окаменелости, связанные с обычными и разнообразными окаменелостями, указывающими на такую среду.[36] В частности, P. barrandei и П. кобби встречаются в морской среде, связанной с окаменелостями Акутирам. Птеригот был более распространен, чем Акутирам и поэтому многие виды встречаются без ассоциированных представителей Акутирам а также такие как британские П. лайтбоди.[11] Жилая среда P. australis, тоже морской, находился на особо глубокой воде.[37] Единственные известные ископаемые организмы того же возраста и места, что и P. australis находятся морские лилии рода Дендрокринус и Kooptoonocrinus и офиоуроиды рода Protaster.[38]
Месторождения, где найден голотипный экземпляр P. kopaninensis были найдены запасы окаменелостей и многих других животных. Среди них есть трилобиты такие как Леонаспис, Raphiophorus и фрагментарный гарпетид трилобит. Также сохранились двустворчатые моллюски, такие как Кардиола, и граптолиты, такие как Monograptus.[23] Птеригот чаще всего встречается в экосистемах с разнообразной фауной эвриптерид, P. lanarkensis из Формация Кипа Бёрна Шотландии встречается вместе с несколькими другими родами эвриптерид; Slimonia, Eusarcana, Nanahughmilleria, Parastylonurus, Erettopterus и Карциносома.[39] P. impacatus из Эстонии встречается вместе с родами Erettopterus, Erieopterus, Карциносома, Mixopterus, Долихоптер и Eysyslopterus а также цефаласпидоморфных рыб.[40] P. monroensis жил рядом Mixopterus, Hughmilleria, Евриптер и Карциносома.[41] P. nobilis жили рядом с представителями Акутирам, Erettopterus и Eusarcana а также с разнообразной фауной конодонты, брюхоногие моллюски, головоногие моллюски, остракоды, малакостраки, трилобиты и двустворчатые моллюски и хрящевые рыбы Онхус.[42]
В раннем девоне Формация Данди Шотландии, окаменелости P. anglicus встречаются вместе с широким спектром различных эвриптерид, включая Erieopterus, Pagea, Тарзоптерелла и Stylonurus, а также акантод рыбы Ишнакантус и Мезакантус, акула Климатий и остеостракий рыбы Цефаласпис. Окаменелости P. carmani того же возраста, найденного в Сланцевый карьер Голландии из Огайо также сохраняют разнообразную девонскую фауну различных эвриптерид, таких как Долихоптер, Стробилоптер и Erettopterus, а также птераспидоморф рыбы Аллокритаспис и плакодерма рыбы Этапсис.[39] Колотые раны на ископаемом пораспид рыба, относящаяся к родственной птеригид Jaekelopterus продемонстрировать, что птериготиды были способны охотиться на бронированных рыб, таких как птераспидоморфы и плакодермы.[35] Отложения, которые дали окаменелости П. семирадский также дали окаменелости Евриптер и несколько родов рыб птераспидоморф; Ларноваспис, Фиаласпис, Корваспис и Подоласпис.[43][44]
Пять видов Птеригот были обнаружены в отложениях силурийского возраста в Валлийский пограничье, P. ludensis, P. arcuatus, P. grandidentatus, П. лайтбоди и P. denticulatus. Валлийская приграничная территория сохраняет одну из наиболее обширных известных фаун эвриптерид с точными родами и видами в зависимости от точного времени. В возрасте Венлока P. grandidentatus произошло вместе с Mixopterus, P. arcuatus возраста Среднего Ладлоу произошли вместе с Salteropterus, Erettopterus и Карциносома пока П. лайтбоди и P. denticulatus были современниками друг друга и виды родов Erettopterus и Карциносома. P. ludensis, По возрасту даунтонский, возник вместе с разнообразным множеством эвриптерид, состоящих из Карциносома, Долихоптер, Erettopterus, Hughmilleria, Parahughmilleria, Евриптер, Nanahughmilleria, Марсупиптер, Stylonurus, Тарзоптерелла, Slimonia и Salteropterus.[11]
Иногда единственные известные ископаемые эвриптериды, встречающиеся с Птеригот также являются птериготами. P. marylandicus происходит вместе только с Erettopterus.[45] Сомнительный P. taurinus из Англии был найден в месторождениях, где также были обнаружены остатки Erettopterus а также разнообразную рыбу, такую как акантоды, телодонтиформы а также цефаласпидоморфы Гемицикласпис и Thyestes.[46]
В некоторых случаях, Птеригот представляет собой единственную известную эвриптериду в своей среде обитания. P. bolivianus был обнаружен ассоциированным только с трилобитами рода Метакрифы в своем типе местности.[8] P. floridanus был обнаружен в среде, которая, как известно, включала брюхоногие моллюски (такие как Плектонотус ), остракоды, пелециподы, тентакулиты (такие как Тентакулиты ) и разнообразная фауна двустворчатых моллюсков (в том числе Эошизодус, Pleurodapis, Арисайга, Птеринопектен и Modiomorpha ).[47] P. gaspesiensis из Канады был извлечен из среды обитания различных видов двустворчатых моллюсков и брюхоногих моллюсков, а также трилобитов. Факопс и малакостракан Тропидокарис, но никакие другие известные эвриптериды.[48]
Смотрите также
использованная литература
- ^ а б c d Брэдди, Саймон Дж .; Пошманн, Маркус; Тетли, О. Эрик (2007). «Гигантский коготь показывает самого большого членистоногого». Письма о биологии. 4 (1): 106–109. Дои:10.1098 / rsbl.2007.0491. ЧВК 2412931. PMID 18029297.
- ^ а б c d е ж г час Ламсделл, Джеймс С.; Брэдди, Саймон Дж. (2009-10-14). "Правило Копа и теория Ромера: закономерности разнообразия и гигантизма у эвриптерид и палеозойских позвоночных ». Письма о биологии: rsbl20090700. Дои: 10.1098 / rsbl.2009.0700. ISSN 1744-9561. PMID 19828493. Дополнительная информация В архиве 2018-02-28 в Wayback Machine
- ^ Тетли, О. Эрик (2007). «История распространения и расселения Eurypterida (Chelicerata)» (PDF). Палеогеография, палеоклиматология, палеоэкология. 252 (3–4): 557–574. Дои:10.1016 / j.palaeo.2007.05.011. Архивировано из оригинал (PDF) на 2011-07-18.
- ^ а б Кьеллесвиг-Веринг, Эрик Н. (1961). «Эвриптериды девонских сланцев Голландского карьера Огайо». Геология Fieldiana. 14: 79–98.
- ^ а б Кларк, Джон Мейсон (1912). "Евриптерида Нью-Йорка". Мемуары (Музей и научная служба штата Нью-Йорк). 14.
- ^ Плотник, Рой Э .; Баумиллер, Томаш К. (1988). «Птеригид телсон как биологический руль направления». Lethaia. 21 (1): 13–27. Дои:10.1111 / j.1502-3931.1988.tb01746.x.
- ^ а б Стёрмер, Лейф (1955). «Меростомы». Часть P Arthropoda 2, Chelicerata. Трактат по палеонтологии беспозвоночных. С. 30–31.
- ^ а б c d е ж г час я Кьеллесвиг-Веринг, Эрик Н. (1964). "Синопсис семьи Pterygotidae Clarke и Ruedemann, 1912 (Eurypterida)". Журнал палеонтологии. 38 (2): 331–361. JSTOR 1301554.
- ^ Мерчисон, Родерик Импи (1839). Силурийская система, основанная на геологических исследованиях в графствах Салоп, Херефорд, Рэднор, Монтгомери, Кармартен, Брекон, Пембрук, Монмут, Глостер, Вустер и Стаффорд: с описаниями угольных месторождений и вышележащих формаций. Albemarle Street. п. 606.
крылатый крылатый одно имя.
- ^ а б c d Кьеллесвиг-Веринг, Эрик Н. (1964). "Краткое изложение семейства Pterygotidae Clarke и Ruedemann, 1912 (Eurypterida)". Журнал палеонтологии. 38 (2): 331–361. JSTOR 1301554.
- ^ а б c d е ж г час я j Кьеллесвиг-Веринг, Эрик Н. (1961). «Силурийская Eurypterida валлийского пограничья». Журнал палеонтологии. 35 (4): 789–835. JSTOR 1301214.
- ^ Маккой, Фредерик (2009). «XLI. - О классификации некоторых британских ископаемых ракообразных с уведомлениями о новых формах в университетской коллекции в Кембридже». Летопись и журнал естественной истории. 4 (24): 392–414. Дои:10.1080/03745486009494858.
- ^ Миллер, Рэндалл Ф. (2007). «Pterygotus anglicus Agassiz (Chelicerata: Eurypterida) из Атолвилля, формация Кэмпбеллтаун нижнего девона, Нью-Брансуик, Канада». Палеонтология. 50 (4): 981–999. Дои:10.1111 / j.1475-4983.2007.00683.x. ISSN 1475-4983.
- ^ Локвуд, S (1870). "Конский краб". Американский натуралист. 4 (5): 257–274. Дои:10.1086/270576.
- ^ а б c Данлоп, Дж. А., Пенни, Д., Джекель, Д. 2018. Сводный список ископаемых пауков и их родственников. В мировом каталоге пауков. Музей естественной истории Берн
- ^ а б c d е ж г час Ciurca, Samuel J .; Тетли, О. Эрик (2007). "Pterygotids (Chelicerata; Eurypterida) из силурийской формации Вернон в Нью-Йорке". Журнал палеонтологии. 81 (4): 725–736. Дои:10.1666 / pleo0022-3360 (2007) 081 [0725: PEFTSV] 2.0.CO; 2. ISSN 0022-3360.
- ^ Тетли, О. Эрик (2007). «История распространения и расселения Eurypterida (Chelicerata)». Палеогеография, палеоклиматология, палеоэкология. 252 (3–4): 557–574. Дои:10.1016 / j.palaeo.2007.05.011. ISSN 0031-0182.
- ^ Маккой, Фредерик (1899). "I. - Заметка о новом австралийском птериготе". Геологический журнал. 6 (5): 193–194. Bibcode:1899ГеоМ .... 6..193М. Дои:10.1017 / S0016756800143249. ISSN 1469-5081.
- ^ Бикнелл, Рассел Д. С.; Смит, Патрик М .; Пошманн, Маркус (2020). «Переоценка свидетельств австралийских эвриптерид». Исследования Гондваны. 86: 164–181. Дои:10.1016 / j.gr.2020.06.002.
- ^ Олив, Себастьян; Прадель, Алан; Мартинес-Перес, Карлос; Жанвье, Филипп; Ламсделл, Джеймс С.; Герио, Пьер; Рабе, Николас; Дюранло-Ганьон, Филипп; Карденас-Розо, Андрес Л .; Сапата Рамирес, Паула А .; Ботелла, Эктор (2019). «Новые сведения о позднедевонских позвоночных и связанной с ними фауне из формации Куч (массив Флореста, Колумбия)». Журнал палеонтологии позвоночных. 39 (3): e1620247. Дои:10.1080/02724634.2019.1620247. S2CID 198237241.
- ^ Д. Уотерстон, Чарльз (1964). "II. — Наблюдения за птеригидными эвриптеридами". Сделки Королевского общества Эдинбурга. 66 (2): 9–33. Дои:10.1017 / S0080456800023309.
- ^ а б Тетли, О. Эрик; Бриггс, Дерек Э. Г. (2009). «Происхождение птеригидных эвриптерид (Chelicerata: Eurypterida)». Палеонтология. 52 (5): 1141–1148. Дои:10.1111 / j.1475-4983.2009.00907.x. ISSN 1475-4983.
- ^ а б c Хлупак, Иво (1994). "Эвриптериды Pterygotid (Arthropoda, Chelicerata) в силурии и девоне Богемии". Журнал Чешского геологического общества. 39 (2–3). ISSN 1802-6222.
- ^ Семпер, Макс (1897). "Die Gigantostraken des Älteren Böhmischen Palaeozoicum" (PDF). Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients.
- ^ Данлоп, Джейсон; Эрик Тетли, О (2006). "Эвриптериды Эмбрика Стрэнда". Neues Jahrbuch für Geologie und Paläontologie - Monatshefte. 2006 (11): 696–704. Дои:10.1127 / njgpm / 2006/2006/696.
- ^ Гриффинг, Дэвид Х .; Мост, Джон С .; Хоттон, Кэрол Л. (2000). «Прибрежно-речные палеоокружения и палеоэкология растений нижнего девона (эмс), залив Гаспе, Квебек, Канада». Геологическое общество, Лондон, Специальные публикации. 180 (1): 61–84. Дои:10.1144 / GSL.SP.2000.180.01.05. ISSN 0305-8719. S2CID 129630806.
- ^ Ф. Миллер, Рэндалл (2007). «Pterygotus anglicus Agassiz (Chelicerata: Eurypterida) из Атолвилля, формация Кэмпбеллтон нижнего девона, Нью-Брансуик, Канада». Палеонтология. 50 (4): 981–999. Дои:10.1111 / j.1475-4983.2007.00683.x.
- ^ Доусон, Джон В. (1871). «Предкаменноугольные растения». Ископаемые растения девонских и верхнесилурийских отложений Канады. Геологическая служба Канады. п. 65. Дои:10.5962 / bhl.title.38239. ISBN 978-0665059063.
- ^ Уайтавс, Дж. Ф. (1899). «Девонская система в Канаде». Наука. Новая серия. 10 (247): 402–412. Дои:10.1126 / science.10.247.402. JSTOR 1625781. PMID 17751574.
- ^ Рудеманн, Рудольф (1921). «Повторяющаяся фауна Питтсфорда (Салина)». Бюллетень Музея штата Нью-Йорк: 205–222.
- ^ Кьеллесвиг-Веринг, Эрик Н. (1950). «Новая силурийская эвриптерида из Флориды». Журнал палеонтологии. 24 (2): 229–231. JSTOR 1299504.
- ^ Кайзер, Александр; Клок, Хако (2008). «Означают ли гигантские когти гигантские тела? Альтернативный взгляд на преувеличенные отношения масштабирования». Письма о биологии. 4 (3): 279–280. Дои:10.1098 / рсбл.2008.0015. ЧВК 2610042. PMID 18353748.
- ^ а б Ламсделл, Джеймс С.; Легг, Дэвид А. (2010). «Изолированный pterygotid ramus (Chelicerata: Eurypterida) из девонской формации Beartooth Butte, штат Вайоминг». Журнал палеонтологии. 84 (6): 1206–1208. Дои:10.1666/10-040.1. S2CID 129807060.
- ^ Gould, Gina C .; Макфадден, Брюс Дж. (2004). Глава 17: Гигантизм, карликовость и правило Копа: «Ничто в эволюции не имеет смысла без филогении»"". Бюллетень Американского музея естественной истории. 285: 219–237. Дои:10.1206 / 0003-0090 (2004) 285 <0219: c> 2.0.co; 2.
- ^ а б c Маккой, Виктория Э .; Ламсделл, Джеймс С.; Пошманн, Маркус; Андерсон, Росс П .; Бриггс, Дерек Э. Г. (2015). «Приятно видеть вас: глаза и когти показывают эволюцию различных экологических ролей гигантских птеригидных эвриптерид». Письма о биологии. 11 (8): 20150564. Дои:10.1098 / rsbl.2015.0564. ЧВК 4571687. PMID 26289442.
- ^ Хлупач, Иво (1994). "Эвриптериды Pterygotid (Arthropoda, Chelicerata) в силурии и девоне Богемии" (PDF). Журнал Чешского геологического общества. 39/2-3: 147–162.
- ^ «Окаменелости: эвриптериды Мельбурнской группы, формация Дарджайл в Мельбурне (силурийский период Австралии)». fossilworks.org. Получено 2018-02-27.
- ^ «Формация Даргайл, участок водохранилища Виннеке (силурийский период Австралии)». fossilworks.org. Получено 2018-07-27.
- ^ а б Р. Э. Плотник. 1999. Среда обитания лландоверско-лохковских эвриптерид. В A. J. Boucot, J. D. Lawson (ред.), Палеосообщества - на примере силурия и нижнего девона
- ^ «Связанная с эвриптеридами биота горизонта корнецикула, Сааремаа, Эстония: Корнецикула, Эстония». paleobiodb.org. Получено 2018-07-27.
- ^ "Связанная с эвриптеридами биота сланцевого пласта Питтсфорд, Питтсфорд, Нью-Йорк: Ладлоу, Нью-Йорк". paleobiodb.org. Получено 2018-07-27.
- ^ «Пластинчатые известняково-сланцевые фации, верхний придолий, граничный разрез юг барранда: Придоли, Чехия». paleobiodb.org. Получено 2018-07-27.
- ^ "Сообщество Eurypterus remipes tetragonopthalmus, Устье подсвита, Подолье (силурий Украины)". paleobiodb.org. Получено 2018-07-27.
- ^ «Связанная с эвриптеридами биота чортковского горизонта Подолья (девон Украины)». paleobiodb.org. Получено 2018-07-27.
- ^ "Эвриптериды из Keefer Sst. Mbr, Mifflintown Fmn., Хэнкок, Мэриленд: Венлок, Мэриленд". paleobiodb.org. Получено 2018-07-27.
- ^ «Связанная с эвриптеридами биота формации Ледбери, Эвайас-Гарольд, Англия: Придоли, Соединенное Королевство». paleobiodb.org. Получено 2018-07-27.
- ^ «Скважина Sun Oil Co., керн 44 (3552-3568 футов): Лохковиан, Флорида». paleobiodb.org. Получено 2018-07-27.
- ^ "Сообщество Гаспе, песчаник Гаспе (девон Канады)". paleobiodb.org. Получено 2018-07-27.
внешние ссылки
 СМИ, связанные с Птеригот в Wikimedia Commons
СМИ, связанные с Птеригот в Wikimedia Commons
| ||
| Таксономия |  | |
| Геохронология |
| |
| География |
| |
| Известные роды | ||
| Ичногенера | ||
| Связанные группы | ||
| Статьи по Теме | ||