Криноид - Crinoid
| Морские лилии | |
|---|---|
 | |
| Морские лилии на рифе острова Бату Мончо, Индонезия | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Иглокожие |
| Подтип: | Crinozoa |
| Класс: | Crinoidea Миллер, 1821[2] |
| Подклассы | |
Articulata (540 видов) | |
Морские лилии морские животные, составляющие класс Crinoidea, один из классов филума Иглокожие, который также включает морская звезда, хрупкие звезды, морские ежи и морские огурцы.[3] Те морские лилии, которые во взрослой форме прикрепляются к морскому дну стеблем, обычно называются морские лилии, а незаметные формы называются перо звезды или коматулиды, являясь членами крупнейших морских лилий порядок, Коматулида.
Для взрослых морских лилий характерно расположение рта на верхней поверхности. Он окружен кормящими руками и соединен с U-образной кишкой, при этом анус расположен на ротовом диске около рта. Хотя основной паттерн иглокожих с пятичленной симметрией можно распознать, у большинства морских лилий пять рук подразделяются на десять и более. Они имеют перистые пиннулы и широко расставлены, чтобы собирать планктонный частицы из воды. На каком-то этапе жизни у большинства морских лилий есть стебель, который прикрепляется к субстрат, но многие живут прикрепленными только в подростковом возрасте и начинают свободно плавать во взрослом возрасте.
Насчитывается всего около 600 живых видов морских лилий,[4] но в прошлом этот класс был гораздо более многочисленным и разнообразным. Некоторые толстые известняк кровати, датируемые средним и поздним срокомПалеозой эпохи почти полностью состоят из разрозненных фрагментов криноидей.
Этимология
Название «Crinoidea» происходит от древнегреческого слова κρίνον (Кринон), "лилия", с суффиксом –Oid что означает «нравится».[5][6] Они живут как на мелководье[7] и на глубине до 9 000 метров (30 000 футов).[8] Те морские лилии, которые во взрослом виде прикрепляются к морскому дну стеблем, обычно называют морскими лилиями.[9] Незаметные формы называются перо звезды[10] или коматулиды, являющиеся членами крупнейших морских лилий порядок, Коматулида.[11]
Морфология
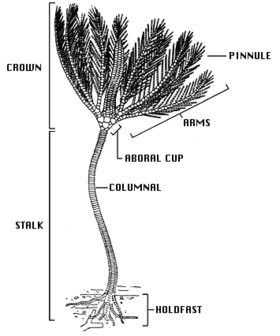
Основная форма тела лилий - это стебель (отсутствует у взрослых перьевых звезд) и корона, состоящая из чашеобразного центрального тела, известного как тека, и набора из пяти лучей или плеч, обычно разветвленных и перистых. В рот и анус оба расположены на верхней стороне теки, поэтому спинной (верхняя) поверхность ротовой поверхности, в отличие от других групп иглокожих, таких как морские ежи, морская звезда и хрупкие звезды где рот находится на нижней стороне.[12] Многочисленные известковые пластинки составляют основную часть криноидей с небольшой долей мягких тканей. Эти косточки хорошо окаменелости, и есть пласты известняка, датируемые Нижний карбон около Clitheroe, Англия, образовалась почти исключительно из разнообразной фауны окаменелостей лилий.[13]

Стебель морских лилий состоит из колонны высокопористых косточек, соединенных связочной тканью. Крепится к основанию плоским стойко держаться или с мутовками из сочлененных корневидных структур, известных как Cirri. Дальнейшие усики могут появиться выше по стеблю. У морских лилий, прикрепляющихся к твердым поверхностям, усики могут быть крепкими и изогнутыми, напоминая птичьи лапы, но когда криноидеи живут на мягких осадках, усики могут быть тонкими и стержневидными. У молодых перьевых звездочек есть стебель, но он позже утрачивается, и многие виды сохраняют несколько усиков у основания кроны. Большинство современных морских лилий плавают свободно и имеют только рудиментарный стебель. У тех глубоководных видов, которые все еще сохраняют стебель, он может достигать 1 м (3 фута) в длину (хотя обычно намного меньше), а известны ископаемые виды со стеблем длиной 20 м (66 футов).[14]
Theca - это пятичленный (имеет пятичастную симметрию) и является гомологичный с телом или диском других иглокожих. Основание теки образовано чашеобразным набором косточек (костных пластинок), чашечка, а верхняя поверхность образована слабо кальцинированными тегмен, перепончатый диск. Тегмен разделен на пять «амбулакральных зон», включая глубокую бороздку, из которой трубка ноги проект, и пять «межмбулакральных зон» между ними. Рот находится около центра или на краю тегмена, и амбулакральный бороздки ведут от основания рук ко рту. В анус также находится на тегмене, часто на небольшом приподнятом конусе, в межмбулакральный площадь. Тека относительно невелика и содержит органы пищеварения лилий.[14]
Руки поддерживаются серией сочленяющихся косточек, похожих на те, что на стебле. Первоначально у морских лилий было только пять рук, но в большинстве современных форм они разделены на две части у косточки II, что дает всего десять рук. У большинства живых видов, особенно у свободно плавающих перьевых звезд, руки разветвляются еще несколько раз, давая в общей сложности до двухсот ветвей. Будучи сочлененными, руки могут сгибаться. Поочередно с обеих сторон они выстланы более мелкими сочлененными отростками, известными как «пиннулы», которые придают им вид перьев. Обе руки и пиннулы имеют трубка ноги по краям амбулакральных бороздок. Трубчатые ножки бывают группами по три разного размера; у них нет присосок, они используются для удержания и манипулирования частицами пищи. Пазы снабжены реснички которые облегчают кормление, перемещая органические частицы вдоль руки и в рот.[14]

Стебель, тека и руки «настоящего» (стебельчатого) криноида (семейства Isselicrinidae )

Оксикомантус Беннетти (коматулид)

Тегмен а Лампрометра пальма. Рот находится в центре 5 питающих канавок, а задний проход - в верхней части колонны.

Крупный план усиков, которые позволяют коматулидам ходить и прикрепляться

Крупный план пиннулы Tropiometra carinata (с паразитами Мизостома фускомакулатум )





Биология
Кормление

Криноиды пассивны подвесные питатели, фильтрация планктон и мелкие частицы детрит из морской воды, протекающей мимо них их перьями руками. Плечи поднимаются, образуя веерообразную форму, которая удерживается перпендикулярно току. Мобильные морские лилии перемещаются, чтобы высесться на камнях, коралловых головах или других возвышенностях, чтобы максимально использовать возможности кормления. Частицы пищи улавливаются первичными (самыми длинными) ножками трубки, которые полностью вытянуты и удерживаются в вертикальном положении от пиннул, образуя сетку, улавливающую пищу, в то время как вторичные и третичные ножки трубки участвуют в манипулировании всем, что встречается.[14]
Ножки трубки покрыты липкой слизь это улавливает любые соприкасающиеся частицы. Как только они поймают частицу пищи, ножки трубки бросают ее в амбулакральный бороздка, по которой реснички продвигают слизь и частицы пищи к рту. Накладки сбоку от канавки помогают удерживать поток слизи на месте. Общая длина поверхности для улавливания пищевых продуктов может быть очень большой; 56 рук Японская морская лилия с кронштейнами 24 см (9 дюймов) имеют общую длину 80 м (260 футов), включая пиннулы. Вообще говоря, морские лилии, живущие в среде с относительно небольшим количеством планктона, имеют более длинные и сильно разветвленные руки, чем у морских лилий, живущих в среде, богатой пищей.[14]
Рот опускается в короткий пищевод. Настоящего желудка нет, поэтому пищевод соединяется напрямую с кишечник, который проходит в виде единой петли прямо вокруг внутренней части чашечки. Кишечник часто включает многочисленные дивертикулы, некоторые из которых могут быть длинными или разветвленными. Конец кишечника открывается короткой мышечной прямая кишка. Это восходит к анус, который выступает из небольшого конического выступа на краю тегмена. Фекалии образуются в виде больших гранул со слизистой оболочкой, которые падают на покров, а оттуда на субстрат.[14]
Хищничество
Экземпляры морского ежа Calocidaris micans найдено в районе лилий Endoxocrinus parrae, было показано, что они содержат большое количество стеблей в кишечнике. Они состоят из сочлененных косточек с мягкими тканями, тогда как местный осадок содержит только разъединенные косточки без мягких тканей. Это делает весьма вероятным, что эти морские ежи хищники криноидей, и что криноидеи убегают, предлагая при этом часть своего стебля.[15]
Различные окаменелости криноидей намекают на возможных доисторических хищников. Копролиты как рыбы, так и головоногие моллюски были обнаружены косточки различных морских лилий, таких как пелагический криноид Саккокома, от Юрский Lagerstatten Solnhofen,[16] а поврежденные стебли лилий со следами укусов, совпадающими с зубными пластинами кокостеид плакодермы были найдены в конце Девонский Польша.[17] Чашечки нескольких девонских до Каменноугольный морские лилии имеют раковину улитки, Platyceras, тесно связанные с ними.[18] У некоторых улитка расположена над анусом, что позволяет предположить, что Platyceras был копрофаг комменсал, в то время как у других животное расположено прямо над скважиной, что предполагает более опасные отношения.[19]
Водяная сосудистая система
Как и другие иглокожие, морские лилии обладают водная сосудистая система что поддерживает гидравлический давление в ножках трубки. Это не связано с внешней морской водой через мадрепорит, как и у других иглокожих, но только через большое количество пор соединенных с целомудрие (полость тела). Основным резервуаром жидкости является кольцевой канал с мускулистыми стенками, который соединен с целомом каменными каналами, выстланными известняком. Целом разделен на ряд взаимосвязанных пространств посредством брыжейки. Он окружает внутренние органы диска и имеет ветви внутри стебля и рук, с меньшими ветвями, заходящими в пиннулы. Это сокращение кольцевого канала, которое расширяет ножки трубки. Три узких ветви целома входят в каждое плечо, две на оральной стороне и одна аборально, и в пиннулы. Действие ресничек вызывает медленный поток жидкости (1 мм в секунду) в этих каналах, наружу в оральных ветвях и внутрь в аборальных, и это основное средство транспортировки питательных веществ и продуктов жизнедеятельности. Нет сердца и отдельной системы кровообращения, но у основания диска есть большой кровеносный сосуд, известный как осевой орган, содержащий несколько тонких трубок с слепыми концами неизвестной функции, которые простираются в стебель.[14]
Эти различные заполненные жидкостью пространства, помимо транспортировки питательных веществ по телу, также функционируют как дыхательная, так и выделительная система. Кислород поглощается в основном через ножки трубки, которые являются наиболее тонкостенными частями тела, а дальнейший газообмен происходит на большой площади рук. Нет специализированных органов для выделения, а отходы собираются фагоцитарный целомоциты.[14]
Нервная система
Нервная система лилий делится на три части, между которыми имеется множество связей. Оральная или самая верхняя часть - единственная гомологичный с нервной системой других иглокожих. Он состоит из центрального нервного кольца, окружающего рот, и лучевых нервов, ответвляющихся в руки, и выполняет сенсорную функцию. Ниже находится промежуточное нервное кольцо, от которого отходят лучевые нервы, снабжающие руки и пиннулы. Эти нервы имеют двигательную природу и контролируют мускулатуру трубчатых ножек. Третья часть нервной системы расположена в оральной части и отвечает за сгибание и движение рук, пиннул и усиков. Он сосредоточен на массе нервной ткани у основания чашечки и обеспечивает по одному нерву для каждой руки и несколько нервов для ножки.[14]
Размножение и жизненный цикл
Криноидеи не способны к клональному размножению, как некоторые морская звезда и хрупкие звезды, но способны восстанавливать утраченные части тела. Руки, оторванные хищниками или поврежденные неблагоприятными условиями окружающей среды, могут вырасти заново, и даже висцеральная масса может восстановиться в течение нескольких недель. Эта регенерация может быть жизненно важной для выживания после нападения хищных рыб.[14]
Морские лилии раздельнополый, причем люди могут быть мужчинами или женщинами. У большинства видов гонады расположены в пиннулах, но в некоторых случаях они расположены в руках. Не все пиннулы репродуктивные, только те, которые находятся ближе всего к макушке. В гаметы продуцируются в половых каналах, заключенных в генитальные целомы. Пиннулы в конечном итоге разрываются, чтобы освободить сперма и яйца в окружающую морскую воду. У некоторых родов, таких как Antedon оплодотворенные яйца прикрепляются к рукам секретами эпидермальных желез; у других, особенно холодноводных видов из Антарктиды, яйца задумчивый в специализированных мешочках на руках или пиннулах.[14]
Оплодотворенные яйца вылупляются, чтобы выпустить в свободное плавание. личинки желточников. Двусторонне симметричная личинка имеет бочкообразную форму с кольцами реснички бегает по всему телу, а пучок сенсорных волосков на верхнем полюсе. Хотя среди четырех других существующих классов иглокожих существуют как питающиеся (планктотрофные), так и не питающиеся (лецитотрофные) личинки, все современные морские лилии, по-видимому, являются потомками выжившей клады, прошедшей через горлышко бутылки после Пермское вымирание, в это время теряя стадию питающейся личинки.[20] Период свободного плавания личинки длится всего несколько дней, прежде чем она осядет на дно и прикрепится к подстилающей поверхности с помощью липкой железы на ее нижней стороне. Затем личинка претерпевает длительный период метаморфозы в преследуемый малолетний, становясь при этом радиально симметричным. Даже свободно плавающие перьевые звезды проходят через эту стадию, и взрослые особи в конечном итоге отрываются от стебля.[14]
Передвижение

Большинство современных лилий, то есть перьевых звезд, подвижны и не имеют стебля во взрослом возрасте. Примеры ископаемых морских лилий, которые были интерпретированы как свободно плавающие, включают Марсупица, Саккокома и Uintacrinus.[21] Обычно криноидеи перемещаются на новые места ползанием, используя усиков в качестве ног. Такое движение может быть вызвано изменением направления течения, необходимостью забраться на возвышение для кормления или агонистическим поведением встретившегося человека.[22] Морские лилии тоже умеют плавать. Они делают это путем скоординированных повторяющихся последовательных движений рук в трех группах. Сначала движение идет вверх, но вскоре становится горизонтальным, со скоростью около 7 см (2,8 дюйма) в секунду, при этом поверхность рта находится впереди. Плавание обычно происходит в виде коротких всплесков активности продолжительностью до получаса, а у коматулида. Florometra serratissima по крайней мере, происходит только после механической стимуляции или как реакция бегства, вызванная хищником.[22]
В 2005 году был зафиксирован стебельный криноид, тянувшийся по морскому дну от берега. Остров Гранд Багама. Хотя было известно, что стебельчатые морские лилии могут двигаться, до этой записи самое быстрое движение, известное для стебельчатых криноидей, составляло 0,6 метра (2 фута) в час. Запись 2005 года показала, что один из них движется по морскому дну с гораздо большей скоростью - от 4 до 5 см (1,6–2,0 дюйма) в секунду, или от 144 до 180 м (472–591 фут) в час.[23]
Эволюция
Происхождение


Если игнорировать загадочное Эхматокрин из Burgess Shale, самые ранние известные группы криноидей восходят к Ордовик, 480 миллионов лет назад. Существуют две конкурирующие гипотезы относительно происхождения группы: традиционная точка зрения утверждает, что морские лилии произошли изнутри бластозойные (в эокриноиды и их потомков, бластоиды и цистоиды ), тогда как наиболее популярная альтернатива предполагает, что морские лилии рано отделяются от эдриоастероиды.[24] Дискуссию трудно урегулировать, отчасти потому, что все три предка-кандидата обладают многими общими характеристиками, включая радиальную симметрию, известковые пластинки и стебельчатое или прямое прикрепление к субстрату.[24]
Разнообразие
Иглокожие с минерализованным скелетом вошли в летопись окаменелостей в раннем Кембрийский (540 млн лет назад), и в течение следующих 100 миллионов лет криноидеи и бластоиды (также стебельчатые фильтраторы) были доминирующими.[25] В то время иглокожие включали двадцать таксонов класс ранг, только пять из которых пережили последовавшие за этим массовые вымирания. Долгая и разнообразная геологическая история морских лилий показывает, насколько хорошо иглокожие приспособились к питанию через фильтр.[3]
Криноидеи пережили два периода резкого адаптивное излучение первый - в ордовике (485–444 млн лет назад), а второй - в раннем триасе (около 230 млн лет назад).[26] Это излучение триаса привело к широкому распространению форм, обладающих гибкими рукавами; подвижность, преимущественно в ответ на давление хищников, также стало гораздо более распространенным, чем сидячие.[27] Это излучение произошло несколько раньше, чем Мезозойская морская революция, возможно, потому, что это было в основном вызвано увеличением бентосных хищников, особенно ехиноидов.[28] Затем последовал выборочный массовое вымирание в конце Пермский период период, в течение которого вымерли все бластоиды и большинство лилий.[26] После вымирания в конце пермского периода криноидеи так и не восстановили морфологическое разнообразие и доминирующее положение, которыми они занимались в палеозое; они использовали набор открытых для них экологических стратегий, отличный от тех, которые оказались столь успешными в палеозое.[26]
Окаменелости
Некоторые ископаемые морские лилии, такие как Пентакриниты, похоже, жили прикрепленными к плавающим корягу, и часто встречаются целые колонии. Иногда коряги заболачивались и опускались на дно, унося с собой прикрепленных криноидей. Стебель Пентакриниты может быть длиной несколько метров. Современные родственники Пентакриниты живут в слабых потоках, прикрепленных к камням концом своего стебля. Самый крупный ископаемый криноид из всех зарегистрированных имел стебель длиной 40 м (130 футов).[29]
В 2012 году трое геологов сообщили, что изолировали сложные органические молекулы от животных возрастом 340 миллионов лет (Миссисипец ) окаменелости нескольких видов морских лилий. Определен как "похожий [ing ...] ароматный или полиароматический хиноны «, это самые старые молекулы, которые окончательно связаны с конкретными отдельными окаменелостями, так как считается, что они были запечатаны внутри пор косточек осажденным кальцитом во время процесса окаменения.[30]
Окаменелости лилий, и в особенности разрозненные столбчатые лилии, могут быть настолько многочисленными, что иногда служат основными поддерживающими обломками в осадочных породах.[нужна цитата ] Скалы такой природы называются энкринитирует.
Таксономия


Crinoidea была признана отдельной клады иглокожих с момента определения группы Миллером в 1821 году.[31] Согласно Всемирный регистр морских видов, Articulata, единственный существующий подкласс Crinoidea, включает следующие семейства: -[32]
- порядок Коматулида Кларк, 1908 год.
- суперсемья Antedonoidea Норман, 1865 г.
- семья Antedonidae Норман, 1865 г.
- семья Pentametrocrinidae А. Х. Кларк, 1908 г.
- семья Зенометрид А. Х. Кларк, 1909 г.
- суперсемья Ателекриноидея Купальщица, 1899 г.
- семья Ателекриниды Купальщица, 1899 г.
- суперсемья Comatuloidea Флеминг, 1828 г.
- семья Коматулиды Флеминг, 1828 г.
- суперсемья Himerometroida А. Х. Кларк, 1908 г.
- семья Colobometridae А. Х. Кларк, 1909 г.
- семья Eudiocrinidae А. Х. Кларк, 1907 г.
- семья Himerometridae А. Х. Кларк, 1907 г.
- семья Мариометриды А. Х. Кларк, 1909 г.
- семья Zygometridae А. Х. Кларк, 1908 г.
- суперсемья Notocrinoidea Мортенсен, 1918 г.
- семья Aporometridae Х. Л. Кларк, 1938 г.
- семья Notocrinidae Мортенсен, 1918 г.
- суперсемья Паракоматулоидеи Гесс, 1951 год. †
- суперсемья Тропиометроид А. Х. Кларк, 1908 г.
- семья Asterometridae Жислен, 1924 г.
- семья Calometridae А. Х. Кларк, 1911 г.
- семья Charitometridae А. Х. Кларк, 1909 г.
- семья Ptilometridae А. Х. Кларк, 1914 г.
- семья Thalassometridae А. Х. Кларк, 1908 г.
- семья Tropiometridae А. Х. Кларк, 1908 г.
- Коматулида incertae sedis
- семья Атопокриниды Мессинг, 2011 (в Hess & Messing, 2011)
- семья Батикриниды Купальщица, 1899 г.
- семья Bourgueticrinidae Лориоль, 1882 г.
- семья Guillecrinidae Миронов и Сорокина, 1998 г.
- семья Phrynocrinidae А. Х. Кларк, 1907 г.
- семья Септокриниды Миронов, 2000 г.
- суперсемья Antedonoidea Норман, 1865 г.
- порядок Циртокринида
- Подотряд Циртокринина
- семья Sclerocrinidae Якель, 1918 г.
- Подотряд Холоподина
- семья Eudesicrinidae Купальщица, 1899 г.
- семья Holopodidae Циттель, 1879 г.
- Подотряд Циртокринина
- порядок Encrinida †
- порядок Hyocrinida
- семья Hyocrinidae Плотник, 1884 г.
- порядок Isocrinida
- Подотряд Изокринина
- семья Cainocrinidae Симмс, 1988
- семья Isocrinidae Жислен, 1924 г.
- семья Isselicrinidae Кликушкин, 1977 г.
- семья Proisocrinidae Расмуссен, 1978
- Подотряд Пентакринитина †
- семья Pentacrinitidae Серый, 1842 г. †
- Подотряд Изокринина
- порядок Миллерикринида †
Филогения
В филогения, геологическая история и классификация Crinoidea был обсужден Райтом и др. (2017).[33] Эти авторы представили новые классификации на основе филогении и рангов, основанные на результатах недавних филогенетических анализов.[31][34][35][36] Их ранговая классификация высших таксонов лилий (вплоть до порядка), не решена полностью и с многочисленными группами incertae sedis (неопределенного размещения), показано на кладограмма.
| Crinoidea | |





В культуре
Ископаемые сегменты столбчатых лилий, извлеченные из известняк добыт на Линдисфарн, или были найдены выброшенными на берег, были врезаны в украшения на шею или розарии, и стал известен как Бусы Святого Катберта в Средний возраст.[37] Точно так же на Среднем Западе Соединенных Штатов окаменелые сегменты колонн морских лилий иногда называют Индийские бусы.[38] Криноиды - это государственное ископаемое из Миссури.[39]
Ископаемые морские лилии

Ископаемое из Германии, демонстрирующее стебель, чашечку и руки с пиннулами.

Окаменелости лилий возрастом 330 миллионов лет из Айова

Seirocrinus subangularis с раннего Юрский Посидония Сланец в Holzmaden, Германия

Столбцы лилий (Isocrinus nicoleti ) из Среднего Юрский Формация Кармель на перекрестке Маунт-Кармель, Юта

Корневидный криноид из верхнего ордовика, южный Огайо

Внутренний слепок просвета стебля лилий (и внешний слепок) из нижнего карбона, штат Огайо

Окаменелости Seirocrinus subsingularis из юрской формации Holzmaden Black Shale, Германия







использованная литература
- ^ Замора, Самуил; Рахман, Имран А .; Осич, Уильям I. (2015). «Палеогеографические последствия появления нового криноида иокринида (Disparida) из ордовика (дарривиллиан) в Марокко». PeerJ. 3: e1450. Дои:10.7717 / peerj.1450. ЧВК 4675106. PMID 26664800.
- ^ Ханссон, Ханс (2012). "Crinoidea". Черви. Всемирный регистр морских видов. Получено 2013-01-30.
- ^ а б Рупперт, Эдвард Э .; Фокс, Ричард, С .; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание. Cengage Learning. С. 917–918. ISBN 978-81-315-0104-7.
- ^ "Сеть разнообразия животных: Crinoidea". Музей зоологии Мичиганского университета. Получено 26 августа 2012.
- ^ Новый универсальный полный словарь Вебстера. 2-е изд. 1979 г.
- ^ "криноид". Интернет-словарь этимологии.
- ^ Змарзлы, Д. (1985). «Фауна мелководных морских лилий атолла Кваджалейн, Маршалловы острова: экологические наблюдения, межколловые сравнения и зоогеографические сходства». Тихоокеанская наука. 39: 340–358.
- ^ Oji, T .; Ogawa, Y .; Хантер, А. В. и Китадзава, К. (2009). «Обнаружение плотных скоплений морских лилий в желобе Идзу-Огасавара, Япония». Зоологическая наука. 26 (6): 406–408. Дои:10.2108 / zsj.26.406. PMID 19583499. S2CID 5991969.
- ^ "Морская лилия". Британская энциклопедия. Получено 14 марта 2011.
- ^ "Перо звезды". Британская энциклопедия. Получено 14 марта 2011.
- ^ Ausich, Вильгельм I; Мессинг, Чарльз Г. "Crinoidea". Древо жизни. Получено 14 марта 2011.
- ^ О'Хара, Тимоти; Бирн, Мария (2017). Австралийские иглокожие: биология, экология и эволюция. Csiro Publishing. С. 171–180. ISBN 978-1-4863-0763-0.
- ^ Гесс, Ганс; Brett, Carlton E .; Ausich, Вильгельм I; Симмс, Майкл Дж. (2002). Ископаемые морские лилии. Издательство Кембриджского университета. С. 3–5, 45–46. ISBN 978-0-521-52440-7.
- ^ а б c d е ж г час я j k л Рупперт, Эдвард Э .; Фокс, Ричард, С .; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание. Cengage Learning. С. 917–927. ISBN 978-81-315-0104-7.
- ^ Baumiller, Tomasz K .; Муи, Рич; Мессинг, Чарльз Г. (2008). «Ежи на лугу: палеобиологические и эволюционные последствия хищничества цидароидных лилий». Палеобиология. 34 (1): 22–34. Дои:10.1666/07031.1. JSTOR 20445573. S2CID 85647638.
- ^ Гесс, Ганс (2003). «Верхняя юра Зольнхофен Платтенкальк в Баварии, Германия». В Brett, Carlton E .; Ausich, Вильгельм I; Симмс, Майкл Дж. (Ред.). Ископаемые морские лилии. Издательство Кембриджского университета. С. 216–24. ISBN 978-0-521-52440-7.
- ^ Горжелак, Закон Пшемиса; Ракович, Лукаш; Саламон, Мариуш А .; Шрек, Петр (2011). «Предполагаемые следы укусов плакодермы на девонских лилиях из Польши». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 259: 105–12. Дои:10.1127/0077-7749/2010/0111.
- ^ Brett, Carlton E .; Уокер, Салли Э. (2002). «Хищники и хищники в палеозойской морской среде» (PDF). Документы Палеонтологического общества. 8: 93–118. Дои:10.1017 / S1089332600001078. Архивировано из оригинал (PDF) на 2012-08-13. Получено 2014-04-06.
- ^ Gahn, Forest J .; Баумиллер, Томаш К. (2003). «Заражение среднедевонских (живетских) камерных лилий платицератидными брюхоногими моллюсками и его последствия для характера их биотического взаимодействия» (PDF). Lethaia. 36 (2): 71–82. Дои:10.1080/00241160310003072. HDL:2027.42/75509.
- ^ Рафф, Р. А; Бирн, М. (2006). «Активная эволюционная жизнь личинок иглокожих». Наследственность. 97 (3): 244–52. Дои:10.1038 / sj.hdy.6800866. PMID 16850040.
- ^ "О криноидах". FossilEra. Получено 15 марта 2019.
- ^ а б Shaw, G.D .; Фонтейн, А. (2011). "Передвижение коматулида Florometra serratissima (Иглокожие: Crinoidea) и его адаптивное значение ». Канадский журнал зоологии. 68 (5): 942–950. Дои:10.1139 / z90-135.
- ^ Baumiller, Tomasz K .; Мессинг, Чарльз Г. (6 октября 2005 г.). «Ползание в стебельчатых морских лилиях: наблюдения на месте, функциональная морфология и последствия для палеозойских таксонов». Рефераты Геологического общества Америки с программами. 37. п. 62. Архивировано с оригинал 7 апреля 2014 г.. Получено 6 апреля 2014.
- ^ а б Guensburg, Thomas E .; Муи, Рич; Посыпать, Джеймс; Дэвид, Бруно; Лефевр, Бертран (2010). «Руки пельматозоидов из среднего кембрия Австралии: Преодоление разрыва между брахиолами и плечевыми костями? Комментарий: Нет моста». Lethaia. 43 (3): 432–40. Дои:10.1111 / j.1502-3931.2010.00220.x.
- ^ Ваггонер, Бен (16 января 1995). «Иглокожие: летопись окаменелостей». Введение в иглокожих. Музей палеонтологии: Калифорнийский университет в Беркли. Получено 30 марта 2019.
- ^ а б c Фут, Майк (1999). «Морфологическое разнообразие эволюционной радиации палеозойских и постпалеозойских лилий». Палеобиология. 25 (sp1): 1–116. Дои:10.1666 / 0094-8373 (1999) 25 [1: MDITER] 2.0.CO; 2. ISSN 0094-8373. JSTOR 2666042.
- ^ Баумиллер, Томаш К. (2008). «Экологическая морфология лилий». Ежегодный обзор наук о Земле и планетах. 36: 221–249. Bibcode:2008AREPS..36..221B. Дои:10.1146 / annurev.earth.36.031207.124116.
- ^ Baumiller, T. K .; Саламон, М. А .; Gorzelak, P .; Mooi, R .; Messing, C.G .; Ган, Ф. Дж. (2010). «Постпалеозойская радиация криноидей в ответ на бентические хищники предшествовала мезозойской морской революции». Труды Национальной академии наук. 107 (13): 5893–6. Bibcode:2010PNAS..107.5893B. Дои:10.1073 / pnas.0914199107. JSTOR 25665085. ЧВК 2851891. PMID 20231453. ИНИСТ:22572914.
- ^ Понсонби, Дэвид; Дуссар, Джордж (2005). Анатомия моря. Ванкувер: Raincoast Books. п. 129. ISBN 978-0-8118-4633-2.
- ^ O'Malley, C.E .; Ausich, W. I .; Чин, Ю.-П. (2013). «Выделение и характеристика самых ранних таксон-специфичных органических молекул (Миссисипи, Crinoidea)». Геология. 41 (3): 347. Bibcode:2013Гео .... 41..347O. Дои:10.1130 / G33792.1. Сложить резюме – Phys.org (19 февраля 2013 г.). Обратите внимание, что первое предложение статьи Phys.org противоречит самой статье, в которой рассматривается несколько выделений молекул из определенных окаменелостей за последнее десятилетие.
- ^ а б Ausich, Вильгельм I; Каммер, Томас У .; Ренберг, Элизабет С .; Райт, Дэвид Ф. (2015). «Ранняя филогения лилий в кладе пельматозоидов». Палеонтология. 58 (6): 937–952. Дои:10.1111 / pala.12204.
- ^ Мессинг, Чарльз (2007). "Артикулата". Черви. Всемирный регистр морских видов. Получено 31 марта 2019.
- ^ Райт, Дэвид Ф .; Ausich, Вильгельм I; Коул, Селина Р .; Питер, Марк Э .; Ренберг, Элизабет С. (2017). «Филогенетическая таксономия и классификация Crinoidea (Echinodermata)». Журнал палеонтологии. в печати (4): 829–846. Дои:10.1017 / jpa.2016.142.
- ^ Райт, Дэвид Ф. (2017). «Байесовская оценка филогенеза окаменелостей и эволюции ранних и средних палеозойских лилий (иглокожих)». Журнал палеонтологии. 91 (4): 799–814. Дои:10.1017 / jpa.2016.141.
- ^ Коул, Селина Р. (2017). «Филогения и морфологическая эволюция ордовикских камер (класс Crinoidea, тип Echinodermata)». Журнал палеонтологии. 91 (4): 815–828. Дои:10.1017 / jpa.2016.137.
- ^ Роуз, Грег У .; Jermiin, Lars S .; Wilson, Nerida G .; Экхаут, Игорь; Лантербек, Дебора; Одзи, Тацуо; Янг, Крейг М .; Браунинг, Тина; Цистернас, Паула; Helgen, Lauren E .; Стаки, Мишель; Мессинг, Чарльз Г. (2013). «Фиксированный, свободный и фиксированный: непостоянная филогения современных Crinoidea (Echinodermata) и их пермско-триасовое происхождение». Молекулярная филогенетика и эволюция. 66 (6): 161–181. Дои:10.1016 / j.ympev.2012.09.018. PMID 23063883.
- ^ Лейн, Н. Гэри; Осич, Уильям I. (2001). «Легенда о четках святого Катберта: палеонтологическая и геологическая перспектива». Фольклор. 112 (1): 65–73. JSTOR 1260865.
- ^ «Идентификация неизвестных окаменелостей (по их форме)». Геологическая служба Кентукки / Университет Кентукки. Получено 21 июн 2009.
- ^ "Ископаемое в штате Миссури". Офис государственного секретаря, штат Миссури. Получено 31 марта 2019.
внешние ссылки
- Мессинг, Чарльз. "Морская звезда на палочке: знакомство с морскими лилиями". Vimeo.
 СМИ, связанные с Crinoidea в Wikimedia Commons
СМИ, связанные с Crinoidea в Wikimedia Commons Данные, относящиеся к Криноид в Wikispecies
Данные, относящиеся к Криноид в Wikispecies