Термометр РНК - RNA thermometer
An Термометр РНК (или же Термосенсор РНК) это температура -чувствительный некодирующая РНК молекула, которая регулирует экспрессия гена. Термометры с РНК часто регулируют гены, необходимые во время тепловой удар или же холодный шок ответ, но были вовлечены в другие регулирующие роли, такие как патогенность и голодание.[1]
Обычно термометры РНК работают, изменяя свои вторичная структура в ответ на колебания температуры. Этот структурный переход может затем обнажить или перекрыть важные области РНК, такие как сайт связывания рибосомы, что затем влияет на перевод скорость кодирования близлежащего белка ген.
РНК-термометры вместе с рибопереключатели, используются в качестве примеров в поддержку Гипотеза мира РНК. Эта теория предполагает, что когда-то РНК была единственной нуклеиновая кислота присутствует в ячейках, и был заменен текущим ДНК → РНК → белок система.[2]
Примеры термометров РНК включают: FourU,[3] то Hsp90 СНГ-регулирующий элемент,[4] то РОЗА элемент,[5] то Термометр Lig РНК[6], а Термометр hsp17.[7]
Открытие
О первом термочувствительном элементе РНК было сообщено в 1989 году.[8] До этого исследования мутации, предшествующие сайт начала транскрипции в лямбда (λ) фаг cIII мРНК было обнаружено, что они влияют на уровень трансляции белка cIII.[9] Этот белок участвует в селекции либо литический или же лизогенный жизненный цикл фага λ с высокими концентрациями cIII, способствующими лизогению.[9] Дальнейшее изучение этого участка РНК, расположенного выше по течению, выявило две альтернативные второстепенные конструкции; экспериментальное исследование показало, что конструкции взаимозаменяемы и зависят от обоих ион магния концентрация и температура.[8][10] В настоящее время считается, что этот термометр для РНК способствует вступлению в литический цикл при тепловом стрессе, чтобы бактериофаг для быстрой репликации и выхода из клетки-хозяина.[1]
Термин «термометр РНК» не вводился до 1999 г.[11] когда это было применено к rpoH Элемент РНК, идентифицированный в кишечная палочка.[12] В последнее время, биоинформатика поиск был использован для обнаружения нескольких новых термометров РНК-кандидата.[13] Однако традиционный поиск на основе последовательностей неэффективен, поскольку вторичная структура элемента намного больше консервированный чем последовательность нуклеиновой кислоты.[13]
Распределение
Наиболее известные термометры РНК расположены в 5 'нетранслируемая область (UTR) кодирования информационной РНК белки теплового шока - хотя предполагалось, что этот факт может быть частично связан с систематическая ошибка выборки и присущие трудности обнаружения коротких, неконсервативных последовательностей РНК в геномные данные.[14][15]
Хотя преимущественно встречается в прокариоты, потенциальный термометр РНК был обнаружен в млекопитающие включая люди.[16] РНК-1 термодатчика теплового шока (HSR1) активирует фактор транскрипции теплового шока 1 (HSF1) и индуцирует защитные белки, когда температура клетки превышает 37 ° C (температура тела ), предотвращая перегрев ячеек.[16]
Структура

РНК-термометры структурно просты и могут быть изготовлены из коротких последовательностей РНК; самый маленький - всего 44 нуклеотиды и находится в мРНК белка теплового шока, hsp17, в Synechocystis разновидность PCC 6803.[18][19] Обычно эти элементы РНК имеют длину от 60 до 110 нуклеотидов.[20] и они обычно содержат заколка для волос с небольшим количеством несовпадающих пар оснований, которые снижают стабильность структуры, тем самым облегчая разворачивание в ответ на повышение температуры.[21]
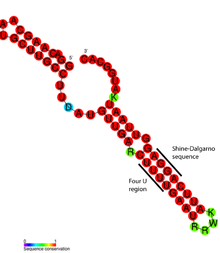
Детальный структурный анализ термометра ROSE RNA показал, что несовпадающие основания действительно участвуют в нестандартном спаривании оснований, которое сохраняет спиральную структуру РНК (см. Рисунок). Необычные пары оснований состоят из пар G-G, U-U и UC-U. Поскольку эти неканонические пары оснований относительно нестабильны, повышенная температура вызывает локальное плавление структуры РНК в этой области, обнажая последовательность Шайна-Далгарно.[17]
Некоторые РНК-термометры значительно сложнее одной шпильки, как в случае области, обнаруженной в МРНК CspA который, как считается, содержит псевдоузел, а также несколько шпилек.[22][23]
Синтетический Термометры РНК были разработаны с простой структурой с одной шпилькой.[24] Тем не менее вторичная структура таких коротких РНК-термометров могут быть чувствительны к мутации, как одиночный изменение базы может сделать шпильку неактивной in vivo.[25]
Механизм

Термометры РНК находятся в 5′ UTR матричной РНК перед геном, кодирующим белок.[1] Здесь они могут перекрывать сайт связывания рибосомы (RBS) и предотвращать трансляцию мРНК в белок.[14] При повышении температуры структура шпильки может «расплавиться» и обнажить RBS или Последовательность Шайна-Далгарно для связывания малой субъединицы рибосомы (30S ), который затем собирает другие машины для перевода.[1] В стартовый кодон, обычно обнаруживается на 8 нуклеотидов ниже последовательности Шайна-Далгарно,[14] сигнализирует о начале гена, кодирующего белок, который затем транслируется в пептид продукт от рибосома. В дополнение к этому СНГ-игра актеров механизм, единственный пример транс-игра актеров Термометр РНК был найден в МРНК RpoS где считается, что он участвует в голодании.[1]
Конкретным примером мотива РНК-термометра является термометр FourU, найденный в Salmonella enterica.[3] При воздействии температур выше 45 ° C стебель-петля который пар оснований напротив, последовательность Шайна-Дальгарно становится непарной и позволяет мРНК проникать в рибосому для осуществления трансляции.[25] Mg2+ Также было показано, что концентрация ионов влияет на стабильность FourU.[26] Самый хорошо изученный термометр РНК находится в rpoH ген в кишечная палочка.[27] Этот термодатчик активирует белки теплового шока при высоких температурах через σ32, специализированный тепловой шок фактор сигма.[11]
Хотя РНК-термометры обычно связаны с экспрессией белков, индуцированной нагреванием, они также могут регулировать белки холодового шока.[22] Например, выражение два 7кДа белки регулируются термометром РНК в теплолюбивый бактерия Термус термофильный[28] и аналогичный механизм был идентифицирован в Энтеробактерии.[23]
Термометры РНК, чувствительные к температуре 37 ° C, могут использоваться патогены для активации генов, специфичных для инфекции.[14] Например, усиление регулирования из прфА, кодирующий ключевой регулятор транскрипции вирулентность гены в Listeria monocytogenes, было продемонстрировано слияние 5 'ДНК прфА к зеленый флуоресцентный белок ген; затем слияние генов было транскрибировано с промотора Т7 в Кишечная палочка, а флуоресценция наблюдалась при 37 ° C, но не при 30 ° C.[29]
Последствия для гипотезы мира РНК
Гипотеза мира РНК утверждает, что когда-то РНК была как носителем наследственной информации, так и ферментативно активный, с различными последовательностями, действующими как биокатализаторы, регуляторы и сенсоры.[30] Затем гипотеза предполагает, что современные ДНК, РНК и жизнь на основе белков развился и отбор заменил большинство ролей РНК другими биомолекулы.[2]
РНК-термометры и рибопереключатели считаются эволюционно древний из-за их широкого распространения среди отдаленных организмов.[31] Было высказано предположение, что в мире РНК термодатчики РНК должны были отвечать за температурно-зависимую регуляцию других молекул РНК.[2][32] РНК-термометры в современных организмах могут быть молекулярные окаменелости что может указывать на ранее более широкое значение в мире РНК.[2]
Другие примеры
- Цис-регуляторный элемент Hsp90 регулирует hsp90 в Дрозофила, увеличивая скорость трансляции белка теплового шока при высоких температурах.[4]
- В ibpAB оперон из Кишечная палочка предполагается, что он будет содержать два кооперативных РНК-термометра: РОЗА элемент и IbpB термометр.[33]
- РОЗА1 и РОЗАAT2 находятся в ризобиальные Bradyrhizobium japonicum и Agrobacterium tumefaciens соответственно. Они существуют в 5 ′ UTR HspA мРНК и подавляют трансляцию белков теплового шока при физиологических температурах.[5][34]
- Цианобактериальные РНК-термометры
- Термометр межгенной РНК
- Термометры Neisseria RNA
- Термометр Lig РНК
Рекомендации
- ^ а б c d е ж Narberhaus F, Waldminghaus T, Chowdhury S (январь 2006 г.). «Термометры РНК». FEMS Microbiol. Rev. 30 (1): 3–16. Дои:10.1111 / j.1574-6976.2005.004.x. PMID 16438677.
- ^ а б c d Аткинс Дж. Ф., Гестеланд РФ, Чех Т. (2006). Мир РНК: природа современной РНК предполагает мир пребиотической РНК. Плейнвью, Нью-Йорк: Лаборатория Колд-Спринг-Харбор. ISBN 978-0-87969-739-6.
- ^ а б Waldminghaus T, Heidrich N, Brantl S, Narberhaus F (июль 2007 г.). "FourU: новый тип термометра РНК в Сальмонелла". Мол. Микробиол. 65 (2): 413–424. Дои:10.1111 / j.1365-2958.2007.05794.x. PMID 17630972.
- ^ а б Ахмед Р., Дункан РФ (2004). "Регуляция трансляции мРНК Hsp90. Элементы 5'-нетранслируемой области AUG, необходимые для предпочтительной трансляции теплового шока". J Biol Chem. 279 (48): 49919–49930. Дои:10.1074 / jbc.M404681200. PMID 15347681.
- ^ а б Нокер А., Хаушер Т., Балсигер С., Крстулович Н.П., Хеннеке Н., Нарберхаус Ф. (2001). «Термосенсор на основе мРНК контролирует экспрессию генов теплового шока ризобий». Нуклеиновые кислоты Res. 29 (23): 4800–4807. Дои:10.1093 / nar / 29.23.4800. ЧВК 96696. PMID 11726689.
- ^ Мацунага Дж., Шлакс П.Дж., Хааке Д.А. (2013-11-15). «Роль цис-действующих последовательностей РНК в температурно-зависимой экспрессии мультиадгезивных белков-лиг у Leptospira interrogans». Журнал бактериологии. 195 (22): 5092–5101. Дои:10.1128 / jb.00663-13. ISSN 0021-9193. ЧВК 3811586. PMID 24013626.
- ^ Kortmann J, Sczodrok S, Rinnenthal J, Schwalbe H, Narberhaus F (2011). «Перевод по запросу простым термодатчиком на основе РНК». Нуклеиновые кислоты Res. 39 (7): 2855–2868. Дои:10.1093 / nar / gkq1252. ЧВК 3074152. PMID 21131278.
- ^ а б Алтувиа С., Корницер Д., Тефф Д., Оппенгейм А.Б. (1989-11-20). «Альтернативные структуры мРНК гена cIII бактериофага лямбда определяют скорость инициации его трансляции». Журнал молекулярной биологии. 210 (2): 265–280. Дои:10.1016 / 0022-2836 (89) 90329-Х. PMID 2532257.
- ^ а б Altuvia S, Oppenheim AB (июль 1986 г.). «Трансляционные регуляторные сигналы в кодирующей области гена бактериофага лямбда cIII». Журнал бактериологии. 167 (1): 415–419. Дои:10.1128 / jb.167.1.415-419.1986. ЧВК 212897. PMID 2941413.
- ^ Алтувиа С., Корницер Д., Коби С., Оппенгейм А.Б. (1991-04-20). «Функциональные и структурные элементы мРНК гена cIII бактериофага лямбда». Журнал молекулярной биологии. 218 (4): 723–733. Дои:10.1016/0022-2836(91)90261-4. PMID 1827163.
- ^ а б Сторц Г. (1999-03-15). «Термометр РНК». Гены и развитие. 13 (6): 633–636. Дои:10.1101 / gad.13.6.633. PMID 10090718.
- ^ Морита М.Т., Танака Й., Кодама Т.С., Кёгоку Ю., Янаги Х., Юра Т. (1999-03-15). «Трансляционная индукция фактора транскрипции теплового шока sigma32: свидетельство встроенного термодатчика РНК». Гены и развитие. 13 (6): 655–665. Дои:10.1101 / gad.13.6.655. ЧВК 316556. PMID 10090722.
- ^ а б Waldminghaus T, Gaubig LC, Narberhaus F (ноябрь 2007 г.). «Полногеномное биоинформатическое предсказание и экспериментальная оценка потенциальных термометров РНК». Молекулярная генетика и геномика: MGG. 278 (5): 555–564. Дои:10.1007 / s00438-007-0272-7. PMID 17647020.
- ^ а б c d Нарберхаус F (2010). «Трансляционный контроль генов бактериального теплового шока и вирулентности с помощью чувствительных к температуре мРНК». РНК Биол. 7 (1): 84–89. Дои:10.4161 / rna.7.1.10501. PMID 20009504. Получено 2011-04-23.
- ^ Йоханссон Дж. (2009). Термосенсоры РНК в бактериальных патогенах. Contrib Microbiol. Вклад в микробиологию. 16. Базель. С. 150–160. Дои:10.1159/000219378. ISBN 978-3-8055-9132-4. PMID 19494584.
- ^ а б Шамовский И., Иванников М., Кандел Е.С., Гершон Д., Нудлер Э. (март 2006 г.). «РНК-опосредованный ответ на тепловой шок в клетках млекопитающих». Природа. 440 (7083): 556–560. Bibcode:2006Натура.440..556С. Дои:10.1038 / природа04518. PMID 16554823.
- ^ а б Чоудхури С., Марис С., Аллен Ф. Х., Нарберхаус Ф. (07.06.2006). «Молекулярная основа измерения температуры с помощью термометра РНК». Журнал EMBO. 25 (11): 2487–2497. Дои:10.1038 / sj.emboj.7601128. ЧВК 1478195. PMID 16710302.
- ^ Kortmann J, Sczodrok S, Rinnenthal J, Schwalbe H, Narberhaus F (апрель 2011 г.). «Перевод по запросу простым термодатчиком на основе РНК». Исследования нуклеиновых кислот. 39 (7): 2855–2868. Дои:10.1093 / nar / gkq1252. ЧВК 3074152. PMID 21131278.
- ^ Kortmann J, Sczodrok S, Rinnenthal J, Schwalbe H, Narberhaus F (апрель 2011 г.). «Перевод по запросу простым термодатчиком на основе РНК». Нуклеиновые кислоты Res. 39 (7): 2855–2868. Дои:10.1093 / nar / gkq1252. ЧВК 3074152. PMID 21131278. Получено 2011-04-23.
- ^ Waldminghaus T, Fippinger A, Alfsmann J, Narberhaus F (декабрь 2005 г.). «РНК-термометры распространены у альфа- и гамма-протеобактерий». Биол. Chem. 386 (12): 1279–1286. Дои:10.1515 / BC.2005.145. PMID 16336122.
- ^ Narberhaus F (январь – февраль 2010 г.). «Трансляционный контроль генов бактериального теплового шока и вирулентности с помощью чувствительных к температуре мРНК». РНК Биология. 7 (1): 84–89. Дои:10.4161 / rna.7.1.10501. PMID 20009504.
- ^ а б Breaker RR (январь 2010 г.). «РНК отключается на морозе». Мол. Клетка. 37 (1): 1–2. Дои:10.1016 / j.molcel.2009.12.032. ЧВК 5315359. PMID 20129048.
- ^ а б Джулиодори А.М., Ди Пьетро Ф., Марци С., Маскида Б., Вагнер Р., Ромби П., Гуалерци Колорадо, Пон CL (январь 2010 г.). «МРНК cspA представляет собой термодатчик, который модулирует трансляцию белка холодового шока CspA». Мол. Клетка. 37 (1): 21–33. Дои:10.1016 / j.molcel.2009.11.033. PMID 20129052.
- ^ Neupert J, Karcher D, Bock R (ноябрь 2008 г.). «Разработка простых термометров с синтетической РНК для контролируемой температуры экспрессии генов в Escherichia coli». Исследования нуклеиновых кислот. 36 (19): e124. Дои:10.1093 / nar / gkn545. ЧВК 2577334. PMID 18753148.
- ^ а б Николова Е.Н., Аль-Хашими Х.М. (сентябрь 2010 г.). «Термодинамика плавления РНК, по одной паре оснований за раз». РНК. 16 (9): 1687–1691. Дои:10.1261 / rna.2235010. ЧВК 2924531. PMID 20660079.
- ^ Ринненталь Дж., Клинкерт Б., Нарберхаус Ф, Швальбе Х (4 июля 2011 г.). "Модуляция стабильности Сальмонелла термометр РНК типа 4U ». Исследования нуклеиновых кислот. 39 (18): 8258–8270. Дои:10.1093 / nar / gkr314. ЧВК 3185406. PMID 21727085.
- ^ Шах П., Гилкрист МА (2010). Спирин А.С. (ред.). «Является ли уникальное свойство термочувствительности РНК-термометров?». PLOS ONE. 5 (7): e11308. Дои:10.1371 / journal.pone.0011308. ЧВК 2896394. PMID 20625392.
- ^ Мега Р., Манзоку М., Синкай А., Накагава Н., Курамицу С., Масуи Р. (август 2010 г.). "Очень быстрая индукция белка холодового шока путем понижения температуры в Термус термофильный". Biochem. Биофиз. Res. Сообщество. 399 (3): 336–340. Дои:10.1016 / j.bbrc.2010.07.065. PMID 20655297.
- ^ Йоханссон Дж., Мандин П., Рензони А., Кьяруттини С., Спрингер М., Коссарт П. (сентябрь 2002 г.). "Термосенсор РНК контролирует экспрессию генов вирулентности в Listeria monocytogenes". Клетка. 110 (5): 551–561. Дои:10.1016 / S0092-8674 (02) 00905-4. PMID 12230973.
- ^ Гилберт В (Февраль 1986 г.). «Мир РНК». Природа. 319 (6055): 618. Bibcode:1986Натура.319..618Г. Дои:10.1038 / 319618a0.
- ^ Серганов А., Патель DJ (октябрь 2007 г.). «Рибозимы, рибопереключатели и не только: регуляция экспрессии генов без белков». Природа Обзоры Генетика. 8 (10): 776–790. Дои:10.1038 / nrg2172. ЧВК 4689321. PMID 17846637.
- ^ Bocobza SE, Aharoni A (октябрь 2008 г.). «Включение света на растительных рибовключателях». Тенденции в растениеводстве. 13 (10): 526–533. Дои:10.1016 / j.tplants.2008.07.004. PMID 18778966.
- ^ Gaubig LC, Waldminghaus T, Narberhaus F (январь 2011 г.). «Множественные уровни контроля регулируют экспрессию оперона теплового шока Escherichia coli ibpAB». Микробиология. 157 (Pt 1): 66–76. Дои:10.1099 / мик.0.043802-0. PMID 20864473.
- ^ Балсигер С., Рагаз С., Барон С., Нарберхаус Ф. (2004). «Репликон-специфическая регуляция малых генов теплового шока в Agrobacterium tumefaciens». J Бактериол. 186 (20): 6824–6829. Дои:10.1128 / JB.186.20.6824-6829.2004. ЧВК 522190. PMID 15466035.