Проневральные гены - Proneural genes
Проневральные гены кодировать факторы транскрипции из основная спираль-петля-спираль (bHLH), которые отвечают за разработку нейроэктодермальный клетки-предшественники. Проневральные гены выполняют несколько функций в нейронное развитие. Они объединяют информацию о положении и вносят вклад в спецификацию идентичности клеток-предшественников.[1] Из того же эктодермальный типы клеток, нервные или эпидермальные клетки могут развиваться на основе взаимодействий между пронейральными и нейрогенными генами.[2] Нейрогенные гены называются так потому, что мутанты с потерей функции показывают увеличение количества развитых нейронных предшественников. С другой стороны, мутанты пронейральных генов не способны развить клетки-предшественники нейронов.[3]
Пронейральные гены экспрессируются в группах клеток (пронейральных кластерах), из которых будет выделена одна клетка-предшественник - обычно посередине, что приводит к образованию множества различных типов клеток. нейроны в центральный и периферическая нервная система.[4][5] Проневральные гены кодируют группу белков bHLH, которые играют решающую роль в контроле судьбы клеток в различных типах тканей.[6] Основные белки спираль-петля-спираль характеризуются двумя альфа-спирали разделены петлей. Спирали опосредуют димеризация, а прилегающая базовая область требуется для ДНК привязка.[1] В человеческий геном содержит примерно 125 факторов bHLH.[7]
Открытие
Впервые пронейральные гены были идентифицированы в 1920-х годах.[кем? ], когда мутант мухи, лишенные некоторых внешних органов чувств или щетина были найдены.[8] Позже, в 1970-х, комплекс ахэте-щиток, комплекс генов, которые участвуют в регуляции ранних стадий нервного развития у Дрозофила, были определены[кем? ].[9] С помощью молекулярных инструментов удалось выделить первые четыре гена этого комплекса: ачаэте (ас), щиток (sc), смертоносный щиток (lsc) и asense (асэ).[9] Другой пронейральный ген, атональный (ато)[10] был изолирован совсем недавно[когда? ] и два родственных ато гена, amos и cato, были позже выделены, определив второе семейство пронейральных генов - атональный комплекс.[11] Недавно[когда? ], первым гомологом пронейральных генов мух, обнаруженным у млекопитающих, был пюре1.[4]
Список пронейральных генов
Этот список относится к bHLH белки, обнаруженные в беспозвоночные и позвоночные. Они сгруппированы в разные семейства на основе более близкого сходства последовательностей в домене bHLH:[1]
| Организмы | Е-белки | атональная семья | семья нато | семья олиго | семья NeuroD | Нейрогенин семья | семья ахате-скутов | Семья нскл |
|---|---|---|---|---|---|---|---|---|
| беспозвоночные | Без дочери | Атонал, Амос, Катон, Лин32 | НАТО3 | Двурожие | CnASH, Ахете (ac), Asense (асэ), Щиток (sc), Смертельный щиток (lsc) | |||
| позвоночные | E-12 | Math1, Math5 | НАТО3 | Beta3, Beta4, Olig1, Olig2, Olig3 | NeuroD1 (NeuroD), NeuroD2 (Ndrf), NeuroD4 (Math3), NeuroD6 (Math2, Nex1) | амфиоксNgn, Ngn1, Ngn2, Ngn3 | Cash4, Маш1, Маш2, Хаш3 | Nscl1, Nscl2 |
Функции пронейральных генов
Гены ASC и Ngn семьи и, возможно, члены семейства гомологов ато, имеют сходную пронейральную функцию в позвоночные к их Дрозофила аналоги, тогда как другие нейронные bHLH гены участвуют в определении судьбы нейронов или в дифференцировке нейронов, но не играют пронейральной роли.[1]
Нейронные функции
Проневральные белки связываются ДНК как гетеродимерные комплексы, образованные белки bHLH или Е белки. Потому что гетеродимеризация является предпосылкой для ДНК обязательные, факторы, которые мешают димеризация эффективно действовать как пассив репрессоры активности пронейральных генов. Проневральные белки специфически связываются ДНК последовательности, содержащие основной гексануклеотидный мотив, CANNTG, известный как Электронная коробка. Основная область и спираль 1 bHLH домен образуют длинный альфа-спираль который связан с областью петли к спирали 2. Прямые контакты между bHLH остатки и ДНК отвечают за общую способность нейронных белки bHLH привязать к сердцевине E-boxsequence.[1] Клетки в кластере, которые экспрессируют пронейральный ген (называемый пронейральным кластером), можно рассматривать как клетки группы эквивалентности. В пронейральном кластере клетки конкурируют друг с другом, так что только часть клеток выделяется для развития в нейрональных предшественников. Этот процесс выделения опосредуется межклеточные взаимодействия интерпретируется через действие нейрогенных генов. В нейроэктодерма нейрогенные гены необходимы для выделения клеток внутри пронейральных кластеров для формирования нейрональных предшественников, оставляя оставшиеся клетки пронейральных кластеров для развития в эпидермальные клетки.[12] Проневральные гены могут функционировать аналогичным образом в позвоночные и беспозвоночные, в частности, они были причастны к раннему нейрогенез.[13] Хотя пронейральные белки ответственны за триггер нейрогенез, разные белки требуются для разных нервных и / или глиальная клетка типы. Это означает, что каждый из этих белков способен регулировать оба общих гена-мишени для нейрогенез и уникальные гены-мишени для характеристик подтипа нейронов.[14] Пронейральный bHLH факторы транскрипции не только диск нейрогенез активируя экспрессию каскада нейрональных генов, но они подавляют экспрессию глиальный гены.[15] Нейронный bHLH гены выполняют разные функции в зависимости от: чувствительности к боковое торможение, который определяет, становится ли клетка эпидермальной или нейрональной и экспрессируется ли ген в ЦНС до или после терминала митоз.[4]
Проневральные гены способствуют нейрогенез и подавлять глиогенез но некоторые нейрогенные факторы могут регулировать оба этих процесса в зависимости от концентрации пронейральных генов. Например, BMP (костные морфогенетические белки) продвигать нейрогенез у предшественников, которые экспрессируют высокий уровень Нейрогенин-1 и глиогенез у предшественников, которые экспрессируют низкие уровни Нейрогенин-1.[5]Глиогенез процессы зависят от низких концентраций или отклонения пронейральных генов и могут быть ускорены в зависимости от того, какие пронейральные гены затронуты.[16]
У беспозвоночных
В Дрозофила, пронейральные гены сначала экспрессируются в эктодермальный клетки, обладающие как эпидермальным, так и нейрональным потенциалом. Пронейральная активность приводит к отбору предшественников, которые преданы нервной судьбе, но остаются мультипотентный, с предшественниками органов чувств, дающими начало нейроны, глия и другие типы ненейрональных клеток. Кроме того, некоторые нейробласты из Центральная нервная система также генерировать как нейроны и глия. Прародители периферийный и Центральная нервная система только начинают делиться после того, как экспрессия пронейральных генов снижается.[1]
У позвоночных
Проневральные гены сначала экспрессируются в нейроэпителиальных клетках, которые уже определены для нервной судьбы и самообновляются. Проневральная активность приводит к образованию и отслаиванию предшественников, которые ограничены судьбой нейронов и имеют ограниченный митотический потенциал. По крайней мере, в некоторых клонах пронейральные гены участвуют в приверженности нейральных предшественников судьбе нейронов за счет глиальный судьба.[1]
В процессе бокового торможения

Боковое торможение это межклеточное взаимодействие который происходит в пронейральном кластере, чтобы определить и ограничить клетки, которые вызывают нейробласт.[16] Во время этого взаимодействия зарождающиеся нейробласты экспрессируют пронейральные гены выше определенного порога, и в то же время они экспрессируют связанный с мембраной лиганд, называемый «дельта», который связывает и активирует Notch рецепторы экспрессируются в соседних клетках. Один раз Notch активируется, активность пронейральных генов в этих клетках снижается, вероятно, из-за активации генов в «’ энхансере расщепленного комплекса E (spl) ’’, кодирующем ингибиторные bHLH факторы транскрипции.[17] При ингибировании пронейральные гены предотвращают превращение клеток в нейронные, но также снижают их уровни «дельты». Эти конкретные взаимодействия ограничивают пронейральную активность одной клеткой в каждом пронейральном кластере, что приводит к паттерну соли и перца.[2][16]
Не все пронейральные гены одинаково чувствительны к боковое торможение. Например, в Xenopus, Читнис и Кинтнер продемонстрировали, что «XASH-3» и NeuroD (комплекс ахэте-щиток ) по-разному реагировать на боковое торможение, которые отражают различную способность активировать гены-мишени и различную восприимчивость этих генов-мишеней к репрессии посредством выемка.[16] Предварительные исследования показали, что даже когда Путь передачи сигналов Notch / Delta заблокирован, Wnt2b способен подавлять дифференцировку нейронов через подавление из мРНК экспрессия нескольких пронейральных генов, а также Notch1. С этим механизмом Wnt2b поддерживает недифференцированные клетки-предшественники за счет ослабления экспрессии пронейральных и нейрогенных генов, предотвращая попадание клеток в каскад дифференцировки, регулируемый пронейральными генами и Notch.[18] Несмотря на то что сигнализация выемки участвует в контроле экспрессии пронейральных генов, петли положительной обратной связи необходимы для увеличения или поддержания уровней пронейральных генов. В факторы транскрипции Ответственный за это обслуживание может действовать путем подавления сигнальный путь notch в конкретных клетках или на посттранскрипционном уровне, влияя на пронейральные гены транскрипция и функция.[1]
В нейрогенезе
Нейрогенез в нервной системе беспозвоночных.
В беспозвоночные пронейральные гены, особенно члены комплекс ахэте-щиток (AS-C) продвигать нейрогенез, в то время как нейрогенные гены предотвращают нейрогенез и облегчить эпидермальное развитие. Формирование нейробласты зависит от Ахето-щитовой комплекс гены - ачаэте (ас), щиток (sc), смертоносный щиток (lsc) и дефект вентральной нервной системы (‘’ vnd ’’). Однако только «vnd» может контролировать этот процесс формирования, потому что этот ген активирует экспрессию других генов.[19][20] ac, sc, lsc факторы первоначально экспрессируются в зародышах центральной нервной системы эмбриона (нейроэктодерма ) в пронейральных кластерах, из которых одиночные нейробласты позже возникнут.[20] Каждая клетка пронейрального кластера имеет общий потенциал формирования нейробластов. Местное угнетение оставшихся клеток за счет увеличения нейробласты гарантирует, что только один нейробласт возникает из пронейрального кластера. Все ячейки кластера сохраняют свой потенциал формирования NB, по крайней мере, пока NB увеличивается, но теряют этот потенциал к тому времени, когда ячейка собирается делиться. Паттерны экспрессии пронейральных генов приводят к различным способам нейробласты образование в голове и туловище. Коэкспрессия пронейральных генов в мозг нейробласты преходящ и меняется в зависимости от стадии развития.[21]
Нейрогенез стоматогастральной нервной системы при Дрозофила
Экспрессия пронейральных генов в нейроэктодермальный клетки, составляющие пронейральные кластеры, делают их способными расслаиваться как нейробласты. Несмотря на то что нейробласты являются предшественниками Дрозофилы Центральная нервная система (ЦНС), экспрессия пронейрального гена также участвует в контрольной спецификации и морфогенез из стоматогастральный предшественники нервных клеток. Эти гены экспрессируются и необходимы на всех этапах стоматогастральная нервная система (SNS), чтобы регулировать количество, структуру и структурные характеристики субпопуляций SNS. Правильный баланс между пронейральной и нейрогенной экспрессией генов в SNS placodes участвует в контроле сложной последовательности морфогенетических движений (расслоение, инвагинация и диссоциация), посредством которых эти плакоды вызывают различные SNS субпопуляции.[22]
Нейрогенез в центральной нервной системе позвоночных
В Центральная нервная система не все bHLH гены участвуют в нейрогенез потому что NeuroD и семейства '' Math3 / NeuroM '' также участвуют в решении судьбы нейрональных и глиальных клеток. Другое про-нейронное семейство (которое включает '' math1 '' и '' math5 '') важно для развития небольшого числа нейронных линий, тогда как '' math1 '' также играет роль в спецификации интернейрон личность. Типы ячеек, зависящие от выражения «math1’ ’, относятся к проприоцептивный сенсорный путь.[1] Бертран и др. (2002) подтвердили пронейральную активность «mash1», ngn1 и ngn2 и, возможно, math1 и «math5» в мыши.[1] Нейрогенез в Центральная нервная система зависит от ингибирования пронейрального гена Notch сигнальный путь а отсутствие этого ключевого регулятора приводит к преждевременной дифференциации нейроны. Для поддержания нейральных клеток-предшественников между соседними клетками возникает регуляторная петля, которая включает боковое торможение процесс (см. боковое торможение).[23] В отсутствие Боковое торможение некоторые пронейральные гены, такие как ASCL1 или '' neuroG '' способны индуцировать экспрессию нейрон-специфических генов, приводящую к преждевременному образованию ранних нейронов.[24] Ratié и коллеги (2013) составили, что Notch пронейральная генная сеть играет важную роль в обновлении и переходе клеточной судьбы у мышей.[23]
Нейрогенез в периферической нервной системе позвоночных
в периферическая нервная система, NGNS участвуют в определении всех черепных и спинных сенсорных предшественников. Проневральные гены, такие как пюре1, ngn1 и ngn2 в основном экспрессируются в большинстве предшественников спинной мозг, а также коэкспрессируются в спинном конечный мозг. Вместе эти группы bHLH факторы способствуют генерации всех кора головного мозга прародители. Маш1 единственный ген, экспрессирующийся в брюшной конечный мозг. Однако на вентральном и дорсальном концах нервная трубка экспрессируется другой тип пронейральных генов, например ngn3 и ‘’ math1 ’’.[1]
В глиогенезе
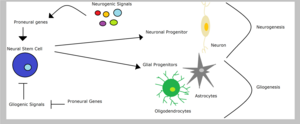
Нервные стволовые клетки могут давать начало нейрональным или глиальным предшественникам, в зависимости от типа сигналов, которые они получают - глиогенных или нейрогенных сигналов соответственно. Глиал клетки-предшественники могли дифференцироваться в олигодендроциты или же астроциты. Однако предопределение клонов нейральных предшественников включает подавление альтернативных судеб. Следовательно, позвоночное животное пронейральные гены способствуют судьбе нейронов и одновременно ингибируют глиальный судьбы. Например, подавление экспрессии пронейрального гена ngn2 в спинной мозг подавляет олигодендроцит дифференциация. В контексте ограниченного глиальный предшественников, пронейральные гены могут иметь функции, отличные от их лучше охарактеризованной роли в спецификации клонов, возможно, в дифференцировке глиальные линии.[1]Сан и его коллеги показали, что пронейральная ngn1 подавляет глиогенез путем связывания транскрипционного соактиваторы подобно CBP /Smad1 или же p300 /Smad1 предотвращение транскрипции глиальная дифференцировка гены.[15]С другой стороны, Notch сигнальный путь способен продвигать глиогенез в стволовые клетки через ингибирование пронейральных генов, таких как пюре1 и нейрогенины.[5]
В судьбе регуляции клеточного цикла
В позвоночные, хотя пронейральные гены определяют нейральную судьбу предшественников, они также способствуют остановке стадии их деления путем изоляции уже определенных клеток-предшественников от влияния внешних определяющих судьбу сигналов. Проневральные гены регулируют клеточный цикл путем активации циклин-зависимая киназа (‘’ Cdk ’’) ингибиторов в некоторых линиях на уровне генов дифференцировки нейронов. С другой стороны, в беспозвоночные подобно Дрозофила, пронейральные гены экспрессируются в основном в неделящихся клетках, но могут также экспрессироваться в делящихся клетках, где Ахето-щитовой комплекс пронейральные гены ингибируют клеточный цикл прогрессия.[1]
В развитии органов чувств
Проневральные гены также играют важную роль в развитии различных типов органы чувств, а именно хордотональные органы (проприорецепторы, которые обнаруживают механические и звуковые колебания) и внешние органы чувств. Члены комплекс ахэте-щиток, Такие как ачаете и щиток, а также "атональный" и "без дочери" эктодермальные клетки способность становиться сенсорными материнскими клетками (SMC).[6][10] В развитии сенсорный органов есть две основные фазы: определение и дифференциация, которые не могут быть механически разделены. Проневральные белки участвуют в обоих процессах за счет активации нижележащих «дифференцирующих генов».[25] которые, в свою очередь, регулируют индукцию характеристик подтипа сенсорных органов. Спецификация органы чувств пронейральными генами - сложный процесс, поскольку они вызывают разные клеточные контексты. Например, в Дрозофила, атонал (ато) может способствовать развитию хордотональные органы, для рецепторов органы обоняния, в зависимости от имагинальный диск в котором это выражено.[1] В Дрозофилы эмбриогенез, пронейральный ген ачаете выражается в четко определенных областях, как в энтодерма, неся ответственность за формирование особого органы чувств у взрослых особей и личинок. Согласно Ruíz-Gomes and Ghysen (1993), эта экспрессия происходит в двух различных фазах: компетентное состояние, в котором пронейральный ген экспрессируется в кластере клеток; детерминированное состояние, в котором конкретная клетка накапливает высокие уровни транскриптов «ac», производя нейронный предшественник.[26] Функция каждого из Комплекс АСК гены варьируются в зависимости от состояния развития (личинки или взрослые).[26] Например, во взрослом состоянии ac и sc гены способствуют дифференцировке двух наборов дополнительных органы чувств и ас ген как минимальная функция, а в состоянии личинки ac и sc гены влияют на один и тот же набор органы чувств и ас отвечает за определение дополнительного набора.[27]
В кортикогенезе
в неокортекс существует широкая нейронная сеть, поддерживаемая астроциты и олигодендроциты (глиальные клетки ) с разными функциями. Во время коркового развития bHLH Факторы контролируют пролиферацию и дифференцировку нервных клеток, а их функции в любое время и в любом месте зависят от их клеточного контекста. NeuroD, NGNS, Маш, «Олиг» и другие семейства пронейральных генов играют решающую роль в решении судьбы клетки во время кортикогенез и различные их комбинации регулируют выбор и время дифференциации в нейрон, астроцит или олигодендроцит.[28]Высокий уровень Ngn1 и Ngn2 необходимы для определения нейрональной идентичности корковых предшественников только на ранних стадиях неокортекс разработка.[29]Особенно, ngn2 также важен для регулирования перехода корковых клеток-предшественников из желудочковой зоны в субвентрикулярная зона. С другой стороны, пюре1 участвует в ранней дифференциации полосатое тело нейроны[30] и достаточного для ускорения деления базальных клеток независимо от его роли в спецификации судьбы нейрональных клеток на более поздних стадиях. Сотрудничество между ngn2 и пюре1 пронейральные гены регулируют переход кортикальных предшественников из апикальных в базальные клеточные компартменты.[28] Спецификация различных подтипов нейронов зависит от группы вовлеченных пронейральных генов.[31] Низкий уровень пронейральной стенограммы в желудочковой зоне экспрессируются, когда происходит спецификация предшественников, и увеличение их экспрессии приводит к началу нейрогенез. NGNS несут ответственность за формирование глутаматергический нейроны в то время как пюре1 дает начало ГАМКергический и холинергический нейроны.[31]
В Дрозофила развитие фоторецепторов
Проневральный белок Atonal (Ato) отвечает за развитие Дрозофилы R8 фоторецепторы. Однако он не действует в одиночку, так как димеризуется вторым идентичным белком, «Daughterless (Da)», который усиливает его экспрессию. Совместное выражение «Ато» и «Да» важно для миграции разных типов клеток омматидий и для репрессии атонала в межкластерных пространствах, функционирующего как тормозные сигналы, которые регулируют как количество, так и положение возникающих клеток R8. Более того, это выражение приводит к активации киназа MAPK что важно для набора клеток и репрессорного эффекта, когда это киназа неактивен, выражение «Ато» обнаруживается в каждой клетке.[32][33] В исходном состоянии Ежик (‘’ Hh ’’) и Декапентаплегический отвечают за активацию ‘’ ato ’’ в небольших клеточных кластерах, что приводит к активации MAPK. Первый, Ежик вызывает экспрессию ‘’ ’ato’ ’не только в клетках, которые в конечном итоге становятся фоторецепторные клетки, поэтому его экспрессия должна быть уточнена и ограничена одной предполагаемой клеткой R8. Ежик сигнализация также требуется для подавления «атонального» выражения между зарождающимися пронейральными кластерами, что обнаруживает двойную роль, имеющую решающее значение для построения точности и геометрии у взрослого сетчатка. Следовательно, регуляция экспрессии ‘’ ato ’’ зависит от уровней Ежик: на низких уровнях (то есть далеко от источника) этот пронуклеарный ген активируется, а на высоких уровнях (близко к источнику) он подавляется. Объединение боковое торможение с двойной ролью Ежик, можно представить, как можно структурировать гексагональный массив ячеек R8.[33]
Ненейронные функции
Помимо их роли в нервная система развития, пронейральные гены также участвуют в процессах, связанных с вторжение трофобласта, эндокринный дифференцировка клеток (а именно в поджелудочная железа и аденогипофиз ), определение пола и легкие, щитовидная железа, надпочечник и слюна развитие желез и желудочно-кишечного тракта.[34]
Во вторжении человеческого трофобласта
Помимо их участия в дифференцировке нейронов и глии, определение пола и органы чувств развития, пронейральные гены также участвуют в трофобласт дифференциация при прогрессировании инвазии, в плацентарный формирование. Исследования выявили проявление нейроД1, нейроD2 и транскрипты "ath2" в различных подгруппах инвазивных трофобласт.[35]
В развитии поджелудочной железы
В развивающейся поджелудочной железе фактор транскрипции ngn3 ’Отмечает популяции клеток, которые переходят от недифференцированных эпителиальных клеток-предшественников к зрелым эндокринный клетки (предшественники эндокринных клеток поджелудочной железы) и, следовательно, не экспрессируют гормоны тем не менее, что говорит о том, что этот ген отключен в дифференцированном положительном гормональном состоянии.[36] ngn3 ’’ Необходимо и достаточно, чтобы стимулировать формирование островковые клетки во время развития поджелудочной железы, аналогично спецификации нервной судьбы у нейроэктодерма.[37]
В определении пола
Несмотря на то что нейрогенез и определение пола кажутся разными биологическими процессами, есть свидетельства того, что '' бездочность (da '') - важный ген для формирования всего Дрозофила периферическая нервная система - также требуется для правильного определение пола.[38] В мухах, щиток работает, чтобы направлять развитие нейронов, но этот ген также приобрел роль в первичном событии определение пола – Х хромосома подсчет - став сигнальным элементом Х-хромосомы (XSE).[39]
В миогенезе
Хотя пронейральные гены действуют в эктодерма, смертоносный щиток действует в соматическая мезодерма определить кластер ячеек, из которого мышца прародители будут выделены.[40] Взаимодействие между этими клетками и эктодерма, приводит к образованию клеток-основателей мышц по аналогии с тем, что происходит в Центральная нервная система.[41]
В клеточной миграции
ngn1 и ngn2 может самостоятельно регулировать механизмы миграция клеток и участвуют в начальных подавление из RhoA прямо перед тем, как нейральные клетки-предшественники становятся постмитотическими, тогда как нейроД в первую очередь участвует в непрерывном подавлении RhoA во время корковой миграции. Другой bHLH такие факторы, как ‘’ math2 ’’, нейроD2 и ‘’ nscl1 ’’ может подавлять RhoA экспрессии и регулируют механизм миграции в постмитотических нейроны.[42]
Рекомендации
- ^ а б c d е ж грамм час я j k л м п Bertrand, N; Castro, D. S .; Guillemot, F (2002). «Проневральные гены и спецификация типов нервных клеток». Обзоры природы Неврология. 3 (7): 517–30. Дои:10.1038 / nrn874. PMID 12094208.
- ^ а б Кунищ, М; Haenlin, M; Кампос-Ортега, Дж. А. (1994). «Боковое ингибирование, опосредованное дельта нейрогенного гена дрозофилы, усиливается пронейральными белками». Труды Национальной академии наук Соединенных Штатов Америки. 91 (21): 10139–43. Дои:10.1073 / пнас.91.21.10139. ЧВК 44973. PMID 7937851.
- ^ Тепасс, У; Хартенштейн, V (1995). «Нейрогенные и пронейральные гены контролируют спецификацию клеточной судьбы в энтодерме дрозофилы». Разработка. 121 (2): 393–405. PMID 7768181.
- ^ а б c Brunet, J. F .; Гайзен, А. (1999). «Деконструкция детерминации клеток: пронейральные гены и идентичность нейронов». BioEssays. 21 (4): 313–8. Дои:10.1002 / (SICI) 1521-1878 (199904) 21: 4 <313 :: AID-BIES7> 3.0.CO; 2-C. PMID 10377893.
- ^ а б c Моррисон, С. Дж. (2001). «Нейрональная дифференцировка: пронейральные гены подавляют глиогенез». Текущая биология. 11 (9): R349–51. Дои:10.1016 / S0960-9822 (01) 00191-9. PMID 11369245.
- ^ а б Chien, C.T .; Hsiao, C.D .; Jan, L. Y .; Ян, Ю. Н. (1996). "Информация о типе нейронов, закодированная в домене" основная спираль-петля-спираль "пронейральных генов". Труды Национальной академии наук Соединенных Штатов Америки. 93 (23): 13239–44. Дои:10.1073 / пнас.93.23.13239. ЧВК 24077. PMID 8917575.
- ^ Ledent, V; Паке, О; Вервурт, М (2002). «Филогенетический анализ основных белков спирали-петли-спирали человека». Геномная биология. 3 (6): ИССЛЕДОВАНИЕ0030. Дои:10.1186 / gb-2002-3-6-research0030. ЧВК 116727. PMID 12093377.
- ^ Гизен, А; Дамбли-Шодьер, С. (1988). «От ДНК к форме: комплекс achaete-scute». Гены и развитие. 2 (5): 495–501. Дои:10.1101 / gad.2.5.495. PMID 3290049.
- ^ а б González, F; Романи, S; Кубас, П; Модолелл, Дж; Кампузано, S (1989). «Молекулярный анализ гена asense, члена комплекса achaete-scute у Drosophila melanogaster, и его новая роль в развитии зрительной доли». Журнал EMBO. 8 (12): 3553–62. ЧВК 402034. PMID 2510998.
- ^ а б Jarman, A. P .; Грау, Y; Jan, L. Y .; Ян, Ю. Н. (1993). «Атонал - пронейральный ген, который управляет формированием хордотональных органов в периферической нервной системе дрозофилы». Клетка. 73 (7): 1307–21. Дои:10.1016 / 0092-8674 (93) 90358-в.. PMID 8324823.
- ^ Goulding, S.E .; Уайт, Н. М .; Джарман, А. П. (2000). «Като кодирует основной транскрипционный фактор спираль-петля-спираль, участвующий в правильной дифференцировке органов чувств дрозофилы». Биология развития. 221 (1): 120–31. Дои:10.1006 / dbio.2000.9677. PMID 10772796.
- ^ Jan, Y. N .; Ян, Л. Ю. (1993). «Белки HLH, нейрогенез мух и миогенез позвоночных». Клетка. 75 (5): 827–30. Дои:10.1016 / 0092-8674 (93) 90525-у. PMID 8252617.
- ^ Кампос-Ортега, Дж. А. (1993). «Механизмы раннего нейрогенеза у Drosophila melanogaster». Журнал нейробиологии. 24 (10): 1305–27. Дои:10.1002 / neu.480241005. PMID 8228961.
- ^ Powell, L.M .; Джарман, А. П. (2008). «Контекстная зависимость пронейральных белков bHLH». Текущее мнение в области генетики и развития. 18 (5): 411–7. Дои:10.1016 / j.gde.2008.07.012. ЧВК 3287282. PMID 18722526.
- ^ а б Солнце, Y; Надаль-Висенс, М; Misono, S; Lin, M. Z .; Зубяга, А; Хуа, X; Fan, G; Гринберг, М. Э. (2001). «Нейрогенин способствует нейрогенезу и ингибирует дифференцировку глии независимыми механизмами». Клетка. 104 (3): 365–76. Дои:10.1016 / S0092-8674 (01) 00224-0. PMID 11239394.
- ^ а б c d Читнис, А; Кинтнер, К. (1996). «Чувствительность пронейральных генов к латеральному торможению влияет на структуру первичных нейронов у эмбрионов Xenopus». Разработка. 122 (7): 2295–301. PMID 8681809.
- ^ Schlatter, R .; Майер, Д. (2005). «Усилитель расщепления комплексов дрозофилид и комплексов ахет-щиток, полученных из простых ур-комплексов, сохраненных в комарах и пчелах». BMC Эволюционная биология. 5: 67. Дои:10.1186/1471-2148-5-67. ЧВК 1310631. PMID 16293187.
- ^ Кубо, Ф; Takeichi, M; Накагава, S (2005). «Wnt2b ингибирует дифференцировку клеток-предшественников сетчатки в отсутствие активности Notch, подавляя экспрессию пронейральных генов». Разработка. 132 (12): 2759–70. Дои:10.1242 / dev.01856. PMID 15901663.
- ^ Skeath, J. B .; Panganiban, G.F .; Кэрролл, С. Б. (1994). «Дефектный ген вентральной нервной системы контролирует экспрессию пронейрального гена на двух различных стадиях во время образования нейробластов у дрозофилы». Разработка. 120 (6): 1517–24. PMID 8050360.
- ^ а б Skeath, J. B .; Кэрролл, С. Б. (1992). «Регулирование экспрессии пронейральных генов и клеточной судьбы во время сегрегации нейробластов в эмбрионе Drosophila». Разработка. 114 (4): 939–46. PMID 1618155.
- ^ Urbach, R; Schnabel, R; Технау, Г. М. (2003). «Паттерн образования нейробластов, митотических доменов и экспрессии пронейральных генов во время раннего развития мозга у дрозофилы». Разработка. 130 (16): 3589–606. Дои:10.1242 / dev.00528. PMID 12835378.
- ^ Hartenstein, V; Тепасс, У; Грушински-Дефео, Э (1996). «Проневральные и нейрогенные гены контролируют спецификацию и морфогенез предшественников стоматогастральных нервных клеток у дрозофилы». Биология развития. 173 (1): 213–27. Дои:10.1006 / dbio.1996.0018. PMID 8575623.
- ^ а б Соотношение, л; Посуда, М; Барлой-Хаблер, Ф; Romé, H; Gicquel, I; Dubourg, C; Дэвид, V; Дюпе, V (2013). «Новые гены активируются, когда передача сигналов NOTCH нарушается во время развития гипоталамуса». Нейронное развитие. 8: 25. Дои:10.1186/1749-8104-8-25. ЧВК 3880542. PMID 24360028.
- ^ Kageyama, R .; Ohtsuka, T .; Shimojo, H .; Имаёши, И. (2008). «Динамическая передача сигналов Notch в нейральных клетках-предшественниках и пересмотренный взгляд на латеральное ингибирование». Природа Неврология. 11 (11): 1247–1251. Дои:10.1038 / №2208. PMID 18956012.
- ^ Кубас, П; Де Селис, Дж. Ф .; Кампузано, S; Modolell, J (1991). «Проневральные скопления экспрессии achaete-scute и формирование сенсорных органов в имагинальном крыловом диске дрозофилы». Гены и развитие. 5 (6): 996–1008. Дои:10.1101 / gad.5.6.996. PMID 2044965.
- ^ а б Руис-Гомес, М; Гайзен, А. (1993). «Экспрессия и роль пронейрального гена, achaete, в развитии личиночной нервной системы дрозофилы». Журнал EMBO. 12 (3): 1121–30. ЧВК 413313. PMID 8458326.
- ^ Dambly-Chaudiere, C .; Гайзен, А. (1987). «Независимые субпаттерны органов чувств требуют независимых генов комплекса ахето-щиток у личинок дрозофилы». Гены и развитие. 1 (3): 297–306. Дои:10.1101 / gad.1.3.297.
- ^ а б Ross, S.E .; Гринберг, М. Э .; Стайлз, К. Д. (2003). "Основные факторы спирали-петли-спирали в развитии коры". Нейрон. 39 (1): 13–25. Дои:10.1016 / S0896-6273 (03) 00365-9. PMID 12848929.
- ^ Schuurmans, C; Арман, О; Nieto, M; Стенман, Дж. М .; Britz, O; Кленин, Н; Браун, C; Langevin, L.M .; Зейбт, Дж; Тан, H; Cunningham, J.M .; Дайк, Р. Уолш, К; Кэмпбелл, К; Polleux, F; Guillemot, F (2004). «Последовательные фазы корковой спецификации включают нейрогенин-зависимые и независимые пути». Журнал EMBO. 23 (14): 2892–902. Дои:10.1038 / sj.emboj.7600278. ЧВК 514942. PMID 15229646.
- ^ Юн, К; Фишман, S; Джонсон, Дж; Hrabe De Angelis, M; Weinmaster, G; Рубинштейн, Дж. Л. (2002). «Модуляция передачи сигналов notch с помощью Mash1 и Dlx1 / 2 регулирует последовательную спецификацию и дифференцировку типов клеток-предшественников в подкорковом телеэнцефалоне». Разработка. 129 (21): 5029–40. PMID 12397111.
- ^ а б Britz, O; Маттар, П; Nguyen, L; Langevin, L.M .; Циммер, К; Алам, S; Guillemot, F; Шурманс, C (2006). «Роль пронейральных генов в созревании корковых клеток-предшественников». Кора головного мозга. 16 Дополнение 1: i138–51. Дои:10.1093 / cercor / bhj168. PMID 16766700.
- ^ Chen, C.K .; Чиен, К. Т. (1999). «Отрицательная регуляция атонала в формировании пронейрального кластера фоторецепторов Drosophila R8». Труды Национальной академии наук Соединенных Штатов Америки. 96 (9): 5055–60. Дои:10.1073 / пнас.96.9.5055. ЧВК 21815. PMID 10220417.
- ^ а б Домингес, М. (1999). «Двойная роль Hedgehog в регуляции пронейрального гена атонального во время развития омматидий». Разработка. 126 (11): 2345–53. PMID 10225994.
- ^ Pogoda, H.M .; фон Дер Хардт, S; Герцог, Вт; Kramer, C; Schwarz, H; Хаммершмидт, М. (2006). «Проневральный ген ascl1a необходим для эндокринной дифференцировки и выживания клеток в аденогипофизе рыбок данио». Разработка. 133 (6): 1079–89. Дои:10.1242 / dev.02296. PMID 16481349.
- ^ Westerman, B.A .; Поутсма, А; Маруяма, К. Schrijnemakers, H.F .; Van Wijk, I.J .; Аудеянс, К. Б. (2002). «Проневральные гены NEUROD1 и NEUROD2 экспрессируются во время инвазии трофобласта человека». Механизмы развития. 113 (1): 85–90. Дои:10.1016 / S0925-4773 (01) 00665-7. PMID 11900979.
- ^ Johansson, K. A .; Dursun, U .; Jordan, N .; Gu, G .; Beermann, F .; Gradwohl, G.R .; Грейпин-Боттон, А. (2007). «Временной контроль активности нейрогенина 3 в предшественниках поджелудочной железы выявляет окна компетентности для генерации различных типов эндокринных клеток». Клетка развития. 12 (3): 457–465. Дои:10.1016 / j.devcel.2007.02.010. PMID 17336910.
- ^ Lee, J.C .; Smith, S. B .; Ватада, H; Lin, J; Шил, D; Ван, Дж; Mirmira, R.G .; Герман, М. С. (2001). «Регуляция проэндокринного гена поджелудочной железы нейрогенин3». Сахарный диабет. 50 (5): 928–36. Дои:10.2337 / диабет.50.5.928. PMID 11334435.
- ^ Клайн, Т. В. (1976). «Зависящий от пола, чувствительный к температуре материнский эффект бездочечной мутации Drosophila melanogaster». Генетика. 84 (4): 723–42. ЧВК 1213604. PMID 827461.
- ^ Wrischnik, L.A .; Timmer, J. R .; Megna, L.A .; Клайн, Т. В. (2003). «Привлечение пронейрального гена к пути определения пола у дрозофилы». Генетика. 165 (4): 2007–27. ЧВК 1462923. PMID 14704182.
- ^ Кармена, А; Бейт, М; Хименес, Ф (1995). «Летальный щиток, пронейральный ген, участвует в спецификации мышечных предшественников во время эмбриогенеза дрозофилы». Гены и развитие. 9 (19): 2373–83. Дои:10.1101 / gad.9.19.2373. PMID 7557389.
- ^ Bate, C.M .; Грюневальд, Э. Б. (1981). «Эмбриогенез нервной системы насекомых II: второй класс клеток-предшественников нейронов и происхождение межсегментарных соединительных элементов». Журнал эмбриологии и экспериментальной морфологии. 61: 317–30. PMID 7264548.
- ^ Ge, Вт; Он, F; Kim, K. J .; Бланки, Б; Coskun, V; Nguyen, L; Ву, Х; Чжао, Дж; Heng, J. I .; Мартинович, К; Тао, Дж; Wu, H; Кастро, Д; Sobeih, M. M .; Corfas, G; Gleeson, J. G .; Гринберг, М. Э .; Guillemot, F; Солнце, Ю. Э. (2006). «Связь миграции клеток с нейрогенезом с помощью пронейральных факторов bHLH». Труды Национальной академии наук. 103 (5): 1319–24. Дои:10.1073 / pnas.0510419103. ЧВК 1345712. PMID 16432194.