Рецептор Fc - Fc receptor
| Иммуноглобулиноподобный рецептор | |
|---|---|
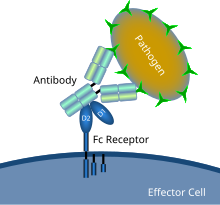 Схематическая диаграмма, показывающая взаимодействие рецептора Fc с микробным патогеном, покрытым антителами | |
| Идентификаторы | |
| Символ | Рецептор Fc |
| Мембранома | 10 |
А Рецептор Fc белок, обнаруженный на поверхности определенных клеток, в том числе, среди прочего, В-лимфоциты, фолликулярные дендритные клетки, естественные клетки-киллеры, макрофаги, нейтрофилы, эозинофилы, базофилы, человек тромбоциты, и тучные клетки - которые способствуют защитным функциям иммунная система. Его имя происходит от его специфичности связывания для части антитело известный как Fc (фрагмент кристаллизующийся) область. Рецепторы Fc связываются с антителами, которые прикрепляются к инфицированным клеткам или вторгаются патогены. Их деятельность стимулирует фагоцитарный или же цитотоксический клетки для разрушения микробы, или инфицированные клетки опосредованным антителами фагоцитоз или же антителозависимая клеточно-опосредованная цитотоксичность. Немного вирусы Такие как флавивирусы используют рецепторы Fc, чтобы помочь им инфицировать клетки с помощью механизма, известного как антителозависимое усиление инфекции.[1]
Классы
Существует несколько различных типов рецепторов Fc (сокращенно FcR), которые классифицируются на основе тип антитела что они признают. Латинская буква, используемая для идентификации типа антитела, преобразуется в соответствующую греческую букву, которая помещается после части имени «Fc». Например, те, которые связывают наиболее распространенный класс антител, IgG, называются Fc-гамма-рецепторами (FcγR), те, которые связывают IgA называются рецепторами Fc-альфа (FcαR), а те, которые связывают IgE называются рецепторами Fc-эпсилон (FcεR). Классы FcR также различаются по экспрессирующим их клеткам (макрофаги, гранулоциты, естественные клетки-киллеры, Т- и В-клетки) и по сигнальным свойствам каждого рецептора.[2]
Fc-гамма рецепторы
Все рецепторы Fcγ (FcγR) принадлежат к суперсемейство иммуноглобулинов и являются наиболее важными рецепторами Fc для индукции фагоцитоз из опсонизированный (отмечены) микробы.[3] Это семейство включает несколько членов: FcγRI (CD64), FcγRIIA (CD32 ), FcγRIIB (CD32), FcγRIIIA (CD16a), FcγRIIIB (CD16b), которые различаются по своим антителам. родство из-за их разных молекулярный структура.[4] Например, FcγRI связывается с IgG сильнее, чем FcγRII или FcγRIII. FcγRI также имеет внеклеточный часть состоит из трех иммуноглобулин (Ig) -подобные домены, на один домен больше, чем у FcγRII или FcγRIII. Это свойство позволяет FcγRI связывать единственную молекулу IgG (или мономер ), но все рецепторы Fcγ должны связывать несколько молекул IgG в пределах одного иммунный комплекс для активации.[5]
Рецепторы Fc-гамма различаются по своему сродству к IgG, и аналогично разные подклассы IgG обладают уникальным сродством к каждому из рецепторов Fc гамма.[6] Эти взаимодействия дополнительно регулируются гликаном (олигосахаридом) в положении CH2-84.4 IgG.[6] Например, создавая стерические препятствия, фукоза, содержащая гликаны CH2-84.4, снижает сродство IgG к FcγRIIIA.[6] Напротив, гликаны G0, которые лишены галактозы и заканчиваются вместо этого фрагментами GlcNAc, обладают повышенным сродством к FcγRIIIA.[6]
Другой FcR экспрессируется на нескольких типах клеток и по структуре похож на MHC класс I. Этот рецептор также связывает IgG и участвует в сохранении этого антитела.[7] Однако, поскольку этот рецептор Fc также участвует в передаче IgG от матери либо через плацента ей плод или в молоко к ее грудному ребенку младенец, это называется неонатальный Рецептор Fc (FcRn ).[8][9] Недавние исследования показали, что этот рецептор играет роль в гомеостазе сывороточных уровней IgG.
Fc-альфа рецепторы
Только один рецептор Fc принадлежит к подгруппе FcαR, который называется FcαRI (или CD89).[10] FcαRI находится на поверхности нейтрофилы, эозинофилы, моноциты, некоторые макрофаги (в том числе Клетки Купфера ), и немного дендритные клетки.[10] Он состоит из двух внеклеточных Ig-подобных доменов и является членом обоих суперсемейство иммуноглобулинов и семейство рецепторов многоцепочечного иммунного распознавания (MIRR).[3] Он сигнализирует, связываясь с двумя сигнальными цепями FcRγ.[10] Другой рецептор также может связывать IgA, хотя он имеет более высокое сродство к другому антителу, называемому IgM.[11] Этот рецептор называется рецептором Fc-альфа / мю (Fcα / μR) и относится к типу I. трансмембранный белок. С одним Ig-подобным доменом во внеклеточной части этот рецептор Fc также является членом суперсемейства иммуноглобулинов.[12]
Fc-эпсилон рецепторы
Известны два типа FcεR:[3]
- рецептор с высоким сродством FcεRI является членом суперсемейства иммуноглобулинов (имеет два Ig-подобных домена). FcεRI находится на эпидермальном Клетки Лангерганса, эозинофилы, тучные клетки и базофилы.[13][14] В результате своего клеточного распределения этот рецептор играет важную роль в контроле аллергические реакции. FcεRI также экспрессируется на антигенпрезентирующие клетки, и контролирует выработку важных иммунных медиаторов, называемых цитокины которые продвигают воспаление.[15]
- рецептор низкой аффинности FcεRII (CD23 ) это Лектин С-типа. FcεRII выполняет множество функций в качестве мембраносвязанного или растворимого рецептора; он контролирует рост и дифференцировку В-клеток и блокирует связывание IgE эозинофилов, моноцитов и базофилов.[16]
Таблица результатов
| Имя рецептора | Лиганд основного антитела | Сродство к лиганду | Распределение клеток | Эффект после связывания с антителом |
| FcγRI (CD64 ) | IgG1 и IgG3 | Высокая (Kd ~ 10−9 M) | Макрофаги Нейтрофилов Эозинофилы Дендритные клетки | Фагоцитоз Активация клетки Активация респираторного взрыва Индукция уничтожения микробов |
| FcγRIIA (CD32 ) | IgG | Низкий (Kd> 10−7 M) | Макрофаги Нейтрофилов Эозинофилы Тромбоциты Клетки Лангерганса | Фагоцитоз Дегрануляция (эозинофилы) |
| FcγRIIB1 (CD32) | IgG | Низкий (Kd> 10−7 M) | В-клетки Тучные клетки | Нет фагоцитоза Подавление клеточной активности |
| FcγRIIB2 (CD32) | IgG | Низкий (Kd> 10−7 M) | Макрофаги Нейтрофилов Эозинофилы | Фагоцитоз Подавление клеточной активности |
| FcγRIIIA (CD16a ) | IgG | Низкий (Kd> 10−6 M) | NK-клетки Макрофаги (определенные ткани) | Индукция антителозависимая клеточно-опосредованная цитотоксичность (ADCC) Индукция высвобождения цитокинов макрофагами |
| FcγRIIIB (CD16b ) | IgG | Низкий (Kd> 10−6 M) | Эозинофилы Макрофаги Нейтрофилов Тучные клетки Фолликулярные дендритные клетки | Индукция уничтожения микробов |
| FcεRI | IgE | Высокая (Kd ~ 10−10 M) | Тучные клетки Эозинофилы Базофилы Клетки Лангерганса Моноциты | Дегрануляция Фагоцитоз |
| FcεRII (CD23 ) | IgE | Низкий (Kd> 10−7 M) | В-клетки Эозинофилы Клетки Лангерганса | Возможная молекула адгезии Транспорт IgE через эпителий кишечника человека Механизм положительной обратной связи для усиления аллергической сенсибилизации (В-клетки) |
| FcαRI (CD89 ) | IgA | Низкий (Kd> 10−6 M) | Моноциты Макрофаги Нейтрофилов Эозинофилы | Фагоцитоз Индукция уничтожения микробов |
| Fcα / мкР | IgA и IgM | Высокий для IgM, средний для IgA | В-клетки Мезангиальные клетки Макрофаги | Эндоцитоз Индукция уничтожения микробов |
| FcRn | IgG | Моноциты Макрофаги Дендритные клетки Эпителиальные клетки Эндотелиальные клетки Гепатоциты | Передает IgG от матери к плоду через плаценту Передает IgG от матери к ребенку с молоком Защищает IgG от деградации |
Функции
Рецепторы Fc обнаружены в ряде клеток иммунной системы, включая фагоциты подобно макрофаги и моноциты, гранулоциты подобно нейтрофилы и эозинофилы, и лимфоциты из врожденная иммунная система (естественные клетки-киллеры ) или же адаптивная иммунная система (например., В-клетки ).[17][18][19]Они позволяют этим клеткам связываться с антителами, которые прикреплены к поверхности микробов или инфицированных микробами клеток, помогая этим клеткам идентифицировать и устранять микробный патогены. Рецепторы Fc связывают антитела на своих Fc регион (или хвост), взаимодействие, которое активирует клетку, обладающую рецептором Fc.[20] Активация фагоцитов - наиболее частая функция, приписываемая рецепторам Fc. Например, макрофаги начинают глотать и убить IgG -покрытый возбудитель к фагоцитоз после включения их рецепторов Fcγ.[21] Другой процесс с участием Fc-рецепторов называется антителозависимая клеточно-опосредованная цитотоксичность (ADCC). Во время ADCC рецепторы FcγRIII на поверхности естественных киллеров (NK) стимулируют NK-клетки к высвобождению цитотоксических молекул из их гранулы убить клетки-мишени, покрытые антителами.[22]FcεRI выполняет другую функцию. FcεRI представляет собой рецептор Fc на гранулоциты, который участвует в аллергические реакции и защита от паразитарных инфекции. Когда присутствует соответствующий аллергический антиген или паразит, перекрестное сшивание не менее двух IgE Молекулы и их рецепторы Fc на поверхности гранулоцита заставляют клетку быстро высвобождать предварительно сформированные медиаторы из своих гранул.[3]
Сигнальные механизмы - гамма-рецепторы Fc
Активация
Гамма-рецепторы Fc относятся к группе Некаталитические тирозин-фосфорилированные рецепторы которые имеют сходный сигнальный путь, включающий фосфорилирование остатков тирозина.[23] Рецепторы генерируют сигналы внутри своих клеток посредством важного мотива активации, известного как Мотив активации иммунорецептора на основе тирозина (ITAM).[24] ITAM - это определенная последовательность аминокислоты (YXXL), встречающиеся дважды подряд в внутриклеточный хвост рецептора. Когда фосфат группы добавляются в тирозин (Y) остаток ITAM, закрепленный на мембране ферменты из Семейство киназ Src, а сигнальный каскад генерируется внутри ячейки. Этот фосфорилирование реакция обычно следует за взаимодействием рецептора Fc с его лиганд. ITAM присутствует во внутриклеточном хвосте FcγRIIA, и его фосфорилирование вызывает фагоцитоз в макрофагах. FcγRI и FcγRIIIA не имеют ITAM, но могут передавать активирующий сигнал своим фагоцитам, взаимодействуя с другим белком, который имеет. Этот адаптерный белок называется субъединицей Fcγ и, как и FcγRIIA, содержит две последовательности YXXL, которые характерны для ITAM.
Торможение
Присутствия только одного мотива YXXL недостаточно для активации клеток, и он представляет собой мотив (I / VXXYXXL), известный как иммунорецепторный ингибитор на основе тирозина (ITIM). FcγRIIB1 и FcγRIIB2 имеют последовательность ITIM и являются ингибирующими рецепторами Fc; они не вызывают фагоцитоз. Ингибирующее действие этих рецепторов контролируется ферментами, удаляющими фосфатные группы из остатков тирозина; то фосфатазы ШП-1 и КОРАБЛЬ-1 подавляют передачу сигналов рецепторами Fcγ.[25] Связывание лиганда с FcγRIIB приводит к фосфорилированию тирозина мотива ITAM. Эта модификация генерирует сайт связывания для фосфатазы, домен узнавания SH2. Отмена передачи сигналов активации ITAM вызывается ингибированием протеинтирозинкиназ семейства Src и гидролизом мембраны PIP3, прерывая дальнейшую передачу сигналов ниже по течению активирующими рецепторами, такими как активация FcγRs, TCR, BCR и рецепторов цитокинов (например, c-Kit ).[26]
Отрицательная передача сигналов с помощью FcγRIIB в основном важна для регуляции активированных В-клеток. Положительная передача сигналов В-клеток инициируется связыванием чужеродного антигена с поверхностным иммуноглобулином. Выделяется одно и то же антиген-специфическое антитело, которое может подавлять обратную связь или способствовать передаче негативных сигналов. Эта негативная передача сигналов обеспечивается FcγRIIB:[27] Эксперименты с использованием мутантов с делецией В-клеток и доминантно-отрицательных ферментов твердо установили важную роль SH2-домен-содержащей инозитол-5-фосфатазы (SHIP) в передаче отрицательных сигналов. Отрицательная передача сигналов через SHIP, по-видимому, препятствует Рас путь через конкуренцию домена SH2 с Grb2 и Shc и может включать потребление внутриклеточных липидных медиаторов, которые действуют как активаторы аллостерических ферментов или способствуют проникновению внеклеточного Ca2 +.[28]
Сотовая активация
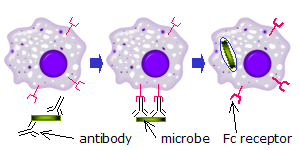
На фагоцитах
Когда IgG молекулы, специфичные для определенного антигена или поверхностного компонента, связываются с патогеном своими Fab регион (фрагмент антигенсвязывающей области), их Fc-области направлены наружу, в непосредственной близости от фагоциты. Фагоциты связывают эти области Fc своими рецепторами Fc.[21] Между рецептором и антителом образуется множество низкоаффинных взаимодействий, которые работают вместе, чтобы прочно связывать микроб, покрытый антителами. Низкая индивидуальная аффинность не позволяет рецепторам Fc связывать антитела в отсутствие антигена и, следовательно, снижает вероятность активации иммунных клеток в отсутствие инфекции. Это также предотвращает агглютинация (свертывание) фагоциты антителом при отсутствии антигена. После связывания патогена взаимодействия между Fc-областью антитела и Fc-рецепторами фагоцита приводят к инициации фагоцитоз. Патоген поглощается фагоцитом в результате активного процесса, включающего связывание и высвобождение комплекса Fc-область / Fc-рецептор, пока не произойдет клеточная мембрана фагоцита полностью вмещает возбудителя.[29]
На NK-клетках
Рецептор Fc на NK-клетках распознает IgG, который связан с поверхностью инфицированной патогеном клетки-мишени и называется CD16 или FcγRIII.[30] Активация FcγRIII IgG вызывает высвобождение цитокины Такие как IFN-γ которые сигнализируют другим иммунным клеткам, и цитотоксические медиаторы, такие как перфорин и гранзим которые проникают в клетку-мишень и способствуют гибели клеток, вызывая апоптоз. Этот процесс известен как антителозависимая клеточно-опосредованная цитотоксичность (ADCC). FcγRIII на NK-клетках также может связываться с мономерным IgG (то есть с IgG, который не связан с антигеном). Когда это происходит, рецептор Fc подавляет активность NK-клетки.[31]
На тучных клетках

IgE антитела связываются с антигены из аллергены. Эти связанные с аллергеном молекулы IgE взаимодействуют с рецепторами Fcε на поверхности тучные клетки. Активация тучных клеток после включения FcεRI приводит к процессу, называемому дегрануляция, посредством чего тучная клетка высвобождает предварительно сформированные молекулы из своего цитоплазматический гранулы; это смесь соединений, включая гистамин, протеогликаны, и сериновые протеазы.[32] Активированные тучные клетки также синтезируют и секретируют липид -производные медиаторы (такие как простагландины, лейкотриены, и фактор активации тромбоцитов ) и цитокины (такие как интерлейкин 1, интерлейкин 3, интерлейкин 4, интерлейкин 5, интерлейкин 6, интерлейкин 13, фактор некроза опухоли альфа, GM-CSF, и несколько хемокины.[33][34] Эти посредники способствуют воспаление за счет привлечения других лейкоциты.
Об эозинофилах
Крупные паразиты, такие как гельминт (червь) Schistosoma mansoni слишком велики для поглощения фагоцитами. У них также есть внешняя структура, называемая покров устойчивый к воздействию веществ, выделяемых макрофагами и тучными клетками. Однако эти паразиты могут покрыться IgE и распознаваться FcεRI на поверхности эозинофилы. Активированные эозинофилы высвобождают предварительно сформированные медиаторы, такие как основной основной белок, и ферменты Такие как пероксидаза, против которого гельминты не устойчивы.[35][36] Взаимодействие FcεRI рецептор с Fc-частью связанного с гельминтом IgE заставляет эозинофил высвобождать эти молекулы по механизму, аналогичному механизму NK-клетки во время ADCC.[37]
На Т-лимфоцитах
CD4 + Т-клетки (зрелые тчас клетки ) помогают В-клеткам, вырабатывающим антитела. При патологии заболевания наблюдается несколько подмножеств активированных эффекторных CD4 + Т-клеток. Более ранние исследования, обобщенные Сандерсом и Линчем в 1993 г., предполагали критическую роль FcR в иммунных ответах, опосредованных CD4 + Т-клетками, и предполагали образование совместного сигнального комплекса между FcR и TCR на поверхности клетки.[38][39][40][41] Chauhan и соавторы сообщили о совместной локализации меченых ICs с комплексом CD3 на активированной поверхности CD4 + Т-клеток, что, таким образом, предполагает сосуществование FcR вместе с комплексом TCR.[42] Оба этих рецептора образуют апикальную структуру на мембране активированных CD4 + Т-клеток, что свидетельствует о боковом движении этих рецепторов.[43] Совместная миграция FcR с TCR и комплексом BCR наблюдается на поверхности клеток, и цитоконъюгаты T: B-клеток демонстрируют это сосуществование в точке контакта.[44] Предыдущий обзор показал, что экспрессия FcR на CD4 + Т-клетках остается открытым вопросом.[45] Это установило текущую парадигму, согласно которой Т-клетки не экспрессируют FcR, и эти результаты никогда не подвергались сомнению и экспериментально не проверялись.[46] Чаухан и его коллеги показали связывание иммунных комплексов (IC), лиганда FcR, с активированными CD4 + Т-клетками.[46] Экспрессия CD16a индуцируется в активированных наивных CD4 + Т-клетках человека, которые экспрессируют CD25, CD69 и CD98, и лигирование с IC приводит к образованию эффекторных клеток памяти.[47] Передача сигналов CD16a опосредуется фосфорилированием Syk (pSyk).[47][48][49]
В настоящее время исследование предполагает индуцированную экспрессию CD32a при активации CD4 + Т-клеток человека, аналогичную CD16a.[48][50] Экспрессия CD32a на CD4 + Т-клетках также была подтверждена тремя независимыми исследованиями исследователей ВИЧ-1. Экспрессия CD16a и CD32a в подмножестве активированных CD4 + Т-клеток теперь подтверждена.[48][50] FcR на поверхности клетки при связывании с IC, состоящими из нуклеиновых кислот, запускают продукцию цитокинов и активируют пути восприятия нуклеиновых кислот. FcR присутствуют как на поверхности клетки, так и в цитозоле. Передача сигнала CD16a усиливает экспрессию толл-подобных рецепторов, чувствительных к нуклеиновым кислотам, и перемещает их на поверхность клетки.[47][51] CD16a - это новый костимулирующий сигнал для CD4 + Т-клеток человека, который успешно заменяет потребность в CD28 во время аутоиммунитета.[52] На аутоиммунном фоне CD4 + Т-клетки обходят требование косигнализации CD28, чтобы стать полностью активированными.[52] Кроме того, блокада косигнализации CD28 не ингибирует развитие клеток TFH, ключевой субпопуляции для генерации аутоантител, продуцирующих аутореактивные плазматические В-клетки.[53] Для иммунного гомеостаза необходим баланс между костимулирующими и тормозными сигналами. Чрезмерная костимуляция и / или недостаточное совместное ингибирование приводит к нарушению толерантности и аутоиммунитету. Костимуляция, опосредованная CD16a, обеспечивает положительный сигнал в активированных CD4 + Т-клетках, а не в покоящихся клетках, в которых отсутствует экспрессия FcγR.[48]
Смотрите также
Рекомендации
- ^ Андерсон Р. (2003). «Манипуляции с макромолекулами клеточной поверхности флавивирусами». Достижения в вирусных исследованиях. 59: 229–74. Дои:10.1016 / S0065-3527 (03) 59007-8. ISBN 9780120398591. ЧВК 7252169. PMID 14696331.
- ^ Оуэн Дж, Пунт Дж, Стрэнфорд С., Джонс П. (2009). Иммунология (7-е изд.). Нью-Йорк: W.H. Фримен и компания. п. 423. ISBN 978-14641-3784-6.
- ^ а б c d Фридман WH (сентябрь 1991 г.). «Fc рецепторы и факторы связывания иммуноглобулина». Журнал FASEB. 5 (12): 2684–90. Дои:10.1096 / fasebj.5.12.1916092. PMID 1916092.
- ^ Индик З.К., Парк Дж. Г., Хантер С., Шрайбер А. Д. (декабрь 1995 г.). «Молекулярное рассечение фагоцитоза, опосредованного рецептором Fc гамма». Кровь. 86 (12): 4389–99. Дои:10.1182 / blood.V86.12.4389.bloodjournal86124389. PMID 8541526.
- ^ Харрисон П. Т., Дэвис В., Норман Дж. К., Хокадей А. Р., Аллен Дж. М. (сентябрь 1994 г.). «Связывание мономерного иммуноглобулина G запускает Fc-гамма-RI-опосредованный эндоцитоз». Журнал биологической химии. 269 (39): 24396–402. PMID 7929100.
- ^ а б c d Маверакис Э., Ким К., Шимода М., Гершвин М.Э., Патель Ф., Уилкен Р., Райчаудхури С., Рухак Л. Р., Лебрилла CB (февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор». Журнал аутоиммунитета. 57 (6): 1–13. Дои:10.1016 / j.jaut.2014.12.002. ЧВК 4340844. PMID 25578468.
- ^ Чжу X, Мэн Джи, Дикинсон Б.Л., Ли Х, Мизогучи Э, Мяо Л., Ван И, Роберт К., Ву Б., Смит П.Д., Ленсер В.И., Блумберг Р.С. (март 2001 г.). «Связанный с MHC класс I неонатальный рецептор Fc для IgG функционально экспрессируется в моноцитах, кишечных макрофагах и дендритных клетках». Журнал иммунологии. 166 (5): 3266–76. Дои:10.4049 / jimmunol.166.5.3266. ЧВК 2827247. PMID 11207281.
- ^ Фиран М., Боудон Р., Раду С., Обер Р.Дж., Экен Д., Антох Ф., Гети В., Уорд Е.С. (август 2001 г.). «Рецептор, связанный с MHC класса I, FcRn, играет важную роль в материнско-фетальном переносе гамма-глобулина у людей». Международная иммунология. 13 (8): 993–1002. Дои:10.1093 / intimm / 13.8.993. PMID 11470769.
- ^ Simister NE, Jacobowitz Israel E, Ahouse JC, Story CM (май 1997 г.). «Новые функции рецептора Fc, относящегося к MHC класса I, FcRn». Сделки Биохимического Общества. 25 (2): 481–6. Дои:10.1042 / bst0250481. PMID 9191140.
- ^ а б c Оттен М.А., ван Эгмонд М. (март 2004 г.). «Рецептор Fc для IgA (FcalphaRI, CD89)». Письма иммунологии. 92 (1–2): 23–31. Дои:10.1016 / j.imlet.2003.11.018. PMID 15081523.
- ^ Shibuya A, Honda S (декабрь 2006 г.). «Молекулярные и функциональные характеристики Fcalpha / muR, нового рецептора Fc для IgM и IgA». Семинары Springer по иммунопатологии. 28 (4): 377–82. Дои:10.1007 / s00281-006-0050-3. PMID 17061088.
- ^ Чо Й, Усуи К., Хонда С., Тахара-Ханаока С., Сибуя К., Сибуя А. (июнь 2006 г.). «Молекулярные характеристики связывания IgA и IgM Fc с Fcalpha / muR». Сообщения о биохимических и биофизических исследованиях. 345 (1): 474–8. Дои:10.1016 / j.bbrc.2006.04.084. PMID 16681999.
- ^ Очиай К., Ван Б., Ригер А., Килгус О., Маурер Д., Фёдингер Д., Кинет Дж. П., Стингл Г., Томиока Х (1994). «Обзор Fc epsilon RI на эпидермальных клетках Лангерганса человека». Международный архив аллергии и иммунологии. 104 Дополнение 1 (1): 63–4. Дои:10.1159/000236756. PMID 8156009.
- ^ Пруссин С., Меткалф Д.Д. (февраль 2006 г.). «5. IgE, тучные клетки, базофилы и эозинофилы». Журнал аллергии и клинической иммунологии. 117 (2 доп. Мини-праймера): S450-6. Дои:10.1016 / j.jaci.2005.11.016. PMID 16455345.
- ^ фон Бубнофф Д., Новак Н., Крафт С., Бибер Т. (март 2003 г.). «Центральная роль FcepsilonRI при аллергии». Клиническая и экспериментальная дерматология. 28 (2): 184–7. Дои:10.1046 / j.1365-2230.2003.01209.x. PMID 12653710.
- ^ Кикутани Х., Ёкота А., Учибаяси Н., Юкава К., Танака Т., Сугияма К., Барсумиан Э. Л., Суэмура М., Кишимото Т. (1989). «Структура и функция рецептора Fc-эпсилон II (Fc-эпсилон RII / CD23): точка контакта между эффекторной фазой аллергии и дифференцировкой В-клеток». Симпозиум Фонда Ciba. Симпозиумы Фонда Новартис. 147: 23–31, обсуждение 31–5. Дои:10.1002 / 9780470513866.ch3. ISBN 9780470513866. PMID 2695308.
- ^ Сельварадж П., Фифадара Н., Нагараджан С., Чимино А., Ван Г. (2004). «Функциональная регуляция человеческих нейтрофильных Fc-рецепторов гамма». Иммунологические исследования. 29 (1–3): 219–30. Дои:10.1385 / ИК: 29: 1-3: 219. PMID 15181284.
- ^ Сулика А., Чемберс WH, Мансиула М., Метес Д., Кори С., Рабинович Х., Уайтсайд Т.Л., Герберман РБ (1995). «Дивергентные пути передачи сигнала и эффекты на функции естественных клеток-киллеров, индуцированные взаимодействием рецепторов Fc с физиологическими лигандами или антирецепторными антителами». Естественный иммунитет. 14 (3): 123–33. PMID 8832896.
- ^ Сарфати М., Фурнье С., Ву С.Й., Делеспес Дж. (1992). «Экспрессия, регуляция и функция человеческого антигена Fc epsilon RII (CD23)». Иммунологические исследования. 11 (3–4): 260–72. Дои:10.1007 / BF02919132. PMID 1287120.
- ^ Рагхаван М., Бьоркман П.Дж. (1996). «Рецепторы Fc и их взаимодействие с иммуноглобулинами» (PDF). Ежегодный обзор клеточной биологии и биологии развития. 12: 181–220. Дои:10.1146 / annurev.cellbio.12.1.181. PMID 8970726.
- ^ а б Суонсон Дж. А., Хоппе А. Д. (декабрь 2004 г.). «Координация передачи сигналов во время фагоцитоза, опосредованного рецептором Fc». Журнал биологии лейкоцитов. 76 (6): 1093–103. Дои:10.1189 / jlb.0804439. HDL:2027.42/141562. PMID 15466916.
- ^ Sun PD (2003). «Структура и функция рецепторов естественных клеток-киллеров». Иммунологические исследования. 27 (2–3): 539–48. Дои:10.1385 / ИК: 27: 2-3: 539. PMID 12857997.
- ^ Душек О., Гойетт Дж., Ван дер Мерве ПА (ноябрь 2012 г.). «Некаталитические тирозин-фосфорилированные рецепторы». Иммунологические обзоры. 250 (1): 258–276. Дои:10.1111 / imr.12008. PMID 23046135.
- ^ Cambier JC (февраль 1995 г.). «Новая номенклатура для мотива Reth (или ARH1 / TAM / ARAM / YXXL)». Иммунология сегодня. 16 (2): 110. Дои:10.1016/0167-5699(95)80105-7. PMID 7888063.
- ^ Хуанг З.Й., Хантер С., Ким М.К., Индик З.К., Шрайбер А.Д. (июнь 2003 г.). «Влияние фосфатаз SHP-1 и SHIP-1 на передачу сигналов ITIM- и ITAM-содержащими рецепторами Fcgamma FcgammaRIIB и FcgammaRIIA». Журнал биологии лейкоцитов. 73 (6): 823–9. Дои:10.1189 / jlb.0902454. PMID 12773515.
- ^ Cambier JC (июнь 1997 г.). "Тормозных рецепторов предостаточно?". Труды Национальной академии наук Соединенных Штатов Америки. 94 (12): 5993–5. Bibcode:1997PNAS ... 94.5993C. Дои:10.1073 / пнас.94.12.5993. ЧВК 33673. PMID 9177155.
- ^ Такай Т., Оно М., Хикида М., Омори Х., Раветч СП (январь 1996 г.). «Усиленные гуморальные и анафилактические ответы у мышей с дефицитом Fc гамма RII». Природа. 379 (6563): 346–9. Bibcode:1996Натура 379..346Т. Дои:10.1038 / 379346a0. PMID 8552190.
- ^ Coggeshall KM (июнь 1998 г.). «Ингибирующая передача сигналов с помощью B-клеточного Fc гамма RIIb». Текущее мнение в иммунологии. 10 (3): 306–12. Дои:10.1016 / s0952-7915 (98) 80169-6. PMID 9638367.
- ^ Джоши Т., Бутчар Дж. П., Тридандапани С. (октябрь 2006 г.). «Передача сигналов рецептора Fcgamma в фагоцитах». Международный журнал гематологии. 84 (3): 210–6. Дои:10.1532 / IJH97.06140. PMID 17050193.
- ^ Тринкьери Г., Валианте Н. (1993). «Рецепторы для Fc-фрагмента IgG на естественных клетках-киллерах». Естественный иммунитет. 12 (4–5): 218–34. PMID 8257828.
- ^ Сулика А., Галатюк С., Манчуля М., Банку А.С., ДеЛео А., Уайтсайд Т.Л., Херберман РБ (апрель 1993 г.). «Регулирование естественной цитотоксичности человека с помощью IgG. IV. Связь между связыванием мономерного IgG с рецепторами Fc на больших гранулярных лимфоцитах и ингибированием активности естественных киллеров (NK)». Клеточная иммунология. 147 (2): 397–410. Дои:10.1006 / cimm.1993.1079. PMID 8453679.
- ^ Ямасаки С., Сайто Т. (2005). «Регулирование активации тучных клеток через FcepsilonRI». Химическая иммунология и аллергия. 87: 22–31. Дои:10.1159/000087568. ISBN 3-8055-7948-9. PMID 16107760.
- ^ Wakahara S, Fujii Y, Nakao T., Tsuritani K, Hara T., Saito H, Ra C. (ноябрь 2001 г.). «Профили экспрессии генов для Fc-эпсилон RI, цитокинов и хемокинов при активации Fc-эпсилон RI в культивируемых тучных клетках человека, полученных из периферической крови». Цитокин. 16 (4): 143–52. Дои:10.1006 / cyto.2001.0958. PMID 11792124.
- ^ Меткалф Д.Д., Барам Д., Мекори Ю.А. (октябрь 1997 г.). "Тучные клетки". Физиологические обзоры. 77 (4): 1033–79. Дои:10.1152 / Physrev.1997.77.4.1033. PMID 9354811.
- ^ Дэвид Дж. Р., Баттерворт А. Е., Вадас Массачусетс (сентябрь 1980 г.). «Механизм взаимодействия, опосредующего убийство Schistosoma mansoni человеческими эозинофилами». Американский журнал тропической медицины и гигиены. 29 (5): 842–8. Дои:10.4269 / ajtmh.1980.29.842. PMID 7435788.
- ^ Капрон М., Суси Гунни А., Морита М., Чыонг М.Дж., Прин Л., Кинет Дж. П., Капрон А. (1995). «Эозинофилы: от рецепторов иммуноглобулинов Е с низким и высоким сродством». Аллергия. 50 (25 Дополнение): 20–3. Дои:10.1111 / j.1398-9995.1995.tb04270.x. PMID 7677229.
- ^ Gounni AS, Lamkhioued B, Ochiai K, Tanaka Y, Delaporte E, Capron A, Kinet JP, Capron M (январь 1994). «Рецептор IgE с высоким сродством к эозинофилам участвует в защите от паразитов». Природа. 367 (6459): 183–6. Bibcode:1994Натура.367..183С. Дои:10.1038 / 367183a0. PMID 8114916.
- ^ Пихлер WJ, Lum L, Broder S (1978). «Fc-рецепторы на Т-лимфоцитах человека. I. Переход Tgamma в Tmu-клетки». J Immunol. 121 (4): 1540–1548. PMID 308968.
- ^ Сандор М., Линч Р.Г. (май 1993 г.). «Лимфоцитарные рецепторы Fc: частный случай Т-клеток». Иммунол. Сегодня. 14 (5): 227–31. Дои:10.1016 / 0167-5699 (93) 90168-К. PMID 8517922.
- ^ Энгельгардт В., Мацке Дж., Шмидт Р.Э. (1995). «Зависимая от активации экспрессия низкоаффинных рецепторов IgG Fc гамма RII (CD32) и Fc гамма RIII (CD16) в субпопуляциях Т-лимфоцитов человека». Иммунобиология. 192 (5): 297–320. Дои:10.1016 / s0171-2985 (11) 80172-5. PMID 7649565.
- ^ Моретта Л., Уэбб С.Р., Гросси К.Э., Лидьярд П.М., Купер М.Д. (1977). «Функциональный анализ двух субпопуляций человеческих Т-клеток: помощь и подавление В-клеточных ответов Т-клетками, несущими рецепторы IgM или IgG». J Exp Med. 146 (1): 184–200. Дои:10.1084 / jem.146.1.184. ЧВК 2180738. PMID 301544.
- ^ Чаухан А.К., Мур Т.Л. (2011). «Активация Т-клеток терминальным комплексом комплемента и иммунных комплексов». Журнал биологической химии. 286 (44): 38627–38637. Дои:10.1074 / jbc.M111.266809. ЧВК 3207419. PMID 21900254.
- ^ Чаухан А.К., Мур Т.Л. (2011). «Активация Т-клеток терминальным комплексом комплемента и иммунных комплексов». Журнал биологической химии. 286 (44): 38627–38637. Дои:10.1074 / jbc.M111.266809. ЧВК 3207419. PMID 21900254.
- ^ Сандор М., Линч Р.Г. (1993). «Лимфоцитарные рецепторы Fc: частный случай Т-клеток». Иммунология сегодня. 14 (5): 227–231. Дои:10.1016 / 0167-5699 (93) 90168-К. PMID 8517922.
- ^ Nimmerjahn F, Ravetch JV (январь 2008 г.). «Рецепторы Fcgamma как регуляторы иммунных ответов». Nat. Преп. Иммунол. 8 (1): 34–47. Дои:10.1038 / nri2206. PMID 18064051.
- ^ а б Брунс П., Йонссон Ф. (ноябрь 2015 г.). «Эффекторные функции FcR мыши и человека». Иммунол. Rev. 268 (1): 25–51. Дои:10.1111 / imr.12350. PMID 26497511.
- ^ а б c Чаухан А.К., Мур Т.Л., Би Й., Чен С. (январь 2016 г.). «Ко-сигнал FcγRIIIa-Syk модулирует CD4 + Т-клеточный ответ и повышает экспрессию Toll-подобного рецептора (TLR)». J. Biol. Chem. 291 (3): 1368–86. Дои:10.1074 / jbc.M115.684795. ЧВК 4714221. PMID 26582197.
- ^ а б c d Чаухан А.К., Чен С., Мур Т.Л., ДиПаоло Р.Дж. (февраль 2015 г.). «Индуцированная экспрессия FcγRIIIa (CD16a) на CD4 + Т-клетках запускает генерацию IFN-γhigh подмножества». J. Biol. Chem. 290 (8): 5127–40. Дои:10.1074 / jbc.M114.599266. ЧВК 4335247. PMID 25556651.
- ^ Чаухан А.К., Мур Т.Л. (2012). «Иммунные комплексы и поздние белки комплемента запускают активацию тирозинкиназы Syk в CD4 (+) Т-клетках человека». Клин Эксп Иммунол. 167 (2): 235–245. Дои:10.1111 / j.1365-2249.2011.04505.x. ЧВК 3278689. PMID 22235999.
- ^ а б Член парламента Ольгадо, Сананес I, Райден С., Геффнер-младший, Аррувито Л. (2018). «Лигирование CD32 способствует активации CD4 + Т-клеток». Фронт Иммунол. 9: 2814. Дои:10.3389 / fimmu.2018.02814. ЧВК 6284025. PMID 30555482.
- ^ Чаухан АК (2017). «Передача сигналов FcgammaRIIIa модулирует эндосомные ответы TLR в CD4 + Т-клетках человека». J Immunol. 198 (12): 4596–4606. Дои:10.4049 / jimmunol.1601954. ЧВК 5505339. PMID 28500073.
- ^ а б Бур-Джордан Х, Эзенстен Дж. Х., Мартинес-Льорделла М., Пенаранда С., Штумпф М., Блюстоун Дж. А. (2011). «Внутренний и внешний контроль толерантности периферических Т-клеток костимулирующими молекулами семейства CD28 / B7». Иммунол Рев. 241 (1): 180–205. Дои:10.1111 / j.1600-065X.2011.01011.x. ЧВК 3077803. PMID 21488898.
- ^ Вебер Дж. П., Фурманн Ф., Файст Р.К., Лахманн А., Аль Баз М.С., Генц Л.Дж., Ву Ван Д., Маги Х.В., Хафтманн С., Ридель Р., Грун Дж. Р., Шух В., Крочек Р.А., Радбрух А., Машреги М.Ф., Хутлофф А. ( 2015). «ICOS поддерживает фенотип Т-фолликулярных хелперных клеток путем подавления Kruppel-подобного фактора 2». Журнал экспериментальной медицины. 212 (2): 217–233. Дои:10.1084 / jem.20141432. ЧВК 4322049. PMID 25646266.
дальнейшее чтение
- Laneway CA, Трэверс П., Уолдпорт М., Шломчик М.Дж. (2001). «Глава 9. Гуморальный иммунный ответ». Иммунобиология: иммунная система в здоровье и болезнях (5-е изд.). Нью-Йорк: Гарленд. ISBN 978-0-8153-3642-6.
- Аббас А.К., Лихтман А.Х., Пиллай С. (2012). «Глава 12: Эффекторные механизмы гуморального иммунитета». Клеточная и молекулярная иммунология (7-е изд.). Филадельфия: Эльзевьер / Сондерс. ISBN 978-1-4377-1528-6.
- Гербер Дж. С., Моссер Д. М. (февраль 2001 г.). «Стимулирующие и тормозящие сигналы, исходящие от рецепторов макрофагов Fcgamma». Микробы и инфекции. 3 (2): 131–9. Дои:10.1016 / с1286-4579 (00) 01360-5. PMID 11251299.
- Маверакис Э., Ким К., Шимода М., Гершвин М.Э., Патель Ф., Уилкен Р., Райчаудхури С., Рухак Л. Р., Лебрилла CB (февраль 2015 г.). «Гликаны в иммунной системе и теория аутоиммунитета с измененными гликанами: критический обзор». Журнал аутоиммунитета. 57: 1–13. Дои:10.1016 / j.jaut.2014.12.002. ЧВК 4340844. PMID 25578468.
внешняя ссылка
- Fc + рецептор в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)