Формирование парка динозавров - Википедия - Dinosaur Park Formation
| Формирование парка динозавров Стратиграфический диапазон: Кампанский, 76.9–75.8 Ма | |
|---|---|
 Формирование парка динозавров обнажено вдоль Ред Дир Ривер в Провинциальный парк динозавров, юго-восточная Альберта, Канада. | |
| Тип | Геологическое образование |
| Единица | Belly River Group |
| Лежит в основе | Формация Медвежьей Лапы |
| Перекрывает | Формация Олдмана |
| Литология | |
| Начальный | Песчаник (ниже) Аргиллит и алевролит (верхний) |
| Другой | Бентонит и каменный уголь |
| Место расположения | |
| Координаты | 49 ° 12′N 110 ° 24'з.д. / 49,2 ° с.ш.110,4 ° з.Координаты: 49 ° 12′N 110 ° 24'з.д. / 49,2 ° с.ш.110,4 ° з. |
| Приблизительные палеокоординаты | 56 ° 24′N 75 ° 48'з.д. / 56,4 ° с. Ш. 75,8 ° з. |
| Область, край | |
| Страна | |
| Степень | Западно-канадский осадочный бассейн |
| Тип раздела | |
| Названный для | Провинциальный парк динозавров |
| Названный | Эберт, Д.А. и Хэмблин, А.[1][2] |
| Год определен | 1993 |
 Формирование парка динозавров (Канада)  Формация парка динозавров (Альберта) | |
В Формирование парка динозавров является самым верхним членом Belly River Group (также известный как Джудит Ривер Групп ), крупная геологическая единица на юге Альберта. Он был депонирован во время Кампанский этап Поздний мел, примерно между 76,9 и 75,8 миллионами лет назад.[3] Он был депонирован в аллювиальный и прибрежная равнина среды, и он ограничен неморскими Формация Олдмана под ним и морской Формация Медвежьей Лапы над ним.[4]
Формация парка динозавров содержит плотные скопления динозавр скелеты, как сочлененные, так и разрозненные, часто встречаются с сохранившимися остатками мягких тканей. Остатки других животных, таких как рыбы, черепахи, и крокодилы, как и остатки растений, также в изобилии.[5] Образованию присвоено имя Провинциальный парк динозавров, а ЮНЕСКО Объект всемирного наследия где пласт хорошо обнажается в бесплодные земли это фланг Ред Дир Ривер.[2]
Геологическая обстановка

Формирование парка динозавров состоит из отложений, образовавшихся в результате эрозии гор на западе. Он был отложен на аллювиальной прибрежной равнине речными системами, которые текли на восток и юго-восток в море Медвежьей Лапы, большое внутреннее море, которое было частью Западный внутренний морской путь. Это море постепенно затопляло прилегающую прибрежную равнину, оседая на море. сланцы из Формация Медвежьей Лапы на вершине формирования парка динозавров.[4]
Формация парка динозавров имеет толщину около 70 метров (230 футов) в парке динозавров. Нижняя часть формации закладывалась в речной канал среды и состоит в основном из мелкозернистых и среднезернистых, перекрестный песчаники. Верхняя часть, которая была отложена в надбанк и пойма среды, состоящие в основном от массивных до ламинированных, богатых органикой аргиллиты с обильными следами корней и тонкими слоями бентонит. Угольная зона Летбриджа, состоящая из нескольких пластов низший ранг каменный уголь с прослоями аргиллитов и алевролиты, отмечает вершину формации.[4]
Отложения формации «Парк динозавров» аналогичны отложениям нижележащих слоев. Формация Олдмана и они изначально были включены в эту формацию. Эти два образования разделяет региональный несоответствие, однако, и отличаются петрографический и седиментологический различия. Кроме того, шарнирно-сочлененные костные останки и костные кровати редки в формации Олдмана, но многочисленны в формации парка динозавров.[2][4]
Биостратиграфия
Формация парка динозавров может быть разделена как минимум на две отдельные фауны. Нижняя часть свиты характеризуется обилием Коритозавр и Центрозавр. Эта группа видов замещается выше в формации другой орнитисхофауной, характеризующейся наличием Ламбеозавр и Стиракозавр. Появление нескольких новых, редких видов орнитисхиев на самой вершине формации может указывать на то, что третья отдельная фауна заменила вторую во время перехода к более молодым, не относящимся к Парку динозавров отложениям, в то время как внутреннее море выходит на сушу. , но здесь осталось меньше. Безымянный пахиринозавр, Вагацератопс irvinensis, и Ламбеозавр магникристатус может быть более обычным в этой третьей фауне.[6][7]
Приведенная ниже временная шкала следует за синтезом, представленным Фаулером (2016).[3] с дополнительной информацией от Беседка и другие. 2009,[8] Эванс и другие. 2009, и Пенкальски, 2013.[9] Зоны скопления крупных травоядных животных (MAZ) соответствуют данным, представленным Маллоном. и другие., 2012.[10]

Амфибии
В свите обнаружены останки следующих амфибий:[11]
Albanerpetontidae (вымерший, саламандра -подобные амфибии)
- Габрозавр продилатус
- Лиссерпетон
- Опистотритон кайи
- Скаферпетон тектум
- безымянный хвостатан
- 2 неопределенных хвостатых
- 2 безымянных салиентана
- Tyrrellbatrachus brinkmani[12]
- Hensonbatrachus kermiti[13]
Динозавры
В свите обнаружены останки следующих динозавров:[8][14]
Орнитисхианы
В свите обнаружены остатки следующих орнитисхий:[15]
Анкилозавры
| Анкилозавры из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Анодонтозавр | А. inceptus | An анкилозаврин анкилозаврид | ||||
| Диоплозавр | D. acutosquameus | Нижний, 76,5 млн. Назад[8] | Анкилозаврин анкилозаврид |  | ||
| Эдмонтония | E. rugosidens | Нижний, 76.5-75.9 млн. Назад[8] | А нодозаврин нодозаврид |  | ||
| Евоплоцефал | E. пачки | От нижнего до среднего, ~ 76,4-75,6 млн лет назад[9] | Анкилозаврин анкилозаврид |  | ||
| Паноплозавр | P. mirus | Middle, 75.6Ma назад[8] | «Частичный скелет с полным черепом, остеодермами, дополнительными изолированными зубами, посткраниальными элементами, остеодермами».[16] | Нодозаврид нодозаврид |  | |
| Платипельта | P. coombsi | Нижняя, 77,5-76,5 МА[17] | Анкилозаврин анкилозаврид |  | ||
| Сколозавр | С. катлери | Ниже, 76,5 млн. Лет назад или больше[9] | Анкилозаврин анкилозаврид, который коротко считали синонимом Евоплоцефал. Возможно, это происходило из верхних слоев нижележащего Формация Олдмана.[18] |  | ||
| S. thronus | Анкилозаврин анкилозаврид | |||||
Цератопсы
Безымянный Пахиринозавр -подобный таксон восстановлен из формации.
| Цератопсы из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Центрозавр | C. apertus | Middle, 76.2-75.5Ma назад[8] | «[Пятнадцать] черепов, несколько скелетов, все взрослые; обильный материал костного ложа с редкими молодыми и половозрелыми».[19] C. nasicornis может быть синонимом. | А центрозаврин цератопсид |  | |
| Хасмозавр | C. belli | Средний, 76-75,5 млн. Назад[8] | «[Двенадцать] черепов, несколько скелетов».[19] | А хазмозаврин цератопсид | 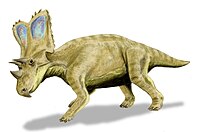 | |
| К. Расселли | Lower, 76.5-76Ма назад[8] | «[Шесть] полных или частичных черепов».[20] | ||||
| Меркурицератопс | M. gemini[21] | Нижний, ~ 77 млн. Назад[21] | "одна апоморфная чешуя"[21] | Хасмозаврин цератопсид |  | |
| Моноклониус | М. lowei | А сомнительный центрозаврин цератопсид. Возможно синоним Центрозавр. | ||||
| Пентацератопс[22] | P. aquilonius[22] | Самый верхний, 74,8 МА[22] | два фрагмента оборки[22] | Сомнительный хазмозаврин цератопсид, который может быть того же вида, что и Spiclypeus shipporum.[23] |  | |
| Спинопс[24] | S. sternbergorum[24] | Нижний, 76,5 мА[24] | «частичная теменная кость, частичная зубная кость, неидентифицируемые фрагменты конечностей, частичный череп и частичный правый чешуйчатый кость».[24] | Центрозаврин цератопсид. На самом деле он может быть из верхней формации Олдмана.[24] |  | |
| Стиракозавр | S. albertensis | Upper, 75.5-75.2Ma назад[8] | «[Два] черепа, [три] скелета, дополнительный материал в костных ложах».[19] | Центрозаврин цератопсид |  | |
| Юнескоцератопс | U. koppelhusae | Неполная нижняя челюсть[25] | А лептоцератопсид Считается, что его длина составляла от одного до двух метров, а вес - менее 91 килограмма. Его зубы были самыми круглыми из всех лептоцератопсид. | |||
| Вагацератопс | V. irvinensis | Upper, 75Мнн назад[8] | «[Три] черепа, скелет без хвоста».[20] | Хасмозавр цератопсид, ранее классифицированный как вид Хасмозавр.[26] |  | |
Орнитоподы
По крайней мере, один неопределенный гипсилофодонт образец был извлечен из пласта.
В обзоре гадрозавров 2001 г. яичная скорлупа и материал для вылупления из формации парка динозавров, Даррен Х. Танке и М. К. Бретт-Сурман пришли к выводу, что гадрозавры гнездились как на древних возвышенностях, так и в низинах среды осадконакопления формации.[27]Горные места гнездования могли быть предпочтительны для менее распространенных гадрозавров, таких как Брахилофозавр или же Паразауролоф. Однако авторам не удалось определить, какие именно факторы повлияли на выбор места гнездования у гадрозавров формации. Они предположили, что поведение, диета, состояние почвы и конкуренция между видами динозавров потенциально влияют на то, где гнездятся гадрозавры.[28]
Сообщалось о субсантиметровых фрагментах яичной скорлупы гадрозавров с текстурой гальки из формации «Парк динозавров». Эта яичная скорлупа похожа на яичную скорлупу гадрозавра. Кули дьявола на юге Альберта а также Два лекарства и Формации реки Джудит в Монтане, США.[29] Яичная скорлупа динозавров, хотя и присутствует, очень редко встречается в формации парка динозавров и встречается только в двух разных местах обнаружения ископаемых.[27] Эти сайты отличаются большим количеством писидий моллюски и других менее распространенных беспозвоночных с панцирем, таких как Unionid моллюски и улитки. Эта ассоциация не случайна, поскольку скорлупа беспозвоночных медленно растворялась бы и выделяла достаточно основного карбоната кальция для защиты яичной скорлупы от встречающихся в природе кислот, которые в противном случае растворили бы их и предотвратили бы окаменение.[29]
В отличие от окаменелостей яичной скорлупы, останки очень молодых гадрозавров встречаются довольно часто. Даррен Танке заметил, что опытный коллекционер может обнаружить несколько особей молодых гадрозавров за один день. Наиболее распространенными останками молодых гадрозавров в формации парка динозавров являются: зубные кости, кости конечностей и стоп, а также позвоночные центр. В материале было мало или совсем ничего истирание это могло произойти в результате транспортировки, то есть окаменелости были захоронены рядом с местом их происхождения.[30] Костные слои 23, 28, 47 и 50 являются продуктивными источниками остатков молодых гадрозавров в формации, особенно костного ложа 50. Кости молодых гадрозавров и ископаемые фрагменты яичной скорлупы не сохранились вместе друг с другом, несмотря на то, что оба они присутствуют в формирование.[31]
| Орнитоподы из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Коритозавр | C. casuarius | Нижнее-Среднее, 76,5–75,5 млн лет назад[8] | «Приблизительно [десять] сочлененных черепов и связанные с ними посткрании, [от десяти до пятнадцати] сочлененных черепов, отдельные элементы черепа, от юношеского до взрослого».[32] | А ламбеозаврин ламбеозаврин гадрозавр |  | |
| Грипозавр | G. notabilis | Lower, 76.2-76Ма назад[8] | «Примерно [десять] полных черепов, [двенадцать] фрагментарных черепов, связанных посткрании».[33] | А критозаврин зауролофин гадрозавр |  | |
| Ламбеозавр | L. lambei | Upper, 75.5-75Ма назад[8] | «Приблизительно [семь] сочлененных черепов с соответствующими посткраниями, [возможно, десять] сочлененных черепов, отдельные элементы черепа, от юных до взрослых».[34] | |||
| L. magnicristatus | Верхний /Формация Медвежьей Лапы, 74.8Ма назад[8] | «[Два] полных черепа, один с сочлененной посткранией».[34] |  | |||
| Паразауролоф | П. уокери | Нижний, 76.5-75.3 млн. Назад[7] | «Полный череп и посткраниальный скелет».[34] | А паразауролофин ламбеозавр гадрозавр. |  | |
| Прозавролоф | P. maximus | Верхний, 75,5 - 74,8 млн лет | «[От двадцати до двадцати пяти] человек, включая не менее [семи] сочлененных черепов и связанных с ними посткраний».[33] | А зауролофин зауролофин гадрозавр |  | |
Пахицефалозавры
| Пахицефалозавры из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Foraminacephale | F. brevis | Также присутствует в Формация Олдмана | Фронтопаретальный купол, различные другие фрагменты черепа, включая подростковый и подростковый материал | Когда-то считалось разновидностью Стегоцерасы |  | |
| Гравитолус | G. albertae | «Лобно-теменный купол».[35] |  | |||
| Hanssuesia | Х. штернберги | Нижний, также присутствует в Формация Олдмана и Формация реки Джудит |  | |||
| Стегоцерасы | С. валидум | Образцы, включая лобно-теменный купол.[35] |  | |||
Теропод
В формации «Парк динозавров» мелкие тероподы встречаются редко из-за того, что их тонкостенные кости имеют тенденцию ломаться или плохо сохраняться.[36] Маленькие кости маленьких тероподов, на которых охотились более крупные, могли быть проглочены целиком и переварены.[37] В этом контексте особенно ценным было открытие небольшого динозавра-теропод с сохранившимися следами зубов.[36] Возможные неопределенные останки авимимидов известны из формации.
Орнитомимиды
| Орнитомимиды из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Орнитомим | O. sp.[38] | Тип образца | An орнитомимид, возможно, вид Струтиомим.[39] |  | ||
| Qiupalong | Q. sp.[40] | Несколько экземпляров | An орнитомимид, возможно, радиация этого рода в Азию.[40] |  | ||
| Ратифицирует | Р. уклоняется | Тип образца | Орнитомимид, ранее являвшийся образцом Струтиомим.[41] |  | ||
Овирапторозавры
Цветовой ключ
| Примечания Неопределенные или предположительные таксоны входят в маленький текст; |
| Овирапторозавры из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Caenagnathus | C. collinsi | Нижняя челюсть, типовой образец | Ценагнатид[42] который соперничал Анзу по размеру.[43] | 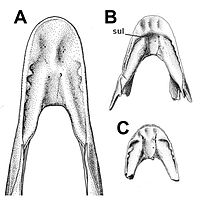 | ||
| Хиростеноты | C. pergracilis | Несколько фрагментарных экземпляров, типовой образец | Ценагнатид среднего размера. |  | ||
| Citipes | C. elegans[43] | Несколько фрагментарных экземпляров, типовой образец | Самый мелкий ценагнатид из свиты.[43] |  | ||
| Макрофалангия | M. canadensis | Младший синоним Хиростеноты pergracilis | ||||
Паравианцы
Новый таксон троодонтид основанный исключительно на зубах, известен из верхней части образования.[44]
| Паравианцы из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| ср. Бапторнис | Неопределенный | А гесперорнитин птица | ||||
| ср. Цимолоптерикс | Неопределенный | Частичный коракоид | Возможный рогоносец птица | |||
| Дромеозавр | D. albertensis | Несколько экземпляров и зубов, типовой образец | Дромеозаврид |  | ||
| Геспероних | H. elizabethae | Тазобедренные кости и частичные пальцы ног и когти, типовой образец | А микрорапторин дромеозавр, также встречается в Формация Олдмана |  | ||
| Латенивенатрикс | L. mcmasterae | Тазобедренные кости, таз, фрагменты черепа, типовой образец | Большой троодонтид размером 3–3,5 м (9,8–11,5 футов). |  | ||
| ср. Палинтроп | Безымянный | Неполные плечевые пояса | An амбиортиформ птица | |||
| ср. Парониходон | ср. P. lacustris | Зубы | Неопределенный манирапторан, также найденный в Река Джудит | |||
| ср. Пектинодон[45] | Неопределенный | Зубы | Троодонт | |||
| Полиодонтозавр | P. grandis | Стоматология, типовой образец | Nomen dubium. Возможно синоним Латенивенатрикс. | |||
| Ричардэстезия | Р. гилмори | Нижняя челюсть, типовой образец | Дромеозаврид | |||
| R. равнобедренный[44] | Зубы | |||||
| Saurornitholestes | S. langstoni | Неполный скелет и зубы, типовой образец. А зубной упомянутый Saurornitholestes Было обнаружено, что сохранились следы зубов, оставленные молодым тираннозавром.[46] | Дромеозаврид |  | ||
| Стенонихозавр | S. inequalis | Почти полный скелет и другие частичные скелеты, типовой образец | Троодонтид когда-то считался разновидностью Troodon |  | ||
Тираннозавры
| Тираннозавры из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место расположения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Дасплетозавр | Безымянный вид[47] | Средний и Верхний, 75.6-75 млн. Назад[8] | Несколько экземпляров | А тираннозаврин тираннозаврид, также присутствует в Формация Медвежьей Лапы |  | |
| Горгозавр | G. libratus | Нижнее-Среднее, 76,6–75,1 млн лет назад[8] | Многочисленные экземпляры, типовой образец[47] | An альбертозаурин тираннозаврид |  | |
Цветовой ключ
| Примечания Неопределенные или предположительные таксоны входят в маленький текст; |
Другие рептилии
Хористодерес
Хористодерес, или же шампсозавры, были водные рептилии. Маленькие экземпляры были похожи на ящериц, тогда как более крупные были внешне похожи на крокодилов. В свите обнаружены остатки следующих хористодеров:[48]
- Шампсозавр (минимум 3 вида)
- Cteniogenys
Крокодилы
В формации были обнаружены останки следующих крокодилов:[49]
- Альбертошампа
- Leidyosuchus
- как минимум 1 безымянный таксон
Ящерицы
В свите обнаружены останки следующих ящериц:[50]
Плезиозавры
В формации были обнаружены останки следующих плезиозавров:[51]
- неопределенный эласмозавриды (очень длинная шея) (чаще встречается)
- неопределенный поликотилиды (с короткой шеей)
Птерозавры
В свите обнаружены останки следующих птерозавров:[52]
- Криодракон [53] (известно по мелким и крупным экземплярам)
- 1 безымянный птерозавр, не являющийся аждархидом
Черепахи
В составе формации найдены останки следующих черепах:[54]
- Adocus
- "Apalone "
- Aspideretoides (3 вида)
- Basilemys
- Boremys
- Judithemys
- Neurankylus
- Плезиобаена
- 2 неопределенных таксона
Млекопитающие
В свите обнаружены останки следующих млекопитающих:[55]
- Мультитуберкулята
- Цимексомис sp.
- Цимолодон виды
- Чимоломис Кларки
- Meniscoessus основной
- Мезодма примаева
- безымянные мультитуберкуляты
- Метатерии
- Alphadon Аллей
- Эодельфис брауни
- Э. катлери
- 5 видов "Pediomys "
- Тургидодон Расселли
- T. praesagus
- Евтерийцы
- Cimolestes sp. (неопределенная таксономия)
- Гипсониктопс Левизи
- Paranyctoides Sternbergi
- Неизвестный терианцы: минимум 1 вид
Рыбы
В свите обнаружены остатки следующих рыб:[56]
- Хондрихтианцы
- Acipenseriformes (осетровые )
- безымянный осетр
- безымянный веслонос
- "Acipenser альбертенз"
- Anchiacipenser acanthaspis[58]
- Голосовой рыбы
- Belonostomus longirostris
- Lepisosteus западный (в гар )
- безымянный Bowfin
- как минимум 2 других голостея
- Teleost рыбы
- Паратарпон апогеронт (ан элопоморф, словно тарпон )
- Cretophareodus (ан остеоглоссоморф )
- Кориопс амниколус
- Estesesox foxi
- Oldmanesox
- Паральбула (включая Филлод)
- как минимум 8 других костистых
Беспозвоночные
В свите обнаружены остатки следующих беспозвоночных:[59]
- Пресная вода двустворчатые моллюски
- Пресная вода брюхоногие моллюски
- Campeloma (2 вида)
- Элимия
- Гониобазис (3 вида)
- Гидробия
- Лиоплакоды (2 вида)
Флора
Окаменелости тела растений
В формации были обнаружены следующие окаменелости растительных тел:[60]
- Голосеменные
- Платиспироксилон (Cupressaceae )
- Подокарпоксилон (Подокарповые )
- Элатоклад (Taxodiaceae )
- Секвойя (Taxodiaceae)
- Секвойаксилон (Taxodiaceae)
- Таксодиоксилон (Taxodiaceae)
- Гинкго
- Покрытосеменные
Палиноморфы
Палиноморфы имеют органические стены микрофоссилий, подобно споры, пыльца, и водоросли. В свите обнаружены следующие палиноморфы:[61]
- Неизвестные производители
- не менее 8 видов
- Хлорофита (зеленый водоросли и сине-зеленые водоросли )
- не менее 12 разновидность
- Pyrrhophyta (динофлагелляты, тип морской водоросли )
- неназначенный кисты
- Мохообразные (мхи, печеночники, и роголистник )
- Anthocerotophyta (роголистник)
- минимум 5 видов
- Мархантиофита (печеночники)
- не менее 14 видов
- Мохообразные (мхи)
- минимум 5 видов
- Anthocerotophyta (роголистник)
- Lycopodiophyta
- Lycopodiaceae (клубные мхи )
- не менее 11 видов
- Селагинелловые (мелкие клубные мхи)
- не менее 6 видов
- Isoetaceae (перья )
- минимум 1 вид
- Lycopodiaceae (клубные мхи )
- Полиподиофита
- Osmundaceae (коричный папоротник )
- не менее 6 видов
- Schizaeaceae (вьющиеся папоротники )
- не менее 20 видов
- Gleicheniaceae (Глейхения и союзники; коралловые папоротники )
- не менее 5 видов
- Cyatheaceae (Циатея и союзники)
- минимум 4 вида
- Dicksoniaceae (Диксония и союзники)
- минимум 3 вида
- Полиподиевые (папоротники )
- минимум 4 вида
- Matoniaceae
- минимум 1 вид
- Марсиловые
- минимум 1 вид
- Osmundaceae (коричный папоротник )
- Pinophyta (голосеменные )
- Cycadaceae (саговники )
- минимум 3 вида
- Caytoniaceae
- минимум 1 вид
- Pinaceae (сосны )
- минимум 4 вида
- Cupressaceae (кипарисы )
- минимум 3 вида
- Подокарповые (Подокарпус и союзники)
- минимум 4 вида
- Хейролепидиевые
- минимум 2 вида
- Ephedraceae (Мормонские чаи )
- не менее 6 видов
- Неизвестные голосеменные: минимум 3 вида.
- Cycadaceae (саговники )
- Магнолиофита (покрытосеменные )
- Магнолиопсида (двудольные )
- Buxaceae (самшит )
- минимум 1 вид
- Gunneraceae (Gunneras )
- минимум 1 вид
- Salicaceae (ивы, тополь, осина )
- минимум 1 вид
- Droseraceae (росянки )
- минимум 1 вид
- Olacaceae (талловое дерево )
- минимум 2 вида
- Loranthaceae (эффектные омелы )
- минимум 1 вид
- Sapindaceae (мыльное дерево )
- минимум 1 вид
- Aceraceae (клены )
- минимум 1 вид
- Proteaceae (протей )
- не менее 9 видов
- Сложноцветные (подсолнухи )
- минимум 1 вид
- Fagaceae (буки, дубы, каштаны )
- минимум 2 вида
- Betulaceae (березы, ольха )
- минимум 1 вид
- Ulmaceae (вяз )
- минимум 1 вид
- Chenopodiaceae (гусиные лапки )
- минимум 1 вид
- Buxaceae (самшит )
- Лилиопсида (однодольные )
- Лилии (лилии )
- не менее 6 видов
- Cyperaceae (осока)
- минимум 1 вид
- Sparganiaceae (тростник )
- возможно 1 вид
- Неизвестные покрытосеменные: не менее 88 видов.
- Лилии (лилии )
- Магнолиопсида (двудольные )
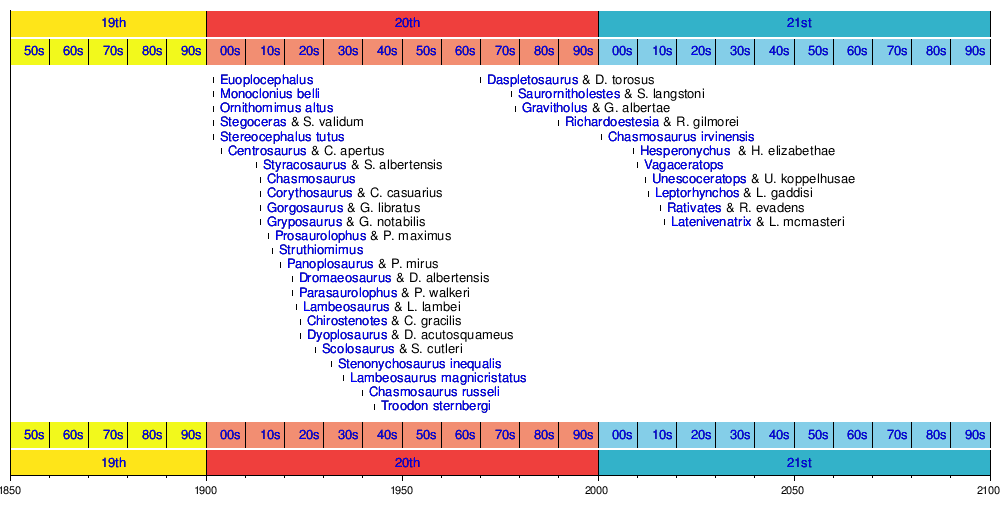
Хронология появления новых таксонов
На следующей временной шкале показаны действительные таксоны, впервые обнаруженные у динозавра. Некоторые виды могли быть отнесены к другим родам после их первоначального описания.
Смотрите также
Сноски
- ^ Лексикон канадских геологических единиц: Формирование парка динозавров В архиве 2013-02-21 в Archive.today
- ^ а б c Eberth, D.A .; Хамблин, А.П. (1993). «Тектоническое, стратиграфическое и седиментологическое значение регионального разрыва в верхней группе реки Джудит (клин реки Белли) на юге Альберты, Саскачевана и северной Монтаны». Канадский журнал наук о Земле. 30: 174–200. Дои:10.1139 / e93-016.
- ^ а б Фаулер, Д. (2016). Новая корреляция меловых образований западных внутренних районов США, I: сантонско-маастрихтские образования и биостратиграфия динозавров. Препринты Peer J.
- ^ а б c d Эберт, Д.А. 2005. Геология. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 54-82. ISBN 0-253-34595-2.
- ^ Карри П.Дж., Коппельхус Э. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 277-291. ISBN 0-253-34595-2.
- ^ Райан и Эванс (2005).
- ^ а б Evans D.C .; Bavington R .; Кампионе Н.Э. (2009). "Необычная черепная коробка гадрозаврид из формации парка динозавров и биостратиграфия Паразауролоф (Ornithischia: Lambeosaurinae) из южной Альберты ". Канадский журнал наук о Земле. 46 (11): 791–800. Bibcode:2009CaJES..46..791E. Дои:10.1139 / E09-050.[постоянная мертвая ссылка ]
- ^ а б c d е ж грамм час я j k л м п о п Беседка, В. М .; Burns, M.E .; Сиссонс, Р. Л. (2009). "Новое описание анкилозавридного динозавра. Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) и ревизия рода ». Журнал палеонтологии позвоночных. 29 (4): 1117–1135. Дои:10.1671/039.029.0405. S2CID 85665879.
- ^ а б c Пенкальски П. (2013). «Новый анкилозаврид из двухмедицинской формации позднего мелового периода в штате Монтана, США». Acta Palaeontologica Polonica. Дои:10.4202 / app.2012.0125.
- ^ Мэллон, Дж. К., Эванс, Д. К., Райан, М. Дж., И Андерсон, Дж. С. (2012). Круговорот мегатравоядных динозавров в формации парка динозавров (верхний кампан) в Альберте, Канада. Палеогеография, палеоклиматология, палеоэкология.
- ^ Гарднер, Дж. Д. 2005. Лиссамфибии. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 186-201. ISBN 0-253-34595-2.
- ^ «Fossilworks: вход в базу данных палеобиологии». fossilworks.org. Получено 23 апреля 2018.
- ^ «Fossilworks: вход в базу данных палеобиологии». fossilworks.org. Получено 23 апреля 2018.
- ^ Карри, П.Дж. 2005. Тероподы, включая птиц. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 367-397. ISBN 0-253-34595-2.
- ^ Райан, М.Дж., Эванс, округ Колумбия, 2005 г. Орнитисхийские динозавры. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 312-348. ISBN 0-253-34595-2.
- ^ «Таблица 17.1» в Weishampel, et al. (2004). Стр. 365.
- ^ Пенкальски, Пол (2018). «Пересмотренная систематика бронированного динозавра Euoplocephalus и его союзников». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 287 (3): 261–306. Дои:10.1127 / njgpa / 2018/0717.
- ^ Пенкальски, Пол (2013). «Новый анкилозаврид из двухмедицинской формации позднего мелового периода в штате Монтана, США». Acta Palaeontologica Polonica. Дои:10.4202 / app.2012.0125.
- ^ а б c «Таблица 23.1» в Weishampel, et al. (2004). Стр. 495.
- ^ а б «Таблица 23.1» в Weishampel, et al. (2004). Стр. 496.
- ^ а б c Райан, Майкл Дж .; Эванс, Дэвид С .; Карри, Филип Дж .; Лёвен, Марк А. (2014). «Новый хасмозаврин из северной Ларамидии увеличивает неравномерность оборок у динозавров цератопсид». Naturwissenschaften. 101 (6): 505–512. Дои:10.1007 / s00114-014-1183-1. PMID 24859020. S2CID 13957187.
- ^ а б c d Николас Р. Лонгрич (2014). "Рогатые динозавры Пентацератопс и Космоцератопс из верхнего кампана Альберты и значение для биогеографии динозавров ". Меловые исследования. 51: 292–308. Дои:10.1016 / j.cretres.2014.06.011.
- ^ Джордан К. Мэллон, Кристофер Дж. Отт, Питер Л. Ларсон, Эдвард М. Юлиано и Дэвид К. Эванс (2016). "Spiclypeus shipporum ген. et sp. nov., смелый новый хасмозавр цератопсид (Dinosauria: Ornithischia) из формации Джудит-Ривер (верхний мел: кампан) в Монтане, США ". PLOS ONE. 11 (5): e0154218. Bibcode:2016PLoSO..1154218M. Дои:10.1371 / journal.pone.0154218. ЧВК 4871577. PMID 27191389.CS1 maint: использует параметр авторов (связь)
- ^ а б c d е Farke, Andrew A .; Майкл Дж. Райан; Пол М. Барретт; Даррен Х. Танке; Деннис Р. Браман; Марк А. Лёвен; Марк Р. Грэм (2011). «Новый центрозаврин из позднего мела в провинции Альберта, Канада, и эволюция теменного орнамента у рогатых динозавров» (PDF). Acta Palaeontologica Polonica. 56 (4): 691–702. Дои:10.4202 / app.2010.0121. S2CID 13717580.
- ^ Майкл Дж. Райан; Дэвид С. Эванс; Филип Дж. Карри; Калеб М. Браун; Дон Бринкман (2012). «Новые лептоцератопсиды из верхнего мела Альберты, Канада». Меловые исследования. 35: 69–80. Дои:10.1016 / j.cretres.2011.11.018.
- ^ Скотт Д. Сэмпсон; Марк А. Лёвен; Эндрю А. Фарке; Эрик М. Робертс; Кэтрин А. Форстер; Джошуа А. Смит; Алан Л. Титус (2010). «Новые рогатые динозавры из штата Юта являются свидетельством внутриконтинентального эндемизма динозавров». PLOS ONE. 5 (9): e12292. Bibcode:2010PLoSO ... 512292S. Дои:10.1371 / journal.pone.0012292. ЧВК 2929175. PMID 20877459.
- ^ а б «Абстракция», Танке и Бретт-Сурман (2001). Стр.206.
- ^ «Выводы» Танке и Бретт-Сурман (2001). Стр. 212.
- ^ а б «Яичная скорлупа», Танке и Бретт-Сурман (2001). Стр.209.
- ^ «Введение», Танке и Бретт-Сурман (2001). Стр.208.
- ^ «Обсуждение», Танке и Бретт-Сурман (2001). Стр. 212.
- ^ «Таблица 20.1» в Weishampel, et al. (2004). Стр.441.
- ^ а б «Таблица 20.1» в Weishampel, et al. (2004). Стр. 440.
- ^ а б c «Таблица 20.1» в Weishampel, et al. (2004). Стр.442.
- ^ а б «Таблица 21.1» в Weishampel, et al. (2004). Стр. 465.
- ^ а б «Введение», Якобсен (2001). Стр.59.
- ^ «Обсуждение», Якобсен (2001). Стр.61.
- ^ Лонгрич, Н. Р. (2014). "Рогатые динозавры Пентацератопс и Космоцератопс из верхнего кампана Альберты и значение для биогеографии динозавров ". Меловые исследования. 51: 292. Дои:10.1016 / j.cretres.2014.06.011.
- ^ Лонгрич, Н. (2008). «Новый большой орнитомимид из формации мелового парка динозавров в провинции Альберта, Канада: значение для изучения диссоциированных останков динозавров». Палеонтология. 51 (4): 983–997. Дои:10.1111 / j.1475-4983.2008.00791.x.
- ^ а б McFeeters, B .; Райан, М. Дж .; Schröder-Adams, C .; Карри, П. Дж. (2017). «Первые случаи появления Qiupalong (Theropoda: Ornithomimidae) в Северной Америке и палеобиогеография производных орнитомимид». ЛИЦА. 2 (1): 355–373. Дои:10.1139 / facets-2016-0074.
- ^ McFeeters, B .; и другие. (2016). «Новый теропод-орнитомимид из формирования парка динозавров в Альберте, Канада». Журнал палеонтологии позвоночных. 36 (6): e1221415. Дои:10.1080/02724634.2016.1221415. S2CID 89242374.
- ^ Longrich, N. R .; Barnes, K .; Clark, S .; Миллар, Л. (2013). «Caenagnathidae из формации Aguja верхнего кампана в Западном Техасе и пересмотр Caenagnathinae». Бюллетень музея естественной истории Пибоди. 54: 23–49. Дои:10.3374/014.054.0102. S2CID 128444961.
- ^ а б c Фанстон, Грегори (27.07.2020). «Каенагнатиды формации парка динозавров (кампан) в Альберте, Канада: анатомия, остеогистология, таксономия и эволюция». Анатомия позвоночных Морфология Палеонтология. 8: 105–153. Дои:10.18435 / vamp29362. ISSN 2292-1389.
- ^ а б Санки, Юлия Т .; Бринкман, Дональд Б .; Гюнтер, Меррили; Карри, Филип Дж. (2002). "МАЛЫЙ ТЕРОПОД И ЗУБЫ ПТИЦЫ ИЗ ПОЗДНЕГО МЕЛОВА (ПОЗДНЕГО КАМПАНИЯ) ГРУППА РЕКИ ДЖУДИТ, АЛЬБЕРТА". Журнал палеонтологии. 76 (4): 751–763. Дои:10.1666 / 0022-3360 (2002) 0762.0.CO; 2. ISSN 0022-3360.
- ^ Карри, Филип Дж .; Ларсон, Дерек В. (23 января 2013 г.). "Многофакторный анализ зубов малых теропод динозавров и их значение для палеоэкологического круговорота во времени". PLOS ONE. 8 (1): e54329. Bibcode:2013PLoSO ... 854329L. Дои:10.1371 / journal.pone.0054329. ISSN 1932-6203. ЧВК 3553132. PMID 23372708.
- ^ «Абстракция», Якобсен (2001). Стр.58.
- ^ а б Карри, Филип Дж. (2003). "Анатомия черепа тираннозаврид из позднего мела Альберты". Acta Palaeontologica Polonica. 48 (2): 191–226.
- ^ К.Гао, Бринкман, Д. 2005. Хористодерес из парка и его окрестностей. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 221-234. ISBN 0-253-34595-2.
- ^ Сяо-Чун Ву. 2005. Крокодилы. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 277-291. ISBN 0-253-34595-2.
- ^ Колдуэлл М.В. Чешуйчатые: происхождение, филогения и палеоэкология. В: Currie, P.J., and Koppelhus, E.B. (ред.). 2005. ‘’ Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 235-248. ISBN 0-253-34595-2.
- ^ Сато Т., Эберт Д.А., Николлс Э.Л. и Манабе М. 2005. Плезиозавровые остатки от неморских до паралических отложений. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 249-276. ISBN 0-253-34595-2.
- ^ Годфри, С.Дж., Карри, П.Дж. 2005. Птерозавры. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 292-311. ISBN 0-253-34595-2.
- ^ Дэвид У. Э. Хоун; Майкл Б. Хабиб; Франсуа Терриен (2019). "Криодракон борей ген. et sp. ноя позднемеловой канадский птерозавр аждархид ". Журнал палеонтологии позвоночных. 39 (3): e1649681. Дои:10.1080/02724634.2019.1649681. S2CID 203406859.
- ^ Бринкман, Д. 2005. Черепахи: разнообразие, палеоэкология и распространение. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 202-220. ISBN 0-253-34595-2.
- ^ Фокс, Р. 2005. Млекопитающие позднего мела. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 417-435. ISBN 0-253-34595-2.
- ^ Нойман А.Г., Бринкман Д. 2005. Рыбы речных русел. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 167-185. ISBN 0-253-34595-2.
- ^ «Fossilworks: вход в базу данных палеобиологии». fossilworks.org. Получено 23 апреля 2018.
- ^ Хироки Сато; Элисон М. Мюррей; Оксана Вернигора; Филип Дж. Карри (2018). «Редкий осетр с сочленением (Chondrostei: Acipenseriformes) из верхнего мела Провинциального парка динозавров, Альберта, Канада». Журнал палеонтологии позвоночных. Интернет-издание (4): (1) - (15). Дои:10.1080/02724634.2018.1488137. S2CID 92574712.
- ^ Джонстон П.А., Хенди А.Дж.В. 2005. Палеоэкология моллюсков группы нижнего мела р. Белли. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 139-166. ISBN 0-253-34595-2.
- ^ Коппельхус, Э. 2005. Палеоботаника. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 131-138. ISBN 0-253-34595-2.
- ^ Браман, Д. Р., Коппельхус, Э. Б. 2005. Кампанские палиноморфы. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, стр. 101-130. ISBN 0-253-34595-2.
Рекомендации
- Беседка, В. М .; Burns, M.E .; Сиссонс, Р. Л. (2009). "Новое описание анкилозавридного динозавра. Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) и ревизия рода ». Журнал палеонтологии позвоночных. 29 (4): 1117–1135. Дои:10.1671/039.029.0405. S2CID 85665879.
- Браман, Д. Р., Коппельхус, Э. Б. 2005. Кампанские палиноморфы. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 101–130.
- Бринкман, Д. 2005. Черепахи: разнообразие, палеоэкология и распространение. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 202-220.
- Колдуэлл М.В. Чешуйчатые: происхождение, филогения и палеоэкология. В: Currie, P.J., and Koppelhus, E.B. (ред.). 2005. ‘’ Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 235–248.
- Карри, П.Дж. 2005. Тероподы, включая птиц. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 367–397.
- Карри П.Дж., Коппельхус Э. (ред.). 2005 г. Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 648 стр.
- Эберт, Д.А. 2005. Геология. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 54–82.
- Фокс, Р. 2005. Млекопитающие позднего мела. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 417-435.
- К. Гао, Бринкман, Д. 2005. Хористодерес из парка и его окрестностей. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 221–234.
- Гарднер, Дж. Д. 2005. Лиссамфибии. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 186-201.
- Годфри, С.Дж., Карри, П.Дж. 2005. Птерозавры. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 292-311.
- Джонстон П.А., Хенди А.Дж.В. 2005. Палеоэкология моллюсков группы нижнего мела р. Белли. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 139–166.
- Коппельхус, Э. 2005. Палеоботаника. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 131–138.
- Лексикон канадских геологических единиц. "Формирование парка динозавров". Архивировано из оригинал на 21.02.2013. Получено 2011-03-29.
- Нойман А.Г., Бринкман Д. 2005. Рыбы речных русел. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 167–185.
- Райан, М.Дж., Эванс, округ Колумбия, 2005 г. Орнитисхийские динозавры. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 312-348.
- Сато Т., Эберт Д.А., Николлс Э.Л. и Манабе М. 2005. Плезиозавровые остатки от неморских до паралических отложений. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 249-276.
- Танке, Д.Х., Бретт-Сурман, М.К. 2001. Свидетельства появления птенцов и гадрозавров размером с птенцов (Reptilia: Ornithischia) из провинциального парка динозавров (формация парка динозавров: кампания), Альберта, Канада. С. 206–218. В: Мезозойская жизнь позвоночных - новые исследования, вдохновленные палеонтологией Филиппа Дж. Карри. Под редакцией Д. Х. Танке и К. Карпентера. Издательство Индианского университета: Блумингтон. xviii + 577 с.
- Сяо-Чун Ву. 2005. Крокодилы. В: Currie, P.J., and Koppelhus, E.B. (ред.), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 277-291