Первичная структура белка - Protein primary structure

Первичная структура белка это линейная последовательность из аминокислоты в пептид или же белок.[1] По соглашению о первичной структуре белка сообщают, начиная с амино- -терминал (N) конец к карбоксил -конец (C) конец. Биосинтез белков чаще всего выполняется рибосомы в камерах. Пептиды также могут быть синтезированный в лаборатории. Первичные структуры белков могут быть прямая последовательность, или выводится из Последовательности ДНК.
Формирование
Биологические
Аминокислоты полимеризуются через пептидные связи с образованием длинных позвоночник, с выступающими вдоль него боковыми цепями различных аминокислот. В биологических системах белки производятся во время перевод по ячейке рибосомы. Некоторые организмы также могут производить короткие пептиды путем синтез нерибосомальных пептидов, которые часто используют аминокислоты, отличные от стандартных 20, и могут быть циклизованы, модифицированы и сшиты.
Химическая
Пептиды могут быть синтезирован химически с помощью ряда лабораторных методов. Химические методы обычно синтезируют пептиды в порядке, обратном (начиная с C-конца) биологическому синтезу белка (начиная с N-конца).
Обозначение
Последовательность белка обычно обозначается строкой букв, в которой перечислены аминокислоты, начиная с амино- -терминальный конец до карбоксил -конечный конец. Можно использовать трехбуквенный или однобуквенный код для обозначения 20 встречающихся в природе аминокислот, а также смесей или неоднозначных аминокислот (аналогично обозначение нуклеиновой кислоты ).[1][2][3]
Пептиды могут быть прямая последовательность, или выводится из Последовательности ДНК. Большой базы данных последовательностей теперь существуют, которые сопоставляют известные белковые последовательности.
| Аминокислота | 3 буквы[4] | 1 буква[4] |
|---|---|---|
| Аланин | Ала | А |
| Аргинин | Arg | р |
| Аспарагин | Asn | N |
| Аспарагиновая кислота | Жерех | D |
| Цистеин | Cys | C |
| Глютаминовая кислота | Glu | E |
| Глутамин | Gln | Q |
| Глицин | Gly | грамм |
| Гистидин | Его | ЧАС |
| Изолейцин | Иль | я |
| Лейцин | Лея | L |
| Лизин | Lys | K |
| Метионин | Встретились | M |
| Фенилаланин | Phe | F |
| Пролин | Pro | п |
| Серин | Сер | S |
| Треонин | Thr | Т |
| Триптофан | Trp | W |
| Тирозин | Тюр | Y |
| Валин | Вал | V |
| Символ | Описание | Представленные остатки |
|---|---|---|
| Икс | Любая аминокислота или неизвестная | Все |
| B | Аспартат или аспарагин | D, N |
| Z | Глутамат или глутамин | E, Q |
| J | Лейцин или изолейцин | Я, Л |
| Φ | Гидрофобный | V, I, L, F, W, M |
| Ω | Ароматный | F, W, Y, H |
| Ψ | Алифатический | V, I, L, M |
| π | Маленький | P, G, A, S |
| ζ | Гидрофильный | S, T, H, N, Q, E, D, K, R, Y |
| + | Положительно заряженный | K, R, H |
| - | Отрицательно заряженный | D, E |
Модификация
Как правило, полипептиды представляют собой неразветвленные полимеры, поэтому их первичная структура часто может быть определена последовательностью аминокислоты вдоль их позвоночника. Однако белки могут становиться сшитыми, чаще всего за счет дисульфидные связи, и первичная структура также требует указания сшивающих атомов, например, указания цистеины участвует в дисульфидных связях белка. Другие сшивки включают десмозин.
Изомеризация
Хиральные центры полипептидной цепи могут подвергаться рацемизация. Хотя это не меняет последовательность, это влияет на химические свойства последовательности. В частности, L-аминокислоты, обычно содержащиеся в белках, могут спонтанно изомеризоваться в атом сформировать D-аминокислоты, которые не расщепляются большинством протеазы. Кроме того, пролин может образовывать стабильные транс-изомеры по пептидной связи.

Посттрансляционная модификация
Наконец, белок может подвергаться различным посттрансляционные модификации, которые кратко описаны здесь.
N-концевая аминогруппа полипептида может быть модифицирована ковалентно, например,
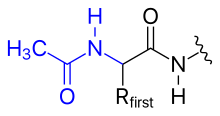
- ацетилирование

- Положительный заряд N-концевой аминогруппы можно устранить, заменив ее ацетильной группой (N-концевое блокирование).
- формилирование

- N-концевой метионин, обычно обнаруживаемый после трансляции, имеет N-конец, заблокированный формильной группой. Эта формильная группа (а иногда и сам остаток метионина, если за ним следует Gly или Ser) удаляется ферментом деформилаза.
- пироглутамат
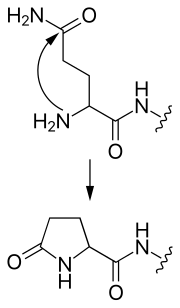
- N-концевой глутамин может атаковать сам себя, образуя циклическую пироглутаматную группу.
- миристоилирование

- Аналогично ацетилированию. Вместо простой метильной группы миристоильная группа имеет хвост из 14 гидрофобных атомов углерода, что делает ее идеальной для закрепления белков. клеточные мембраны.
С-концевая карбоксилатная группа полипептида также может быть модифицирована, например,
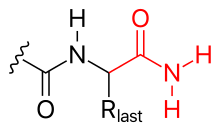
- аминирование (см. рисунок)
- С-конец также можно заблокировать (таким образом, нейтрализуя его отрицательный заряд) аминированием.
- гликозилфосфатидилинозитол (GPI) присоединение
- Гликозил фосфатидилинозитол (GPI) представляет собой большую гидрофобную простетическую группу фосфолипидов, которая прикрепляет белки к клеточные мембраны. Он присоединяется к С-концу полипептида через амидную связь, которая затем соединяется с этаноламином, затем с различными сахарами и, наконец, с липидным фрагментом фосфатидилинозитола.
Наконец, пептид боковые цепи также могут быть модифицированы ковалентно, например,
- фосфорилирование
- Помимо расщепления, фосфорилирование пожалуй, самая важная химическая модификация белков. Фосфатная группа может быть присоединена к гидроксильной группе боковой цепи остатков серина, треонина и тирозина, добавляя отрицательный заряд на этом участке и производя неприродную аминокислоту. Такие реакции катализируются киназы и обратная реакция катализируется фосфатазами. Фосфорилированные тирозины часто используются в качестве «ручек», с помощью которых белки могут связываться друг с другом, тогда как фосфорилирование Ser / Thr часто вызывает конформационные изменения, предположительно из-за введенного отрицательного заряда. Эффекты фосфорилирования Ser / Thr иногда можно моделировать путем мутации остатка Ser / Thr в глутамат.
- Универсальное название для набора очень распространенных и очень разнородных химических модификаций. Фрагменты сахара могут быть присоединены к гидроксильным группам боковой цепи Ser / Thr или к амидным группам боковой цепи Asn. Такие присоединения могут выполнять множество функций, от увеличения растворимости до сложного распознавания. Любое гликозилирование можно заблокировать определенными ингибиторами, такими как туникамицин.
- дезамидирование (образование сукцинимида)
- В этой модификации боковая цепь аспарагина или аспартата атакует следующую пептидную связь, образуя симметричный промежуточный сукцинимид. Гидролиз промежуточного продукта дает либо аспартат, либо β-аминокислоту изо (Asp). Для аспарагина любой из продуктов приводит к потере амидной группы, следовательно, к «дезамидированию».
- Остатки пролина могут быть гидроксилатами по любому из двух атомов, как и лизин (по одному атому). Гидроксипролин является важным компонентом коллаген, который становится нестабильным при его потере. Реакция гидроксилирования катализируется ферментом, который требует аскорбиновая кислота (витамин С), дефицит которого приводит ко многим заболеваниям соединительной ткани, таким как цинга.
- Некоторые белковые остатки могут быть метилированы, в первую очередь положительные группы лизин и аргинин. Остатки аргинина взаимодействуют с фосфатным остовом нуклеиновой кислоты и обычно образуют водородные связи с остатками оснований, в частности гуанин, в комплексах белок – ДНК. Остатки лизина могут быть метилированы отдельно, дважды и даже трижды. Метилирование делает нет однако изменить положительный заряд боковой цепи.
- Ацетилирование аминогрупп лизина химически аналогично ацетилированию N-конца. Однако функционально ацетилирование остатков лизина используется для регулирования связывания белков с нуклеиновыми кислотами. Отмена положительного заряда лизина ослабляет электростатическое притяжение (отрицательно заряженных) нуклеиновых кислот.
- сульфатирование
- Тирозины могут сульфатироваться на их атом. Несколько необычно эта модификация происходит в аппарат Гольджи, а не в эндоплазматический ретикулум. Подобно фосфорилированным тирозинам, сульфатированные тирозины используются для специфического распознавания, например, в рецепторах хемокинов на поверхности клетки. Как и в случае фосфорилирования, сульфатирование добавляет отрицательный заряд к ранее нейтральному сайту.

- пренилирование и пальмитоилирование

- Гидрофобные изопреновые группы (например, фарнезильная, геранильная и геранилгеранильная группы) и пальмитоильные группы могут быть добавлены к атом остатков цистеина для закрепления белков на клеточные мембраны. в отличие от GPI и миритоильные якоря, эти группы не обязательно добавляются на концах.

- карбоксилирование
- Относительно редкая модификация, которая добавляет дополнительную карбоксилатную группу (и, следовательно, двойной отрицательный заряд) к боковой цепи глутамата, образуя остаток Gla. Это используется для усиления связи с ионами «твердых» металлов, таких как кальций.
- АДФ-рибозилирование
- Большая ADP-рибозильная группа может переноситься на несколько типов боковых цепей в белках с гетерогенными эффектами. Эта модификация является мишенью для мощных токсинов разрозненных бактерий, например, Холерный вибрион, Коринебактерии дифтерии и Bordetella pertussis.
- Различные полноразмерные свернутые белки могут быть присоединены на своих С-концах к аммониевым группам боковой цепи лизинов других белков. Убиквитин является наиболее распространенным из них и обычно сигнализирует о том, что белок, меченный убиквитином, должен быть разрушен.
Большинство модификаций полипептидов, перечисленных выше, происходят посттрансляционный, т.е. после белок был синтезирован на рибосома, обычно встречающиеся в эндоплазматический ретикулум, субклеточный органелла эукариотической клетки.
Многие другие химические реакции (например, цианилирование) были применены к белкам химиками, хотя они не обнаруживаются в биологических системах.
Расщепление и лигирование
Помимо перечисленных выше, наиболее важной модификацией первичной структуры является пептидное расщепление (химическим гидролиз или по протеазы ). Белки часто синтезируются в неактивной форме-предшественнике; как правило, N-концевой или C-концевой сегмент блокирует активный сайт белка, подавляя его функцию. Белок активируется отщеплением ингибирующего пептида.
Некоторые белки даже обладают способностью расщепляться. Как правило, гидроксильная группа серина (редко треонина) или тиольная группа остатка цистеина будет атаковать карбонильный углерод предыдущей пептидной связи, образуя тетраэдрически связанный промежуточный продукт [классифицируемый как гидроксиоксазолидин (Ser / Thr) или гидрокситиазолидин ( Cys) промежуточный]. Этот промежуточный продукт имеет тенденцию превращаться в амидную форму, вытесняя атакующую группу, поскольку амидной форме обычно способствует свободная энергия (предположительно из-за сильной резонансной стабилизации пептидной группы). Однако дополнительные молекулярные взаимодействия могут сделать амидную форму менее стабильной; вместо этого удаляется аминогруппа, в результате чего вместо пептидной связи образуется сложноэфирная (Ser / Thr) или тиоэфирная (Cys) связь. Эта химическая реакция называется N-O ацильный сдвиг.
Связь сложный эфир / тиоэфир может быть разделена несколькими способами:
- Простой гидролиз расщепляет полипептидную цепь, где смещенная аминогруппа становится новым N-концом. Это видно по созреванию гликозиласпарагиназы.
- Реакция β-элиминирования также расщепляет цепь, но приводит к образованию пирувоильной группы на новом N-конце. Эта пирувоильная группа может использоваться в качестве ковалентно присоединенного каталитического кофактора в некоторых ферментах, особенно декарбоксилазах, таких как S-аденозилметионинкарбоксилаза (SAMDC), которые используют электроноакцепторную способность пировоильной группы.
- Внутримолекулярная переэтерификация, приводящая к разветвленный полипептид. В интеины, новая сложноэфирная связь разрывается внутримолекулярной атакой аспарагина, который вскоре станет С-концом.
- Межмолекулярная переэтерификация может переносить целый сегмент от одного полипептида к другому, как это видно при автопроцессинге белка Hedgehog.
История
Предположение, что белки представляют собой линейные цепи α-аминокислот, было сделано почти одновременно двумя учеными на той же конференции в 1902 году, 74-м собрании Общества немецких ученых и врачей, проходившем в Карлсбаде. Франц Хофмайстер сделал предложение утром, основываясь на своих наблюдениях за реакцией биурета в белках. Несколько часов спустя за Хофмайстером последовал Эмиль Фишер, который накопил множество химических деталей, подтверждающих модель пептидной связи. Для полноты картины предположение, что белки содержат амидные связи, было сделано еще в 1882 г. французским химиком Э. Гримо.[5]
Несмотря на эти данные и более поздние доказательства того, что протеолитически расщепленные белки давали только олигопептиды, идея о том, что белки представляют собой линейные неразветвленные полимеры аминокислот, не была принята сразу. Некоторые уважаемые ученые, такие как Уильям Эстбери сомневался, что ковалентные связи достаточно сильны, чтобы удерживать вместе такие длинные молекулы; они опасались, что тепловое возбуждение может расколоть такие длинные молекулы. Герман Штаудингер столкнулся с аналогичными предрассудками в 1920-х годах, когда утверждал, что резинка состоял из макромолекулы.[5]
Таким образом, возникло несколько альтернативных гипотез. В гипотеза коллоидного белка заявили, что белки представляют собой коллоидные сборки более мелких молекул. Эта гипотеза была опровергнута в 1920-х годах измерениями ультрацентрифугирования Теодор Сведберг которые показали, что белки имеют четко определенную воспроизводимую молекулярную массу, а с помощью электрофоретических измерений Арне Тизелиус это указывало на то, что белки были отдельными молекулами. Вторая гипотеза. циклол гипотеза продвинутый Дороти Ринч, предположили, что линейный полипептид подвергся химической перегруппировке циклола C = O + HN C (OH) -N, который сшивает амидные группы основной цепи, образуя двумерный ткань. Другие первичные структуры белков были предложены различными исследователями, такими как дикетопиперазиновая модель из Эмиль Абдерхалден и пиррол / пиперидин модель Троенсегаарда в 1942 году. Хотя эти альтернативные модели никогда не принимались во внимание, они были окончательно опровергнуты, когда Фредерик Сэнгер успешно секвенирован инсулин[когда? ] и кристаллографическим определением миоглобина и гемоглобина с помощью Макс Перуц и Джон Кендрю[когда? ].

Первичная структура в других молекулах
Можно сказать, что любой гетерополимер с линейной цепью имеет «первичную структуру» по аналогии с использованием термина для белков, но это использование редко по сравнению с чрезвычайно распространенным использованием в отношении белков. В РНК, который также имеет обширную вторичная структура линейную цепочку оснований обычно называют просто "последовательностью", как в ДНК (который обычно образует линейную двойную спираль с небольшой вторичной структурой). Другие биологические полимеры, такие как полисахариды также можно рассматривать как имеющую первичную структуру, хотя использование не является стандартным.
Отношение к вторичной и третичной структуре
Первичная структура биологического полимера во многом определяет его трехмерную форму (третичная структура ). Последовательность белка можно использовать для предсказывать местные особенности, такие как сегменты вторичной структуры или трансмембранные области. Однако сложность сворачивание белка в настоящее время запрещает прогнозирование третичной структуры белка только из его последовательности. Зная структуру подобного гомологичная последовательность (например, член той же белковая семья ) позволяет с высокой точностью прогнозировать третичная структура к моделирование гомологии. Если доступна полноразмерная последовательность белка, можно оценить ее общую биофизические свойства, например, его изоэлектрическая точка.
Семейства последовательностей часто определяются кластеризация последовательностей, и структурная геномика проекты направлены на создание набора репрезентативных структур для покрытия пространство последовательности возможных неизбыточных последовательностей.
Смотрите также
Примечания и ссылки
- ^ а б СЭНДЖЕР Ф (1952). Расположение аминокислот в белках. Adv. Protein Chem. Успехи в химии белков. 7. С. 1–67. Дои:10.1016 / S0065-3233 (08) 60017-0. ISBN 9780120342075. PMID 14933251.
- ^ Осланд, Рейн; Абрамс, Чарльз; Амп, Кристоф; Болл, Линда Дж .; Бедфорд, Марк Т .; Чезарени, Джанни; Джимона, Марио; Херли, Джеймс Х .; Ярчау, Томас (20 февраля 2002). «Нормализация номенклатуры пептидных мотивов как лигандов модульных белковых доменов». Письма FEBS. 513 (1): 141–144. Дои:10.1016 / S0014-5793 (01) 03295-1. ISSN 1873-3468.
- ^ Aasland R, Abrams C, Ampe C, Ball LJ, Bedford MT, Cesareni G, Gimona M, Hurley JH, Jarchau T., Lehto VP, Lemmon MA, Linding R, Mayer BJ, Nagai M, Sudol M, Walter U, Winder SJ (1968-07-01). «Однобуквенное обозначение аминокислотных последовательностей *». Европейский журнал биохимии. 5 (2): 151–153. Дои:10.1111 / j.1432-1033.1968.tb00350.x. ISSN 1432-1033. PMID 11911894.
- ^ а б Хаусман, Роберт Э .; Купер, Джеффри М. (2004). Клетка: молекулярный подход. Вашингтон, округ Колумбия: ASM Press. п. 51. ISBN 978-0-87893-214-6.
- ^ а б Fruton JS (май 1979 г.). «Ранние теории строения белков». Анна. Акад. Наука. 325: xiv, 1–18. Дои:10.1111 / j.1749-6632.1979.tb14125.x. PMID 378063.