Генетика и археогенетика Южной Азии - Genetics and archaeogenetics of South Asia
Генетика и археогенетика Южной Азии это исследование генетика и археогенетика из этнические группы Южной Азии. Он направлен на раскрытие этих групп. генетическая история. Географическое положение Южной Азии делает ее биоразнообразие важным для изучения раннего распространения анатомически современные люди через Азия.
Исследования на основе митохондриальной ДНК (мтДНК ) вариации сообщили о генетическом единстве среди различных южноазиатских субпопуляций.[1][2][3][4] Выводы исследований, основанных на вариации Y-хромосомы и вариабельности аутосомной ДНК, были разными, хотя многие исследователи утверждают, что большинство предковых узлов филогенетического древа всех типов мтДНК возникли в Южной Азии. Недавние исследования генома, по-видимому, показывают, что большинство жителей Южной Азии являются потомками двух основных предковых компонентов, один из которых ограничен Южной Азией (Предки южных индейцев, происходящие из IVC -люди и коренное население Южной Азии, отдаленно связанное с Андаманский ) и другой компонент (Предки северных индейцев) происходит от IVC-людей и степняков, что делает его более близким к Центральная Азия, Западная Азия и Европа.[5][6][7] Исследование 2016 года выявило еще два наследственных компонента на материковой части Индии, обозначенных как AAA и ATB, которые являются основными для племен, говорящих на австро-азиатском языке и говорящих на тибето-бирманском языке, соответственно. Исследование также предполагает, что популяции Андаманские острова архипелага образуют отдельную родословную, которая «была обнаружена коанцестрально по отношению к Океанический населения ".[8] Геномные исследования описали генетический ландшафт Южной Азии как смесь Западной Евразии и Восточная Азия экзогенные компоненты, которые смешались с группами коренных жителей Южной Азии, чтобы создать современных жителей Южной Азии.[9] Компонент восточноазиатского происхождения, обнаруженный в Индии, в основном ограничен определенными популяциями в предгорьях Гималаев и в северо-восточной части Индии.[10][11]
Установлено, что предковый узел филогенетического дерева всех мтДНК типы (гаплогруппы митохондриальной ДНК ), обычно встречающиеся в Центральной Азии, Западной Азии и Европе, также встречаются в Южной Азии с относительно высокой частотой. Предполагаемое расхождение этого общего предкового узла произошло чуть менее 50 000 лет назад.[12] В Индии основные материнские линии различны. M субклады, за которыми следует р и U подлинии. Время слияния этих митохондриальных гаплогрупп на сегодняшний день приближается к 50 000 лет назад.[12]
Основные отцовские линии, представленные Y-хромосомы гаплогруппы R1a1, R2, ЧАС, L и Гаплогруппа J2.[13] Некоторые исследователи утверждают, что гаплогруппа Y-ДНК R1a1 (M17) имеет автохтонный Южноазиатское происхождение.[14][15] Однако предложения о центральноазиатском происхождении R1a1 также довольно распространены.[16][17]
Обзор
Все линии мтДНК и Y-хромосомы за пределами Африки происходят от трех линий-основателей:
Все эти шесть гаплогрупп-основателей можно найти в современных популяциях Южная Азия. Более того, гаплогруппа M мтДНК и гаплогруппы C и D Y-хромосомы ограничены областью Восток Южной Азии. Весь Запад Евразийский популяции происходят от гаплогрупп N и R мтДНК и гаплогруппы F Y-хромосомы.[18]
Endicott et al. заявляют, что эти факты согласуются с гипотезой единственного исхода из Восточной Африки 65000 лет назад через южный прибрежный маршрут, причем западно-евразийские линии передачи отделяются от южноазиатских линий где-то между Восточной / Северо-Восточной Африкой и Южной Азией.[19]
Преобладающие маркеры большинства геномов жителей Южной Азии тесно связаны с Западные евразийцы и, возможно, возникли в Западная Азия или же Южная Азия сам.[20]
мтДНК

Самый частый гаплогруппы мтДНК в Южной Азии M, р и U (где U - потомок R).[13]
Аргументируя более долгосрочную «конкурирующую модель Y-хромосомы»,[14] Стивен Оппенгеймер считает, что это наводит на мысль, что Индия является источником Евразийский мтДНК гаплогруппы, которые он называет «евразийскими канунами». По мнению Оппенгеймера, весьма вероятно, что почти все человеческие материнские линии в Центральной Азии, на Ближнем Востоке и в Европе произошли только от четырех линий мтДНК, которые возникли в Южной Азии 50 000–100 000 лет назад.[21]
Макрогаплогруппа M
В макрогаплогруппа M, который рассматривается как кластер протоазиатских материнских линий,[12] представляет более 60% мтДНК Южной Азии.[22]
Макрогаплотип M в Индии включает множество подгрупп, которые сильно отличаются от других подлиний в Восточной Азии, особенно от монголоидных популяций.[12] Глубокие корни филогении M ясно указывают на реликвию южноазиатских линий по сравнению с другими подсистемами M (в Восточной Азии и в других местах), предполагая происхождение этих подгаплогрупп in-situ в Южной Азии, скорее всего, в Индии. Эти глубоко укоренившиеся линии не зависят от языка и распространяются на все языковые группы Индии.[22]
Практически все современные линии мтДНК М Центральной Азии принадлежат к восточно-евразийской (Монгольский ), а не южноазиатские подтипы гаплогруппы M, что указывает на отсутствие крупномасштабной миграции из настоящего Тюркский - говорящие народы Средней Азии пришли в Индию. Отсутствие гаплогруппы M у европейцев по сравнению с ее столь же высокой частотой среди выходцев из Южной и Восточной Азии и в некоторых популяциях Центральной Азии контрастирует с западно-евразийскими наклонностями южноазиатских отцовских линий.[12]
Большинство существующих границ мтДНК в Южной и Юго-Западной Азии, вероятно, были сформированы во время первоначального заселения Евразии анатомически современными людьми.[23]
| Гаплогруппа | Важные подпрограммы | Населения |
|---|---|---|
| M2 | M2a, M2b | По всему континенту с небольшим присутствием на Северо-Западе Пик в Бангладеш, Андхра-Прадеш, прибрежном Тамил Наду и Шри-Ланке |
| M3 | M3a | Сосредоточен в северо-западной Индии Самый высокий среди парсов Мумбаи |
| M4 | M4a | Вершины в Пакистане, Кашмире и Андхра-Прадеше |
| M6 | M6a, M6b | Кашмир и у берегов Бенгальского залива, Шри-Ланка |
| M18 | По всей Южной Азии Пик в Раджастане и Андхра-Прадеше | |
| M25 | Умеренно часто в Керале и Махараштре, но довольно редко в других местах Индии |
Макрогаплогруппа R



В макрогаплогруппа R (очень большое и старое подразделение макрогаплогруппа N ) также широко представлена и составляет остальные 40% мтДНК Южной Азии. Очень старое и самое важное его подразделение - гаплогруппа U что, хотя и присутствует в Западная Евразия, имеет несколько субкладов, характерных для Южной Азии.
Наиболее важные южноазиатские гаплогруппы в R:[23]
| Гаплогруппа | Населения |
|---|---|
| R2 | Широко распространен по субконтиненту |
| R5 | широко распространен на большей части территории Индии. Пики в прибрежной юго-западной Индии |
| R6 | широко распространен по низким ценам по всей Индии. Пики среди тамилов и кашмирцев |
| W | Встречается в северо-западных штатах. Пики в Гуджарате, Пенджабе и Кашмире, частота невысока в других местах. |
Гаплогруппа U
Гаплогруппа U является субгаплогруппой макрогаплогруппы R.[23] Распределение гаплогруппы U является зеркальным отражением распределения гаплогруппы M: первая до сих пор не была описана среди жителей Восточной Азии, но часто встречается в европейских популяциях, а также среди выходцев из Южной Азии.[24] Южноазиатские линии U существенно отличаются от европейских, и их слияние с общим предком также насчитывает около 50 000 лет.[1]
| Гаплогруппа | Населения |
|---|---|
| U2 * | (а парагаплогруппа ) редко распространен, особенно в северной половине Южной Азии. Он также встречается на юго-западе Аравии. |
| U2a | показывает относительно высокую плотность в Пакистане и на северо-западе Индии, но также и в Карнатаке, где она достигает более высокой плотности. |
| U2b | имеет самую высокую концентрацию в Уттар-Прадеше, но также встречается во многих других местах, особенно в Керале и Шри-Ланке. Также встречается в Омане. |
| U2c | особенно важен в Бангладеш и Западной Бенгалии. |
| U2l | является, пожалуй, самым важным по численности среди субкладов урана в Южной Азии, достигая особенно высоких концентраций (более 10%) в Уттар-Прадеше, Шри-Ланке, Синде и некоторых частях Карнатаки. Это также имеет некоторое значение в Омане. мтДНК гаплогруппа U2i в Бамшаде называют "западноевразийским" и другие. учиться, но "Восточно-Евразийский (в основном индийский)" в Кивисилде и другие. изучать. |
| U7 | эта гаплогруппа широко представлена в Гуджарате, Пенджабе и Пакистане. Возможная родина этой гаплогруппы охватывает Гуджарат (самая высокая частота, 12%) и Иран, потому что оттуда ее частота резко снижается как на восток, так и на запад. |
Y-хромосома
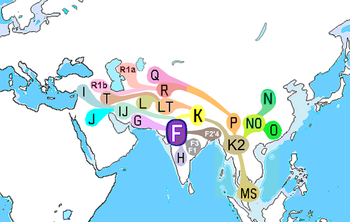
Основными гаплогруппами ДНК Y-хромосомы Южной Азии являются: ЧАС, J2, L, R1a1 и R2.[13] Согласно последним исследованиям, их географическое происхождение указано ниже:
| Основные южноазиатские линии Y-хромосомы: | ЧАС | J2 | L | R1a | R2 |
|---|---|---|---|---|---|
| Басу и другие. (2003) | нет комментариев | нет комментариев | нет комментариев | Центральная Азия | нет комментариев |
| Кивисилд и другие. (2003) | Индия | Западная Азия | Индия | Южная и Западная Азия | Юго-Центральная Азия |
| Кордо и другие. (2004) | Индия | Западная или Центральная Азия | Ближневосточный | Центральная Азия | Юго-Центральная Азия |
| Сенгупта и другие. (2006) | Индия | Ближний Восток и Центральная Азия | Южная Индия | Северная Индия | Северная Индия |
| Thanseem и другие. (2006) | Индия | Левант | Ближний Восток | Южная и Центральная Азия | Южная и Центральная Азия |
| Sahoo и другие. (2006) | Южная Азия | Ближний Восток | Южная Азия | Южная или Западная Азия | Южная Азия |
| Мирабал и другие. (2009) | нет комментариев | нет комментариев | нет комментариев | Северо-Западная Индия или Средняя Азия | нет комментариев |
| Чжао и другие. (2009) | Индия | Ближний Восток | Ближний Восток | Центральная Азия или Западная Евразия | Центральная Азия или Западная Евразия |
| Шарма и другие. (2009) | нет комментариев | нет комментариев | нет комментариев | Южная Азия | нет комментариев |
| Тангарадж и другие. (2010) | Южная Азия | Ближний Восток | Ближний Восток | Южная Азия | Южная Азия |
Гаплогруппа H
Гаплогруппа H (Y-ДНК) с высокой частотой встречается в Южной Азии. H сегодня редко встречается за пределами Южной Азии, но распространен среди Романис, особенно подгруппа H-M82. H также был довольно распространен в древних образцах Европы и до сих пор встречается редко у европейцев и арабов Левант. Гаплогруппа H часто встречается среди населения Индия, Шри-Ланка, Непал, Пакистан и Мальдивы. Все три ветви Гаплогруппа H (Y-ДНК) находятся в Южной Азии.
Это филиал Гаплогруппа F и спускается из GHIJK семья. Гаплогруппа H считается, что возник в Южной Азии между 30 000 и 40 000 лет назад.[25] Вероятное место интродукции - Южная Азия, так как она сосредоточена там. Похоже, что он представляет собой главную гаплогруппу Y-хромосомы палеолитических жителей Южной Азии. Некоторые особи в Южной Азии также принадлежат к гораздо более редкому субкладу H3 (Z5857).[25] Гаплогруппа H никоим образом не ограничивается определенными группами населения. Например, H принадлежит примерно 28,8% индоарийских каст.[14][26] а в племенах около 25–35%.[16][26]
Гаплогруппа J2
Гаплогруппа J2 присутствовала в Южной Азии в основном как J2a-M410 и J2b-M102 со времен неолита (9500 YBP).[27][28] Клады J2 достигают пиковых частот в Северо-Западной и Южной Индии.[27] и находится в 19% в кастах Южной Индии, 11% в кастах Северной Индии и 12% в Пакистане.[14] В Южная Индия, присутствие J2 выше среди средних каст - 21%, за ним следуют высшие касты - 18,6% и низшие - 14%.[14] Среди кастовых групп самая высокая частота J2-M172 наблюдается среди тамильских Веллаларс Южной Индии - 38,7%.[14] J2 присутствует и в трайблах[27] и имеет частоту 11% у австро-азиатских племен. Среди австро-азиатских племен преобладающие J2 встречается в Лодхе (35%).[14] J2 также присутствует в Южной Индии. горное племя Тода на 38,46%,[29] в Племя андх из Телангана на 35,19%[16] и в Племя кол из Уттар-Прадеш с частотой 33,34%.[30] Гаплогруппа J-P209 оказалась более распространенной в Индии. Мусульмане-шииты, из которых 28,7% принадлежат гаплогруппе J, 13,7% - J-M410, 10,6% - J-M267 и 4,4% - J2b (Eaaswarkhanth 2009 ).
В Пакистан, самые высокие частоты J2-M172 наблюдались среди Парсы при 38,89% Дравидийский Говорящий Brahuis на 28,18% и Макрани Белуджи на 24%.[31] Это также встречается в 18,18% в Макрани Сиддис и на 3% в Карнатака Сиддис.[31][32]
J2-M172 встречается с общей частотой 10,3% среди Сингальский народ из Шри-Ланка.[33] В Мальдивы 20,6% мальдивского населения оказались положительными по гаплогруппе J2.[34]
Гаплогруппа L
По словам доктора Спенсер Уэллс, L-M20 возник в Памирский узел регион в Таджикистан и перешел в Пакистан и Индия ок. 30 000 лет назад.[35][36][37] Однако в большинстве других исследований предлагается Западная Азия происхождение L-M20 и связанное с этим расширение в Долина Инда (~ 7000 YBP) до неолит фермеры.[31][38][16][26][39][40] Есть три подветви гаплогруппы L: L1-M76 (L1a1), L2-M317 (L1b) и L3-M357 (L1a2), обнаруженные на разных уровнях в Южной Азии.[14]
Индия
Гаплогруппа L показывает время неолитической экспансии.[41] Клада присутствует в индейском населении с общей частотой ок. 7–15%.[14][16][42][26] Гаплогруппа L чаще встречается среди южноиндийских каст (около 17–19%) и достигает 68% в некоторых кастах в Карнатака но несколько реже в кастах Северной Индии (около 5–6%).[14] Присутствие гаплогруппы L среди племенных групп довольно редко (около 5,6–7%),[14][16][26] однако умеренный, 14,6% наблюдался среди Ченч.[33]
Среди региональных и социальных групп частота от умеренной до высокой наблюдалась у браминов Конканастха (18,6%), пенджабцев (12,1%), гуджаратцев (10,4%), Ламбадис (17.1%), Джатс (36.8%)[33][36]
Пакистан
В Пакистане субклады L1-M76 и L3-M357 L-M20 достигают общих частот 5,1% и 6,8% соответственно.[14]Гаплогруппа L3 (M357) часто встречается среди Burusho (около 12%[43]) и Пуштуны (около 7%[43]). Его самая высокая частота встречается на юго-западе Белуджистан провинция вдоль Макран побережье (28%) до Река Инд дельта. L3a (PK3) встречается примерно в 23% Нуристани на северо-западе Пакистан.[43]
Клада присутствует в умеренном распределении среди населения Пакистана в целом (примерно 14%).[44][45]
Шри-Ланка
В одном исследовании 16% сингальцев оказались положительными по гаплогруппе L-M20.[46] В другом исследовании было обнаружено, что 18% принадлежат к L1.[33]
Гаплогруппа R1a1
В Южной Азии R1a1 часто с высокой частотой наблюдается в ряде демографических групп,[15][47][48] а также с высшим STR разнообразие, которое заставляет некоторых рассматривать его как локус происхождения.[33][30][49]
В то время как R1a возник ок. 22 000[30] до 25 000[50] лет назад его субклад M417 (R1a1a1) диверсифицировался ок. 5800 лет назад.[50] Распределение M417-субкладов R1-Z282 (включая R1-Z280)[51] в Центральной и Восточной Европе и R1-Z93 в Азии[51][50] предполагает, что R1a1a диверсифицирована в Евразийские степи или Средний Восток и Кавказ область, край.[51] Место происхождения этих субкладов играет роль в дебатах о происхождении Индоевропейцы.
Индия
В Индия, высокий процент этой гаплогруппы наблюдается в Брахманы Западной Бенгалии (72%)[47] на восток, Гуджарат Лоханас (60%)[48] на запад, Хатрис (67%)[48] на севере, Айенгарские брамины (31%) на юге.[47] Он также был обнаружен в нескольких Южноиндийский Дравидийский -Говорящий племена в том числе коты (41%) штата Тамил Наду[52] Ченчу (26%) и Валмикис из Андхра-Прадеш[33] так же хорошо как Ядав и Каллар из Тамил Наду предполагая, что M17 широко распространен в этих племенах южных индейцев.[33] Помимо этого, исследования показывают высокий процент в группах, различающихся по регионам, таких как Манипури (50%)[48] на крайний северо-восток и среди Пенджабцы (47%)[33] на крайний северо-запад.
Пакистан
В Пакистане он составляет 71% среди Моханны из Провинция Синд на юг и 46% среди Baltis из Гилгит-Балтистан на север.[48]
Шри-Ланка
23% Сингальский народ Согласно исследованию 2003 года, из выборки 87 субъектов был обнаружен R1a1a (R-SRY1532) положительный.[46]
Мальдивы
В Мальдивы, 23,8% Мальдивский народ оказались R1a1a (M17) положительными.[34]
Непал
Люди в Тераи Область, край, Непал показать R1a1a на 69%.[53]
Гаплогруппа R2
В Южной Азии частота R2 и R2a родословная составляет около 10–15% в Индии и Шри-Ланка и 7–8% в Пакистане. По крайней мере, 90% индивидов R-M124 находятся в Южной Азии.[54] Об этом также сообщается в Кавказ и Центральная Азия на более низкой частоте. Генетическое исследование Mondal et al. 2017 пришел к выводу, что гаплогруппа Гаплогруппа R2 возникла в северной Индии и существовала еще до переселения степей.[55]
Индия
Среди региональных групп он встречается среди Запад Бенгальцы (23%), Нью-Дели Индусы (20%), Пенджабцы (5%) и Гуджаратцы (3%).[33] Среди племенных групп Кармалис Западная Бенгалия показал самый высокий на 100%[15] с последующим Лодха (43%)[56] на восток, а Bhil из Гуджарат на западе - 18%,[30] Tharus севера - 17%,[4] Ченчу и Паллан юга были на 20% и 14% соответственно.[15][14] Среди кастовых групп высокий процент показывают Jaunpur Кшатрии (87%), камма (73%), Бихар Ядав (50%), Хандаят (46%) и Каллар (44%).[15]
Кроме того, во многих Брамин группы в том числе Пенджабские брамины (25%), Бенгальские брамины (22%), Конканастха Брамины (20%), Чатурведи (32%), Бхаргавы (32%), Кашмирские пандиты (14%) и Лингаят Брамины (30%).[30][17][4][15]
Мусульмане Северной Индии имеют частоту 19% (Сунниты ) и 13% (Шииты ),[17] пока Давуди Бора Муслим в западном штате Гуджарат имеют частоту 16% и Маппила мусульман Южной Индии имеют частоту 5%.[57]
Пакистан
Гаплогруппа R2 встречается в 14% Люди бурушо.[43] Среди Люди хунза он находится на уровне 18%, в то время как Парсы покажите это на 20%. Встречается также в северо-восточной части Афганистан.[нужна цитата ]
Шри-Ланка
38% Сингальский из Шри-Ланки, согласно исследованию 2003 года, оказался R2-положительным.[33]
Мальдивы
12% Мальдивский народ Мальдивских островов есть R2.[34]
Непал
В Непале процентное соотношение R2 колеблется от 2% до 26% в разных группах согласно различным исследованиям. Newars показывают значительно высокую частоту - 26%, в то время как люди Катманду покажите это на 10%.
Реконструкция истории населения Южной Азии
В Индийский консорциум по изменению генома (2008 г.), делит население Южной Азии на четыре этнолингвистические группы: Индоевропейский, Дравидийский, Тибето-бирманский и Австро-азиатский.[58][59][60][61][62] В исследованиях молекулярной антропологии используются три разных типа маркеров: вариация митохондриальной ДНК (мтДНК), которая наследуется по материнской линии и является высокополиморфной, вариация Y-хромосомы, которая включает однопародительскую передачу по мужским линиям, и вариация аутосомной ДНК.[4]:04
вариация мтДНК
Большинство исследований, основанных на вариации мтДНК, сообщают о генетическом единстве южноазиатских популяций по языку, кастам и племенным группам.[1][2][3] Вполне вероятно, что гаплогруппа M была принесена в Азию из Восточной Африки южным маршрутом самой ранней волной миграции 78000 лет назад.[1]
В соответствии с Кивисилд и другие. (1999): «Незначительные совпадения с линиями, описанными в других евразийских популяциях, ясно демонстрируют, что недавние иммиграции оказали очень небольшое влияние на врожденную структуру материнской Генофонд жителей Южной Азии. Несмотря на различия, обнаруженные в Индии, эти популяции происходят от ограниченного числа родословных основателей. Эти линии, вероятнее всего, были занесены в Южную Азию в период среднего палеолита, до заселения Европы 48 000 лет назад и, возможно, Старого Света в целом ».[1] Basu et al. (2003) также подчеркивает основополагающее единство женских линий в Индии.[42]
Вариация Y-хромосомы
Выводы, основанные на вариации Y-хромосомы, были более разнообразными, чем на вариации мтДНК. Пока Kivisild et al. (2003) предлагает древнее и общее генетическое наследие мужских линий в Южной Азии, Bamshad et al. (2001) предполагает сходство между южноазиатскими мужскими линиями и западными евразийцами, пропорциональное высшему кастовому рангу, и помещает высшие кастовые группы южных индийских штатов ближе к Восточноевропейцы.[63]
Basu et al. (2003) пришли к выводу, что австро-азиатские племенные народы вошли в Индию сначала из Северо-Западного коридора, а гораздо позже некоторые из них - через Северо-Восточный коридор.[42] Принимая во внимание, что Kumar et al. (2007) проанализировали 25 южноазиатских австро-азиатских племен и обнаружили сильную отцовскую генетическую связь между сублингвистическими группами австро-азиатского населения Южной Азии.[56] Mukherjee et al. (2001) помещает пакистанцев и северных индейцев между западноазиатским и центральноазиатским населением,[64] тогда как Cordaux et al. (2004) утверждает, что индийские кастовые популяции ближе к среднеазиатским.[26] Sahoo et al. (2006) и Sengupta et al. (2006) предполагают, что кастовое население Индии не подвергалось каким-либо примесям в последнее время.[14][15] Сангхамитра Саху завершает свое исследование следующими словами:[15]
Судя по имеющимся данным, нет необходимости искать истоки отцовского наследия большинства индейцев в период зарождения оседлого земледелия за пределами Южной Азии. Многолетняя концепция людей, языка и сельского хозяйства, прибывающих в Индию вместе через северо-западный коридор, не выдерживает тщательного изучения. Недавние утверждения о связи гаплогрупп J2, L, R1a и R2 с одновременным происхождением большинства отцовских линий индийских каст из-за пределов Южной Азии отвергаются, хотя наши результаты подтверждают местное происхождение гаплогрупп F * и H. Из остальных только J2 указывает на недавний недвусмысленный внешний вклад из Западной Азии, а не из Центральной Азии. Текущее распределение частот гаплогрупп, за исключением родословных, в основном определяется географическими, а не культурными детерминантами. По иронии судьбы, именно на северо-востоке Индии, среди групп туберкулеза, есть четкие доказательства крупномасштабного распространения демика, прослеживаемого по генам, культуре и языку, но, очевидно, не по сельскому хозяйству.
Анализ ближайшего соседа, проведенный Mondal et al. 2017 пришел к выводу, что индийская линия Y близка к южной. Европейский популяции и время расхождения между двумя предшествующими степными миграциями. ":[65]
Эти результаты предполагают, что европейские предки в индийском населении могут быть намного старше и сложнее, чем предполагалось, и могут происходить от земледельцев первой волны или даже раньше.
— Mondal et al. 2017 г.
Аутосомная вариация ДНК
ААСИ-АНИ-АСИ
Результаты исследований, основанных на вариабельности аутосомной ДНК, также различались. В крупном исследовании (2009 г.), в котором использовалось более 500 000 биаллельных аутосомных маркеров, Райх выдвинул гипотезу о том, что современное население Южной Азии возникло в результате смешения двух генетически расходящихся предковых популяций, датируемых постголоценовой эпохой. Эти два «реконструированных» древних населения он назвал «предками южных индейцев» (ASI) и «предками северных индейцев» (ANI). По словам Райха: «Родословная ANI значительно выше у индоевропейцев, чем у носителей дравидийского языка, что позволяет предположить, что предки ASI могли говорить на дравидийском языке до того, как смешались с ANI». В то время как ANI генетически близок к жителям Ближнего Востока, Центральной Азии и европейцам, ASI не имеет тесных связей с группами за пределами субконтинента. Поскольку древняя ДНК "ASI" недоступна, коренные Андаманский Онге используются в качестве (несовершенного) представителя ASI (согласно Райху и др., андаманцы, хотя и отличаются от них, являются ближайшим живым населением к ASI). Согласно Райху и др., Предки ANI и ASI встречаются по всему субконтиненту (как на севере, так и на юге Индии) в различных пропорциях, и что «предки ANI колеблются от 39 до 71% в Индии и выше в традиционно верхних регионах. касты и индоевропейские спикеры ".[66]
Moorjani et al. 2013 заявляют, что ASI, хотя и не связаны тесно с какой-либо живой группой, «связаны (отдаленно) с коренными жителями Андаманских островов». Moorjani et al. также предполагают возможный приток генов к андаманцам из популяции, связанной с ASI. Исследование пришло к выводу, что «почти все группы, говорящие на индоевропейских или дравидийских языках, лежат на градиенте разной степени родства с западноевразийцами в ППЦА (именуемой« индийская клина »)».[67]
Исследование, проведенное в 2013 году с использованием однонуклеотидного полиморфизма (SNP), показывает, что геном андаманцев (онге) ближе к геномам других океанических групп негрито, чем к геномам жителей Южной Азии.[68]
По данным Basu et al. В 2016 году дальнейший анализ показал, что геномная структура населения материковой Индии лучше всего объясняется вкладом четырех предковых компонентов. В дополнение к ANI и ASI, Basu et. al (2016) идентифицировал два предковых компонента на материковой части Индии, которые являются основными для племен, говорящих на австро-азиатском языке и говорящих на тибето-бирманском языке, которые они обозначили как AAA (от «Ancestral Austro-Asiatic») и ATB (от «Ancestral Tibeto» -Бурман ») соответственно. Исследование также предполагает, что популяции Андаманские острова архипелага образуют отдельную родословную, которая «была обнаружена коанцестрально Океанический населения ".[8]
Граница смешения между линиями ANI и ASI датируется периодом c. 4,2–1,9 тыс. Лет назад, Moorjani et al. (2013), что соответствует индийскому бронзовому веку и связывается авторами с процессом деурбанизации Цивилизация долины Инда и перемещение населения в систему Ганга в зарождающемся индийском железном веке.[6] Basu et al. (2003) предполагает, что «дравидийские племена, возможно, были широко распространены по всей Индии до прихода кочевников, говорящих на индоевропейском языке» и что «формирование популяций путем деления, которое привело к эффектам основателя и дрейфа, наложило свой отпечаток на генетические структуры современных населения ".[42] Генетик П.П. Маджумдер (2010) недавно утверждал, что результаты Райха и др. (2009) находятся в замечательном соответствии с предыдущими исследованиями с использованием мтДНК и Y-ДНК:[69]
Предполагается, что население Центральной Азии внесло основной вклад в генофонд Индии, особенно в генофонд северной Индии, а мигранты предположительно перебрались в Индию через территорию, которая сейчас называется Афганистаном и Пакистаном. С помощью митохондриальная ДНК Данные по вариациям, сопоставленные с различными исследованиями, показали, что популяции Центральной Азии и Пакистана демонстрируют самый низкий коэффициент генетической дифференциации с популяциями северной Индии, более высокий коэффициент дифференциации с популяциями южной Индии и самый высокий с популяциями северо-востока Индии. Популяции Северной Индии генетически ближе к жителям Центральной Азии, чем популяции других географических регионов Индии ... В соответствии с вышеизложенными выводами, недавнее исследование с использованием более 500000 двуаллельных аутосомных маркеров обнаружило градиент генетической близости индейских популяций и западных евразийцев с севера на юг. Эта особенность, вероятно, связана с пропорциями предков, происходящих от генофонда западной Евразии, который, как показало это исследование, больше у популяций, населяющих северную Индию, чем у тех, которые населяют южную Индию.
Chaubey et al. В 2015 году был обнаружен характерный восточноазиатский наследственный компонент, в основном ограниченный конкретными популяциями в предгорьях Гималаев и северо-восточной части Индии. Самая высокая частота этого компонента наблюдается среди тибето-бирманских говорящих групп северо-востока, а также обнаружена в андаманских популяциях, причем значительное присутствие среди австроазиатских говорящих. Обнаружено, что он в значительной степени отсутствует у индоевропейских и дравидийских носителей, за исключением некоторых конкретных этнических групп, живущих в предгорьях Гималаев и в центральной и южной частях Индии.[10] Однако исследователи предположили, что восточноазиатская родословная (представленная ханьцами), измеренная в изученных андаманских группах, может на самом деле отражать связь андаманцев с меланезийцами и малазийскими негритосами (а не истинную восточноазиатскую примесь).[10] как и в предыдущем исследовании Chaubey et al. предположил «глубокое общее происхождение» андаманцев, меланезийцев и других негритянских групп (а также жителей Южной Азии),[10] и родство негритосов и меланезийцев Юго-Восточной Азии (а также андаманцев) с жителями Восточной Азии.[68]
Lazaridis et al. (2016) отмечает: «Демографическое воздействие степных популяций на Южную Азию было значительным, поскольку Мала, южноиндийский Далит популяция с минимальным количеством предков северных индейцев (ANI) вдоль `` индийской линии '' такой родословной, как предполагается, имеет ~ 18% степное происхождение, в то время как Калаш предполагается, что ~ 50% родословных из Пакистана имеют степное происхождение ".[70] В исследовании Lazaridis et al., Проведенном в 2016 году, оценивается (6,5–50,2%) примесь, связанная со степью, у жителей Южной Азии. Lazaridis et al. далее отмечает, что «полезным направлением будущих исследований является более полный отбор образцов древней ДНК у степных популяций, а также у популяций Центральной Азии (к востоку от Ирана и к югу от степи), что может выявить более близкие источники ANI, чем рассмотренные здесь, и Южной Азии, чтобы напрямую определить траекторию изменения населения в этом районе.
Pathak et al. 2018 пришел к выводу, что Индоевропейский спикеры Гангские равнины и Дравидийский спикеры имеют значительные Ямная эпоха ранней-средней бронзы (Степная_EMBA) родословная, но не степная (Степная_МБА) родословная среднего-позднего бронзового века. С другой стороны, популяции «северо-западных индейцев и пакистанцев» (PNWI) показали значительное происхождение Steppe_MLBA наряду с предками ямной (Steppe_EMBA). В исследовании также было отмечено, что образцы древней Южной Азии имели значительно более высокий уровень Steppe_MLBA, чем Steppe_EMBA (или Ямная). Исследование также показало, что Rors может использоваться как прокси для ANI.[71]
Дэвид Райх в его книге 2018 года Кто мы и как мы сюда попали заявляет, что анализ 2016 года показал, что в ASI значительная часть наследственного компонента происходит от иранских фермеров (около 25% их предков), а остальные 75% их предков происходят от коренных южноазиатских охотников-собирателей. Он добавляет, что ASI вряд ли были местными охотниками-собирателями Южной Азии, как было установлено ранее, но населением, ответственным за распространение сельского хозяйства по всей Южной Азии. В случае с ANI, иранские фермерские предки составляют 50%, а остальные - из степных групп, связанных с Ямной.[72]
Нарасимхан и др. (2018) Таким же образом можно сделать вывод, что ANI и ASI были сформированы во 2-м тысячелетии до нашей эры.[73] Им предшествовала смесь AASI (древних исконных южноиндийских, то есть охотников-собирателей, имеющих общий корень с андаманцами); и иранские земледельцы, прибывшие в Индию ок. 4700–3000 гг. До н.э., и «должен был достичь долины Инда к 4-му тысячелетию до нашей эры».[73] Согласно Нарасимхану и др., Это смешанное население, которое, вероятно, происходило из цивилизации долины Инда, «внесло большой вклад как в ANI, так и в ASI», которые сформировались во 2-м тысячелетии до нашей эры. ANI сформирован из смеси "Инд Периферияродственные группы "и мигранты из степи, а АСИ сформировалась из"Инд Перифериясвязанных групп », которые двинулись на юг и смешались дальше с местными охотниками-собирателями. Предполагается, что предки населения ASI в среднем составляли около 73% от AASI и 27% от фермеров, связанных с Ираном. Нарасимхан и др. наблюдают, что образцы из группа периферии Инда всегда представляет собой смесь одних и тех же двух ближайших источников AASI и иранского земледельческого происхождения; «один из индейцев периферии имеет ~ 42% -ное происхождение AASI, а два других человека имеют ~ 14-18% -ное происхождение AASI» (с остальной частью их предков из иранского населения, связанного с земледелием).[73] Авторы обнаруживают, что коренные охотники-собиратели AASI представляют собой расходящуюся ветвь, которая отделилась примерно в то же время, когда предки восточноазиатских, онге (андаманских) и австралийских аборигенов отделились друг от друга. Он заключил, что «по существу вся родословная современных восточных и южных азиатов (до примеси южных азиатов, связанных с Западной Евразией) происходит от единственного распространения на восток, которое за короткий промежуток времени привело к линиям, ведущим к AASI. , Выходцы из Восточной Азии, онге и австралийцы.[73]
Генетическое исследование Yelmen et al. (2019) показывает, что современное население Южной Азии в целом ближе всего к западноевразийцам. Они пришли к выводу, что современные выходцы из Южной Азии в основном представляют собой смесь генетического компонента коренных жителей Южной Азии и более позднего западно-евразийского компонента (происходящего как из Западной Азии, так и из западных степей). Авторы также утверждают, что генетический компонент коренных жителей Южной Азии отличается от андаманцев, и что андаманцы, таким образом, являются несовершенным представителем. Этот компонент (представленный Андаманскими Онге) не был обнаружен в северной части Индии. Гуджарати, и поэтому предполагается, что южноиндийские племена Пания люди (группа преимущественно предков АСИ) послужили бы лучшим представителем, чем андаманцы (онге) для «коренного южноазиатского» компонента в современных южноазиатских аборигенах, поскольку пания происходят непосредственно от коренных жителей Южной Азии (а не из отдаленных районов). относятся к ним как Онге).[9]
Два генетических исследования (Shinde et al.2019 и Narasimhan et al.2019), в которых анализировались останки цивилизации долины Инда (части Северо-Западной Индии и Восточного Пакистана бронзового века), показали, что у них смешанная родословная: Shinde et al. обнаружили, что их образцы содержат около 50-98% генома народов, связанных с ранними иранскими фермерами, и 2-50% генома коренных южноазиатских охотников-собирателей, имеющих общую родословную с андаманцами, с иранскими родственными. родословная в среднем преобладает. Образцы, проанализированные Narasimhan et al. 45–82% имели родословную, связанную с иранскими фермерами, и 11–50% AASI (или андаманскую родословную, связанную с охотниками и собирателями). Анализируемые образцы обоих исследований практически не имеют "Степное происхождение «компонент, связанный с более поздними индоевропейскими миграциями в Индию. Авторы обнаружили, что соответствующее количество этих предков значительно различается между людьми, и пришли к выводу, что необходимо больше образцов, чтобы получить полную картину истории индийского населения.[74][75]
Генетическая дистанция между кастовыми группами и племенами
Исследования Watkins et al. (2005) и Kivisild et al. (2003) на основе аутосомных маркеров пришли к выводу, что индейские кастовые и племенные популяции имеют общее происхождение.[33][76] Редди и др. (2005) обнаружили довольно однородное распределение частот аллелей среди кастовых групп южных Андхра-Прадеш, но значительно большее генетическое расстояние между кастовыми группами и племенами указывает на генетическая изоляция племен и каст.[77]
Viswanathan et al. (2004) в исследовании генетической структуры и родства между племенными популяциями южной Индии заключает: «Генетическая дифференциация была высокой, и генетические расстояния не имели значительной корреляции с географическими расстояниями. Следовательно, генетический дрейф, вероятно, сыграл значительную роль в формировании паттернов генетической изменчивости, наблюдаемой в южноиндийских племенных популяциях. В остальном, анализ взаимоотношений популяций показал, что все индейские и южноазиатские популяции по-прежнему похожи друг на друга, независимо от фенотипических характеристик, и не обнаруживают какого-либо особого сходства с африканцами. Мы заключаем, что фенотипическое сходство некоторых индийских групп с африканцами не отражают тесные отношения между этими группами, но лучше объясняются конвергенция."[78]
Исследование 2011 г., опубликованное в Американский журнал генетики человека[5] указывает на то, что индейские предковые компоненты являются результатом более сложной демографической истории, чем считалось ранее. По мнению исследователей, Южная Азия имеет два основных наследственных компонента, один из которых распространяется с сопоставимой частотой и генетическим разнообразием в популяциях Центральной Азии, Западной Азии и Европы; другой компонент более ограничен Южной Азией. Однако, если исключить возможность крупномасштабной индоарийской миграции, эти результаты предполагают, что генетическая близость обоих индейских предковых компонентов является результатом множественных потоков генов на протяжении тысяч лет.[5]
Генетическое исследование, опубликованное в 2020 году в журнале ScienceDirect по Rowold et al. обнаружил, что подавляющее большинство Индийский народ («кастовые люди»), составляющие около 91,4%, тесно связаны с западноевразийцами (Европейцы и Жители Ближнего Востока ). Северные африканцы (выборка Египтяне ) были частью этого кластера, но смещены в сторону Африканцы к югу от Сахары. Выходцы из Восточной Азии и Выходцы из Юго-Восточной Азии были наиболее далеки как от жителей Западной Евразии, так и от жителей Африки к югу от Сахары и образовывали совершенно отдельный кластер. Интересно, что некастовые популяции Индии (выборка Солига люди, один из Адиваси ), которые составляют около 8,6% населения Индии, были смещены в сторону африканцев к югу от Сахары и имеют определенные аллели. Также была обнаружена связь с определенными обитателями Океании, что свидетельствует о древнем геноме.[79]
Более того, наличие этих Y-STR-профилей в нескольких популяциях к югу от Сахары и явное отсутствие в других евразийских коллекциях предполагает уникальную генетическую связь между индейскими племенными группами и африканцами к югу от Сахары.
— Rowold et al. 2020 г.
Смотрите также
- Археогенетика
- Этнические группы Южной Азии
- Список этнолингвистических регионов Южной Азии
- Население Индии
- Гаплогруппы Y-ДНК в популяциях Южной Азии
- Генетические исследования гуджарати
Рекомендации
- ^ а б c d е Кивисилд Т, Калдма К., Мецпалу М., Парик Дж., Папиха С., Виллемс Р. (1999). «Место вариантов митохондриальной ДНК Индии в глобальной сети материнских линий и населения Старого Света». Геномное разнообразие. С. 135–152. Дои:10.1007/978-1-4615-4263-6_11. ISBN 978-1-4613-6914-1.CS1 maint: ref = harv (связь)
- ^ а б Баиг М.М., Хан А.А., Кулькарни К.М. (сентябрь 2004 г.). «Разнообразие митохондриальной ДНК в племенных и кастовых группах Махараштры (Индия) и его влияние на их генетическое происхождение». Анналы генетики человека. 68 (Pt 5): 453–60. Дои:10.1046 / j.1529-8817.2004.00108.x. PMID 15469422. S2CID 23032872.
- ^ а б Сингх AK (2007). Наука и технологии для Upsc. Тата Макгроу-Хилл Образование. п. 595. ISBN 978-0-07-065548-5. В архиве из оригинала от 3 января 2014 г.. Получено 24 мая 2016.
- ^ а б c d Трипатия, Викал; Nirmala, A .; Редди, Б. Мохан (4 сентября 2017 г.). «Тенденции молекулярных антропологических исследований в Индии». Международный журнал генетики человека. 8 (1–2): 1–20. Дои:10.1080/09723757.2008.11886015. S2CID 12763485.
- ^ а б c Мецпалу М., Ромеро И.Г., Юнусбаев Б., Чаубей Г., Маллик С.Б., Худжашов Г. и др. (Декабрь 2011 г.). «Общие и уникальные компоненты структуры человеческой популяции и общегеномные сигналы позитивного отбора в Южной Азии». Американский журнал генетики человека. 89 (6): 731–44. Дои:10.1016 / j.ajhg.2011.11.010. ЧВК 3234374. PMID 22152676.
- ^ а б Мурджани П., Тангарадж К., Паттерсон Н., Липсон М., Ло П.Р., Говиндарадж П. и др. (Сентябрь 2013). «Генетические данные о недавнем смешении населения в Индии». Американский журнал генетики человека. 93 (3): 422–38. Дои:10.1016 / j.ajhg.2013.07.006. ЧВК 3769933. PMID 23932107.
- ^ Сильва М., Оливейра М., Виейра Д., Брандао А., Рито Т., Перейра Дж. Б. и др. (Март 2017 г.). «Генетическая хронология Индийского субконтинента указывает на сильное расселение по признаку пола». BMC Эволюционная биология. 17 (1): 88. Дои:10.1186 / s12862-017-0936-9. ЧВК 5364613. PMID 28335724.
- ^ а б Басу А., Саркар-Рой Н., Маджумдер П.П. (февраль 2016 г.). «Геномная реконструкция истории сохранившихся популяций Индии выявляет пять различных наследственных компонентов и сложную структуру». Труды Национальной академии наук Соединенных Штатов Америки. 113 (6): 1594–9. Bibcode:2016PNAS..113.1594B. Дои:10.1073 / pnas.1513197113. ЧВК 4760789. PMID 26811443.
- ^ а б Ельмень, Бурак; Мондаль, Маюх; Марнетто, Давиде; Патхак, Аджай К .; Монтинаро, Франческо; Гальего Ромеро, Ирен; Кивисилд, Тоомас; Мецпалу, Майт; Пагани, Лука (1 августа 2019 г.). «Анализ предков выявил различия в демографических историях и противоположное избирательное давление в современных популяциях Южной Азии». Молекулярная биология и эволюция. 36 (8): 1628–1642. Дои:10.1093 / molbev / msz037. ISSN 0737-4038. ЧВК 6657728. PMID 30952160.
- ^ а б c d Чаубей, Гианешвер (январь 2015 г.). «Восточноазиатское происхождение в Индии» (PDF). Индийский журнал физической антропологии и генетики человека. 34 (2): 193–199.
Здесь анализ полногеномных данных по Индии и Восточной / Юго-Восточной Азии продемонстрировал их ограниченное самобытное происхождение в Индии, в основном, вдоль предгорья Гималаев и северо-восточной части.
- ^ Чаубей, Гианешвер; Кадиан, Анураг; Бала, Сародж; Рао, Вадламуди Рагхавендра (10 июня 2015 г.). «Генетическая близость Бхила, Кола и Гонда, упомянутая в эпической Рамаяне». PLOS ONE. 10 (6): e0127655. Bibcode:2015PLoSO..1027655C. Дои:10.1371 / journal.pone.0127655. ISSN 1932-6203. ЧВК 4465503. PMID 26061398. S2CID 848806.
- ^ а б c d е Кивисилд Т, Папиха С.С., Рутси С., Парик Дж., Калдма К., Рейдла М. и др. (2000), Индийское происхождение: ключ к пониманию человеческого разнообразия в Европе и за ее пределами (PDF), Монографии Института Макдональда, в архиве (PDF) из оригинала 19 февраля 2006 г., получено 11 ноября 2005
- ^ а б c «Y гаплогрупп мира, 2005, Макдональдс» (PDF). Архивировано из оригинал (PDF) 28 июля 2004 г.. Получено 24 октября 2006.
- ^ а б c d е ж грамм час я j k л м п Сенгупта С., Животовский Л.А., Кинг Р., Мехди С.К., Эдмондс С.А., Чоу С.Э. и др. (Февраль 2006 г.). «Полярность и временность распределений Y-хромосомы высокого разрешения в Индии позволяют идентифицировать как местные, так и экзогенные экспансии и выявить незначительное генетическое влияние среднеазиатских скотоводов». Американский журнал генетики человека. 78 (2): 202–21. Дои:10.1086/499411. ЧВК 1380230. PMID 16400607.
- ^ а б c d е ж грамм час Саху С., Сингх А., Химабинду Дж., Банерджи Дж., Ситалаксими Т., Гайквад С. и др. (Январь 2006 г.). «Предыстория индийских Y-хромосом: оценка сценариев демической диффузии». Труды Национальной академии наук Соединенных Штатов Америки. 103 (4): 843–8. Bibcode:2006ПНАС..103..843С. Дои:10.1073 / pnas.0507714103. ЧВК 1347984. PMID 16415161.
- ^ а б c d е ж Thanseem I, Thangaraj K, Chaubey G, Singh VK, Bhaskar LV, Reddy BM и др. (Август 2006 г.). «Генетическое родство среди низших каст и племенных групп Индии: вывод из Y-хромосомы и митохондриальной ДНК». BMC Genetics. 7: 42. Дои:10.1186/1471-2156-7-42. ЧВК 1569435. PMID 16893451.
- ^ а б c Чжао З., Хан Ф., Боркар М., Эррера Р., Агравал С. (2009). «Наличие трех разных отцовских линий среди северных индейцев: исследование 560 Y-хромосом». Анналы биологии человека. 36 (1): 46–59. Дои:10.1080/03014460802558522. ЧВК 2755252. PMID 19058044.
- ^ Эндикотт, Мецпалу и Кивисилд 2007, п. 231.
- ^ Эндикотт, Мецпалу и Кивисилд 2007 С. 234–235.
- ^ Дас, Ранаджит; Упадхьяи, Приянка (25 ноября 2016 г.). «Отслеживание биогеографического происхождения населения Южной Азии с использованием ДНК SatNav». bioRxiv: 089466. Дои:10.1101/089466. S2CID 88966532.
Наша гипотеза подтверждается археологическими, лингвистическими и генетическими свидетельствами, которые позволяют предположить, что в Индию были две заметные волны иммиграции. Большинство ранних кавказоидов были носителями протодравидийского языка, которые мигрировали в Индию примерно на 6000 лет назад.
- ^ Оппенгеймер 2003[страница нужна ]
- ^ а б Пуэнте XS, Веласко Дж., Гутьеррес-Фернандес А., Бертранпетит Дж., Кинг М.С., Лопес-Отин С. (январь 2006 г.). «Сравнительный анализ генов рака в геномах человека и шимпанзе». BMC Genomics. 7: 15. Дои:10.1186/1471-2164-7-15. ЧВК 1382208. PMID 16438707.
- ^ а б c Мецпалу М., Кивисилд Т., Мецпалу Э., Парик Дж., Худжашов Г., Калдма К. и др. (Август 2004 г.). «Большинство существующих границ мтДНК в южной и юго-западной Азии, вероятно, были сформированы во время первоначального заселения Евразии анатомически современными людьми». BMC Genetics. 5: 26. Дои:10.1186/1471-2156-5-26. ЧВК 516768. PMID 15339343.
- ^ Кивисилд, Т .; Bamshad, M.J .; Kaldma, K .; Мецпалу, М .; Мецпалу, Э .; Reidla, M .; Лаос, S .; Parik, J .; Watkins, W.S .; Dixon, M.E .; Papiha, S.S .; Mastana, S.S .; Мир, М.Р .; Ferak, V .; Виллемс, Р. (ноябрь 1999 г.). «Глубокое общее происхождение индийских и западно-евразийских митохондриальных линий ДНК». Текущая биология. 9 (22): 1331–1334. Дои:10.1016 / s0960-9822 (00) 80057-3. PMID 10574762. S2CID 2821966.
- ^ а б «Гаплогруппа H Y-ДНК и ее субклады - 2015». В архиве из оригинала на 1 ноября 2015 г.. Получено 11 октября 2015.
- ^ а б c d е ж Cordaux R, Aunger R, Bentley G, Nasidze I, Sirajuddin SM, Stoneking M (февраль 2004 г.). «Независимое происхождение индейской касты и племенных отцовских линий». Текущая биология. 14 (3): 231–5. Дои:10.1016 / j.cub.2004.01.024. PMID 14761656. S2CID 5721248.
- ^ а б c Сингх, Сакши; Сингх, Ашиш; Раджкумар, Раджа; Сампат Кумар, Катакам; Кадаркарай Сами, Суббурадж; Низамуддин, шейх; Сингх, Амита; Ахмед Шейх, Шахнаваз; Педдада, Видья; Ханна, Винни; Вирайа, Пандичелвам; Пандит, Аридаман; Чаубей, Гианешвер; Сингх, Лалджи; Тангарадж, Кумарасами (12 января 2016 г.). «Рассмотрение влияния неолитической демической диффузии на индийский пул Y-хромосомы через гаплогруппу J2-M172». Научные отчеты. 6 (1): 19157. Bibcode:2016НатСР ... 619157С. Дои:10.1038 / srep19157. ISSN 2045-2322. ЧВК 4709632. PMID 26754573.
- ^ Эррера, Рене Дж .; Гарсия-Бертран, Ральф (2018). ДНК предков, происхождение человека и миграции. Академическая пресса. п. 250. ISBN 978-0-12-804128-4.
- ^ Арункумар Г., Сориа-Эрнанц Д.Ф., Кавита В.Дж., Арун В.С., Шьяма А., Ашокан К.С. и др. (2012). «Дифференциация населения южноиндийских мужских линий коррелирует с расширением сельского хозяйства, предшествовавшим кастовой системе». PLOS ONE. 7 (11): e50269. Bibcode:2012PLoSO ... 750269A. Дои:10.1371 / journal.pone.0050269. ЧВК 3508930. PMID 23209694.
- ^ а б c d е Шарма С., Рай Э., Шарма П., Джена М., Сингх С., Дарвиши К. и др. (Январь 2009 г.). «Индийское происхождение отцовской гаплогруппы R1a1 * подтверждает автохтонное происхождение браминов и кастовой системы». Журнал генетики человека. 54 (1): 47–55. Дои:10.1038 / jhg.2008.2. PMID 19158816. S2CID 22162114.
- ^ а б c Камар Р., Аюб К., Мохюддин А., Хельгасон А., Мазар К., Мансур А. и др. (Май 2002 г.). «Вариации ДНК Y-хромосомы в Пакистане». Американский журнал генетики человека. 70 (5): 1107–24. Дои:10.1086/339929. ЧВК 447589. PMID 11898125.
- ^ Шах AM, Таманг Р., Мурджани П., Рани Д.С., Говиндарадж П., Кулкарни Г. и др. (Июль 2011 г.). «Индийские сидди: потомки африканцев с примесью индейцев». Американский журнал генетики человека. 89 (1): 154–61. Дои:10.1016 / j.ajhg.2011.05.030. ЧВК 3135801. PMID 21741027.
- ^ а б c d е ж грамм час я j k Кивисилд (2003)
- ^ а б c Пийпе, Йерун; де Вугт, Алекс; ван Овен, Маннис; Хеннеман, Питер; ван дер Гааг, Кристиан Дж .; Кайзер, Манфред; де Книжфф, Питер (21 марта 2013 г.). «Перекресток Индийского океана: генетическое происхождение человека и структура населения на Мальдивах». Американский журнал физической антропологии. 151 (1): 58–67. Дои:10.1002 / ajpa.22256. ISSN 0002-9483. ЧВК 3652038. PMID 23526367.
- ^ Уэллс, Спенсер (2007). Глубокое происхождение: внутри генографического проекта. Вашингтон, округ Колумбия: National Geographic. ISBN 978-1426201189.
- ^ а б Махал Д.Г., Мацукас И.Г. (20 сентября 2017 г.). «Разнообразие гаплогрупп Y-STR в популяции джат свидетельствует о нескольких различных древних источниках». Границы генетики. 8: 121. Дои:10.3389 / fgene.2017.00121. ЧВК 5611447. PMID 28979290.
- ^ Спенсер Уэллс (2003), Путешествие человека. Генетическая одиссея. Нью-Дели: Penguin Books India, стр. 167
- ^ Чжао З., Хан Ф., Боркар М., Эррера Р., Агравал С. (2009). «Наличие трех разных отцовских линий среди северных индейцев: исследование 560 Y-хромосом». Анналы биологии человека. 36 (1): 46–59. Дои:10.1080/03014460802558522. ЧВК 2755252. PMID 19058044.
- ^ МакЭлриви К., Кинтана-Мурси Л. (2005). «Перспектива популяционной генетики долины Инда с помощью маркеров, унаследованных от одного родителя». Анналы биологии человека. 32 (2): 154–62. Дои:10.1080/03014460500076223. PMID 16096211. S2CID 109014.
- ^ Тангарадж К., Найду Б.П., Кривелларо Ф., Таманг Р., Упадхьяй С., Шарма В.К. и др. (Декабрь 2010 г.). «Влияние естественных барьеров на формирование генетической структуры популяций Махараштры». PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO ... 515283T. Дои:10.1371 / journal.pone.0015283. ЧВК 3004917. PMID 21187967.
- ^ Тангарадж К., Найду Б.П., Кривелларо Ф., Таманг Р., Упадхьяй С., Шарма В.К. и др. (Декабрь 2010 г.). Кордо Р. (ред.). «Влияние естественных барьеров на формирование генетической структуры популяций Махараштры». PLOS ONE. 5 (12): e15283. Bibcode:2010PLoSO ... 515283T. Дои:10.1371 / journal.pone.0015283. ЧВК 3004917. PMID 21187967.
- ^ а б c d Басу А., Мукерджи Н., Рой С., Сенгупта С., Банерджи С., Чакраборти М. и др. (Октябрь 2003 г.). «Этническая Индия: геномный взгляд, с особым упором на население и структуру». Геномные исследования. 13 (10): 2277–90. Дои:10.1101 / гр. 1413403. ЧВК 403703. PMID 14525929.CS1 maint: ref = harv (связь)
- ^ а б c d Firasat S, Khaliq S, Mohyuddin A, Papaioannou M, Tyler-Smith C, Underhill PA, Ayub Q (январь 2007 г.). «Доказательства Y-хромосомы ограниченного вклада Греции в популяцию патанов в Пакистане». Европейский журнал генетики человека. 15 (1): 121–6. Дои:10.1038 / sj.ejhg.5201726. ЧВК 2588664. PMID 17047675.CS1 maint: ref = harv (связь)
- ^ Камар 2002.
- ^ Mcelreavey 2005.
- ^ а б Кивисилд Т., Рутси С., Мецпалу М., Мецпалу Е., Парик Дж., Калдма К., Усанга Е., Мастана С., Папиха С.С., Виллемс Р. (2003). «Генетика языка и земледелия в Индии» (PDF). В Bellwood P, Renfrew C (ред.). Изучение гипотезы фермерства / распространения языков. Институт археологических исследований Макдональда, Кембридж, Великобритания. С. 215–222.
- ^ а б c Sengupta et al. (2005)[требуется полная цитата ]
- ^ а б c d е Андерхилл П.А., Майрес Н.М., Рутси С., Мецпалу М., Животовский Л.А., Кинг Р.Дж. и др. (Апрель 2010 г.). «Разделение постледникового происхождения европейских и азиатских хромосом Y в гаплогруппе R1a». Европейский журнал генетики человека. 18 (4): 479–84. Дои:10.1038 / ejhg.2009.194. ЧВК 2987245. PMID 19888303.CS1 maint: ref = harv (связь)
- ^ Мирабал С., Регейро М., Каденас А.М., Кавалли-Сфорца Л.Л., Андерхилл П.А., Вербенко Д.А. и др. (Октябрь 2009 г.). «Распределение Y-хромосомы в геолингвистическом ландшафте Северо-Запада России». Европейский журнал генетики человека. 17 (10): 1260–73. Дои:10.1038 / ejhg.2009.6. ЧВК 2986641. PMID 19259129.
- ^ а б c Андерхилл 2014.
- ^ а б c Памяв 2012.
- ^ ArunkumarG 2012.
- ^ Fornarino et al. (2009)
- ^ Манукян, Жан-Грегуар (2006), "Синтез гаплогруппы R2 - 2006 В архиве 1 октября 2015 г. Wayback Machine."
- ^ Мондаль, Маюх; Бергстрём, Андерс; Сюэ, Яли; Калафель, Франсеск; Лааюни, Хафид; Казальс, Ферран; Majumder, Partha P .; Тайлер-Смит, Крис; Бертранпетит, Жауме (1 мая 2017 г.). «Последовательности Y-хромосомы разнообразных индийских популяций и предков андаманцев». Генетика человека. 136 (5): 499–510. Дои:10.1007 / s00439-017-1800-0. HDL:10230/34399. ISSN 1432-1203. PMID 28444560. S2CID 3725426.
- ^ а б Кумар В., Редди А. Н., Бабу Дж. П., Рао Т. Н., Лангстих Б. Т., Тангарадж К. и др. (Март 2007 г.). «Данные по Y-хромосоме предполагают общее отцовское наследие австро-азиатских популяций». BMC Эволюционная биология. 7: 47. Дои:10.1186/1471-2148-7-47. ЧВК 1851701. PMID 17389048.
- ^ Eaaswarkhanth M, Haque I, Ravesh Z, Romero IG, Meganathan PR, Dubey B и др. (Март 2010 г.). «Следы происхождения к югу от Сахары и Ближнего Востока в индийском мусульманском населении». Европейский журнал генетики человека. 18 (3): 354–63. Дои:10.1038 / ejhg.2009.168. ЧВК 2859343. PMID 19809480.
- ^ «Место индийских вариантов мтДНК в глобальной сети материнских линий и населения Старого Света» (PDF). В архиве (PDF) из оригинала 8 марта 2012 г.. Получено 28 сентября 2011.
- ^ "Этнологический отчет для индоевропейцев". Ethnologue.com. В архиве из оригинала 15 октября 2012 г.. Получено 24 ноября 2015.
- ^ Baldi P (1990). Методология языковых изменений и реконструкции. Вальтер де Грюйтер. п. 342. ISBN 978-3-11-011908-4.
- ^ Берлинг (2003) С. 174–178.
- ^ Брэдли (2012) отмечает: МК в более широком смысле, включая языки мунда в восточной части Южной Азии, также известен как австроазиатский.Языки и языковые семьи в Китае В архиве 30 апреля 2017 г. Wayback Machine
- ^ Бамшад М., Кивисилд Т., Уоткинс В.С., Диксон М.Э., Рикер С.Е., Рао Б.Б. и др. (Июнь 2001 г.). «Генетические данные о происхождении кастового населения Индии». Геномные исследования. 11 (6): 994–1004. Дои:10.1101 / гр GR-1733RR. ЧВК 311057. PMID 11381027.
- ^ Мукерджи Н., Небель А., Оппенгейм А., Маджумдер П. П. (декабрь 2001 г.). «Анализ полиморфизма Y-хромосомы с высоким разрешением позволяет выявить признаки перемещения населения из Центральной и Западной Азии в Индию». Журнал генетики. 80 (3): 125–35. Дои:10.1007 / BF02717908. PMID 11988631. S2CID 13267463.
- ^ Мондаль, Маюх; Бергстрём, Андерс; Сюэ, Яли; Калафель, Франсеск; Лааюни, Хафид; Казальс, Ферран; Majumder, Partha P .; Тайлер-Смит, Крис; Бертранпетит, Жауме (25 апреля 2017 г.). «Последовательности Y-хромосомы разнообразных индейских популяций и предков андаманцев». Генетика человека. 136 (5): 499–510. Дои:10.1007 / s00439-017-1800-0. HDL:10230/34399. PMID 28444560. S2CID 3725426.
- ^ Райх Д., Тангарадж К., Паттерсон Н., Прайс А.Л., Сингх Л. (сентябрь 2009 г.). «Реконструкция истории населения Индии». Природа. 461 (7263): 489–94. Bibcode:2009Натура.461..489р. Дои:10.1038 / природа08365. ЧВК 2842210. PMID 19779445.
- ^ Мурджани П., Тангарадж К., Паттерсон Н., Липсон М., Ло П.Р., Говиндарадж П. и др.(Сентябрь 2013). «Генетические данные о недавнем смешении населения в Индии». Американский журнал генетики человека. 93 (3): 422–38. Дои:10.1016 / j.ajhg.2013.07.006. ЧВК 3769933. PMID 23932107.
- ^ а б Чаубей, Гианешвер; Эндикотт, Филипп (июнь 2013 г.). «Жители Андаманских островов в региональном генетическом контексте: пересмотр свидетельств раннего заселения архипелага из Южной Азии». Человеческая биология. 85 (1–3): 153–172. Дои:10.3378/027.085.0307. PMID 24297224. S2CID 7774927.
- ^ Маджумдер П.П. (февраль 2010 г.). «Генетическая история человека в Южной Азии». Текущая биология. 20 (4): R184-7. Дои:10.1016 / j.cub.2009.11.053. PMID 20178765. S2CID 1490419.
- ^ Ошибка цитирования. См. Встроенный комментарий, как исправить.[требуется проверка ]
- ^ Патхак, Аджай К .; Кадиан, Анураг; Кушняревич Алена; Монтинаро, Франческо; Мондаль, Маюх; Онгаро, Линда; Сингх, Манвендра; Кумар, Прамод; Рай, Нирадж; Парик, Юри; Мецпалу, Эне; Рутси, Сиири; Пагани, Лука; Кивисилд, Тоомас; Мецпалу, Майт; Чаубей, Гианешвер; Виллемс, Ричард (декабрь 2018 г.). «Генетическое происхождение современного населения долины Инда из Северо-Западной Индии». Американский журнал генетики человека. 103 (6): 918–929. Дои:10.1016 / j.ajhg.2018.10.022. ЧВК 6288199. PMID 30526867.
- ^ Райх, Дэвид (2018). Кто мы такие и как мы сюда попали: древняя ДНК и новая наука о прошлом человечества. ОУП Оксфорд. ISBN 978-0-19-257040-6. Получено 2 марта 2020.
- ^ а б c d Нарасимхан и др. 2018 г., п. 15.
- ^ Шинде В., Нарасимхан В.М., Роланд Н., Маллик С., Мах М., Липсон М., Накацука Н., Адамски Н., Брумандхошбахт Н., Ферри М., Лоусон А. М., Мишель М., Оппенгеймер Дж., Стюардсон К., Джадхав Н., Ким Ю. Дж., Чаттерджи М. , Munshi A, Panyam A, Waghmare P, Yadav Y, Patel H, Kaushik A, Thangaraj K, Meyer M, Patterson N, Rai N, Reich D (сентябрь 2019 г.). «В геноме древнего Хараппа отсутствует происхождение от степных скотоводов или иранских фермеров». Клетка. 179 (3): 729–735.e10. Дои:10.1016 / j.cell.2019.08.048. ЧВК 6800651. PMID 31495572.
- ^ Нарасимхан В.М., Паттерсон Н., Мурджани П., Роланд Н., Бернардос Р., Маллик С. и др. (Сентябрь 2019 г.). «Формирование человеческих популяций в Южной и Центральной Азии». Наука. 365 (6457): eaat7487. Дои:10.1126 / science.aat7487. ЧВК 6822619. PMID 31488661.
- ^ Watkins WS, Prasad BV, Naidu JM, Rao BB, Bhanu BA, Ramachandran B. и др. (Ноябрь 2005 г.). «Разнообразие и расхождение среди племенных популяций Индии». Анналы генетики человека. 69 (Pt 6): 680–92. Дои:10.1046 / j.1529-8817.2005.00200.x. PMID 16266407. S2CID 31907598.
- ^ Редди Б.М., Найду В.М., Мадхави В.К., Тангарадж Л.К., Кумар В., Лангстих Б.Т. и др. (Декабрь 2005 г.). «Разнообразие микросателлитов в Андхра-Прадеш, Индия: генетическая стратификация против социальной стратификации». Человеческая биология. 77 (6): 803–23. Дои:10.1353 / ступица.2006.0018. PMID 16715839. S2CID 18446485.
- ^ Вишванатан Х., Дипа Э., Кордо Р., Стоункинг М., Уша Рани М.В., Маджумдер П.П. (март 2004 г.). «Генетическая структура и родство среди племенных популяций южной Индии: исследование 24 маркеров аутосомной ДНК». Анналы генетики человека. 68 (Pt 2): 128–38. Дои:10.1046 / j.1529-8817.2003.00083.x. PMID 15008792. S2CID 24230856.
- ^ "Y-хромосома солига, древнего лесного племени Южной Индии". Джин: X. 5: 100026. 1 декабря 2020 г. Дои:10.1016 / j.gene.2019.100026. ISSN 2590-1583.
дальнейшее чтение
- Алликас А., Орд Д., Кург Р., Киви С., Устав М. (июнь 2001 г.). «Роли шарнирной области и ДНК-связывающего домена белка E2 бычьего папилломавируса типа 1 в инициации репликации ДНК». Вирусные исследования. 75 (2): 95–106. Дои:10.1016 / S0168-1702 (01) 00219-2. PMID 11325464.CS1 maint: ref = harv (связь)
- Бехар Д.М., Гарриган Д., Каплан М.Э., Мобашер З., Розенгартен Д., Карафет TM и др. (Март 2004 г.). «Контрастные модели вариаций Y-хромосомы у евреев ашкенази и нееврейского европейского населения». Генетика человека. 114 (4): 354–65. Дои:10.1007 / s00439-003-1073-7. PMID 14740294. S2CID 10310338.
- Бхаттачарья Н.П., Басу П., Дас М., Праманик С., Банерджи Р., Рой Б. и др. (Август 1999 г.). «Незначительный поток мужских генов через этнические границы в Индии, выявленный анализом полиморфизма ДНК Y-хромосомы». Геномные исследования. 9 (8): 711–9. Дои:10.1101 / гр.9.8.711 (неактивно 25 сентября 2020 г.). PMID 10447506.CS1 maint: DOI неактивен по состоянию на сентябрь 2020 г. (связь)
- Канн Р.Л. (март 2001 г.). «Генетические ключи к расселению в человеческих популяциях: прослеживая прошлое из настоящего». Наука. 291 (5509): 1742–8. Bibcode:2001Sci ... 291.1742C. Дои:10.1126 / science.1058948. PMID 11249820. S2CID 19367408.CS1 maint: ref = harv (связь)
- Cinniolu C, King R, Kivisild T, Kalfolu E, Atasoy S, Cavalleri GL, et al. (Январь 2004 г.). «Раскопки гаплотипов Y-хромосомы в Анатолии». Генетика человека. 114 (2): 127–48. Дои:10.1007 / s00439-003-1031-4. PMID 14586639. S2CID 10763736.
- Дас Б., Чаухан П.С., Сешадри М. (октябрь 2004 г.). «Минимальное совместное использование STR-гаплотипов Y-хромосомы среди пяти эндогамных групп населения из западной и юго-западной Индии». Человеческая биология. 76 (5): 743–63. Дои:10.1353 / хаб.2005.0003. PMID 15757245. S2CID 12835244.
- Hemphill BE, Christensen AF (3 ноября 1994 г.). Цивилизация Оксуса как связующее звено между Востоком и Западом: неметрический анализ биологического сходства Бактрена бронзового века. Мэдисон, Висконсин. п. 13.CS1 maint: ref = harv (связь) (доклад прочитан на конференции в Южной Азии)
- Джоблинг М.А., Тайлер-Смит К. (август 2003 г.). «Человеческая Y-хромосома: эволюционный маркер совершеннолетия». Обзоры природы. Генетика. 4 (8): 598–612. Дои:10.1038 / nrg1124. PMID 12897772. S2CID 13508130.
- Кивисилд Т, Рутси С., Мецпалу М., Мастана С., Калдма К., Парик Дж. И др. (Февраль 2003 г.). «Генетическое наследие первых поселенцев сохраняется как в индейских племенах, так и в кастах». Американский журнал генетики человека. 72 (2): 313–32. Дои:10.1086/346068. ЧВК 379225. PMID 12536373.CS1 maint: ref = harv (связь)
- Мецпалу М., Кивисилд Т., Мецпалу Э., Парик Дж., Худжашов Г., Калдма К. и др. (Август 2004 г.). «Большинство существующих границ мтДНК в южной и юго-западной Азии, вероятно, были сформированы во время первоначального заселения Евразии анатомически современными людьми». BMC Genetics. 5: 26. Дои:10.1186/1471-2156-5-26. ЧВК 516768. PMID 15339343.
- Патовари А., Пурканти Р., Сингх М., Чаухан Р.К., Бхартия Д., Двиведи О.П. и др. (Июль 2012 г.). «Систематический анализ и функциональная аннотация вариаций в геноме индейцев». Человеческая мутация. 33 (7): 1133–40. Дои:10.1002 / humu.22091. PMID 22461382. S2CID 11466942.
- Рутси С., Магри С., Кивисилд Т., Бенуцци Дж., Хелп Х., Бермишева М. и др. (Июль 2004 г.). «Филогеография гаплогруппы I Y-хромосомы выявляет отдельные области доисторического потока генов в Европе». Американский журнал генетики человека. 75 (1): 128–37. Дои:10.1086/422196. ЧВК 1181996. PMID 15162323.
- Камар Р., Аюб К., Мохюддин А., Хельгасон А., Мазар К., Мансур А. и др. (Май 2002 г.). «Вариации ДНК Y-хромосомы в Пакистане». Американский журнал генетики человека. 70 (5): 1107–24. Дои:10.1086/339929. ЧВК 447589. PMID 11898125.
- Семино О, Магри С., Бенуцци Дж., Лин А.А., Аль-Захери Н., Батталья В. и др. (Май 2004 г.). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье». Американский журнал генетики человека. 74 (5): 1023–34. Дои:10.1086/386295. ЧВК 1181965. PMID 15069642.
Источники
- Индийский консорциум по изменению генома (апрель 2008 г.). «Генетический ландшафт народа Индии: холст для исследования генов болезней». Журнал генетики. 87 (1): 3–20. Дои:10.1007 / s12041-008-0002-x. PMID 18560169. S2CID 21473349.
- Эндикотт П., Мецпалу М., Кивисилд Т. (2007). «Генетические данные о современных расселениях людей в Южной Азии: перспективы Y-хромозы и митохондриальной ДНК». В Майкл Д. Петраглиа, Бриджит Оллчин (ред.). Эволюция и история человеческих популяций в Южной Азии. Springer. С. 201–228. ISBN 978-1-4020-5561-4.
- Хемфилл Б, Лукач Дж, Кеннеди К. (1991). «Биологические адаптации и сходства хараппцев бронзового века». В Meadow RH (ред.). Раскопки в Хараппе 1986–1990: мультидисциплинарный подход к урбанизму третьего тысячелетия. С. 137–82. ISBN 978-0-9629110-1-9.CS1 maint: ref = harv (связь)
- Кеннеди К.А. (1984). «Переоценка теорий расового происхождения людей цивилизации долины Инда на основе последних антропологических данных». В Kennedy KA, Possehl GL (ред.). Исследования в области археологии и палеоантропологии Южной Азии. Атлантическое нагорье, Нью-Джерси: Гуманитарная пресса. С. 99–107.CS1 maint: ref = harv (связь)
- Кеннеди К.А. (1995). «Были ли арийцы идентифицированы в доисторических скелетных записях из Южной Азии?». У Джорджа Эрдози (ред.). Индоарии древней Южной Азии. Вальтер де Грюйтер. С. 49–54. ISBN 978-3-11-014447-5.
- Кивисилд Т (2000б). Происхождение популяций южной и западной Евразии: исследование мтДНК (PDF). Тартуский университет, Эстония.CS1 maint: ref = harv (связь) (Кандидат наук)
- Маскареньяс Д.Д., Райна А., Астон CE, Сангера Д.К. (2015). «Генетическая и культурная реконструкция миграции древнего происхождения». BioMed Research International. 2015: 651415. Дои:10.1155/2015/651415. ЧВК 4605215. PMID 26491681.
- Нарасимхан В.М., Энтони Д., Мэллори Дж., Рейх Д. (2018). «Геномная формация Южной и Центральной Азии». bioRxiv 10.1101/292581. Дои:10.1101/292581. S2CID 89658279. Цитировать журнал требует
| журнал =(помощь) - Оппенгеймер С (2003). Настоящая Ева: Путешествие современного человека из Африки. Нью-Йорк: Издательство Кэрролл и Граф. ISBN 978-0-7867-1192-5.CS1 maint: ref = harv (связь)
- Памяв (декабрь 2012 г.), «Краткое сообщение: новые бинарные маркеры Y-хромосомы улучшают филогенетическое разрешение внутри гаплогруппы R1a1», Американский журнал физической антропологии, 149 (4): 611–615, Дои:10.1002 / ajpa.22167, PMID 23115110, S2CID 4820868
- Ренфрю С., Бойл К., ред. (2000а). Индийское происхождение: ключ к пониманию человеческого разнообразия в Европе и за ее пределами (PDF). ISBN 978-1-902937-08-3.CS1 maint: ref = harv (связь)
- Андерхилл PA (2003). «Вывод истории человечества: ключи от гаплотипов Y-хромосомы». Симпозиумы Колд-Спринг-Харбор по количественной биологии. 68: 487–93. Дои:10.1101 / sqb.2003.68.487. PMID 15338652. S2CID 15527457.
- Андерхилл П.А., Позник Г.Д., Рутси С., Ярве М., Лин А.А., Ван Дж. И др. (Январь 2015 г.). «Филогенетическая и географическая структура гаплогруппы Y-хромосомы R1a». Европейский журнал генетики человека. 23 (1): 124–31. Дои:10.1038 / ejhg.2014.50. ЧВК 4266736. PMID 24667786.
- Уэллс S (2003). Путешествие человека: генетическая одиссея. Издательство Принстонского университета.
внешняя ссылка
- Введение в гаплогруппы и гаплотипы, Марк А. Джоблинг, Лестерский университет. [1]
- Путешествие человека: Население мира, Фонд Брэдшоу, совместно с Стивен Оппенгеймер.
- Индийская база данных вариаций генома Институт геномики и интегративной биологии
- Список частот R2
- Мальдивы
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|
| |