Пернатый динозавр - Feathered dinosaur

А пернатый динозавр любой вид динозавр обладание перья. Хотя сюда входят все виды птицы, недавние данные подтверждают гипотезу о том, что многие, если не все виды динозавров, не являющиеся птицами, также обладали перьями той или иной формы. Было высказано предположение, что перья изначально развились для целей теплоизоляция, как и остается их целью в пуховые перья птенцов птиц сегодня, до их возможной модификации у птиц в структуры, поддерживающие полет.
Поскольку научные исследования динозавров начались в начале 1800-х годов, считалось, что они тесно связаны с современными рептилии, Такие как ящерицы. Само слово «динозавр», придуманное в 1842 году палеонтологом. Ричард Оуэн, происходит из Греческий для "грозной ящерицы". Эта точка зрения начала меняться во время так называемого ренессанс динозавров в научных исследованиях в конце 1960-х, а к середине 1990-х появились значительные доказательства того, что динозавры были гораздо более тесно связаны с птицы, которые произошли непосредственно от теропод группа динозавры[1] и сами являются подгруппой динозавров.
Знания о происхождении перьев появились в результате открытия новых окаменелостей в течение 2000-х и 2010-х годов, а также благодаря технологиям, которые позволили ученым более внимательно изучить окаменелости. Среди нептичьи динозавры, перья или похожий на перо покров были обнаружены в десятках роды через прямые и косвенные ископаемые свидетельства.[2] Хотя подавляющее большинство открытий перьев было сделано в целурозавровые тероподы перьевидные покровы также были обнаружены как минимум в трех орнитисхианы, предполагая, что перья могли присутствовать на последнем общем предке Орнитосцелида, а динозавр группа, включающая как тероподы и орнитисхианы.[3] Возможно, что перья впервые появились у еще более ранних архозавров, в свете открытия очень похожих на перья пикнофибры в птерозавры.[4] Крокодилы также обладают бета-кератин аналогично тем из птицы, что предполагает, что они произошли от общих предковых генов.[5]
История исследования
Рано

Вскоре после публикации 1859 г. Чарльз Дарвин с О происхождении видов, Британский биолог Томас Генри Хаксли предположил, что птицы были потомками динозавров. Он сравнил скелетную структуру Компсогнат, маленький теропод динозавр, и первый птица ' Литографический археоптерикс (оба были найдены в Верхняя юра Баварский известняк из Solnhofen ). Он показал, что, кроме его рук и перьев, Археоптерикс был очень похож на Компсогнат. Таким образом Археоптерикс представляет переходное ископаемое. В 1868 г. он опубликовал О животных, которые занимают промежуточное положение между птицами и рептилиями, делая дело.[6][7] Первая реставрация пернатого динозавра была Томаса Генри Хаксли изображение в 1876 г. пернатого Компсогнат чтобы сопровождать лекцию об эволюции птиц, которую он прочитал в Нью-Йорке, в которой он предположил, что вышеупомянутый динозавр мог обладать перьями.[8] Ведущий динозавр знаток времени, Ричард Оуэн, не согласен, утверждая Археоптерикс как первая птица вне линии динозавров. В течение следующего столетия утверждают, что птицы были динозавр потомков исчезли, появились более популярные гипотезы происхождения птиц, в том числе:крокодиломорф ' и 'Текодонт 'предки, а не динозавры или другие архозавры.
'Возрождение динозавров'
В 1969 г. Джон Остром описанный Deinonychus antirrhopus, а теропод который он обнаружил в Монтане в 1964 году, и чье скелетное сходство с птицами казалось безошибочным. Остром стал одним из ведущих сторонников теории, согласно которой птицы являются прямыми потомками динозавров. Дальнейшие сравнения скелетов птиц и динозавров, а также кладистический анализ усиленный аргумент для ссылки, особенно для ветви теропод, называемой манирапторы. Скелетные сходства включают шея, то лобок, то запястья (полулунный запястный ), 'руки ' и грудной пояс, то лопатка, то ключица и Грудина. В целом, птицы и динозавры-тероподы обладают более чем сотней различных анатомических особенностей.[нужна цитата ] Другие исследователи обратили внимание на эти общие черты и другие аспекты биологии динозавров и начали предполагать, что по крайней мере некоторые динозавры-теропод были пернатыми.
В то же время, палеохудожники начали создавать современные реставрации высокоактивных динозавров. В 1969 г. Роберт Т. Баккер привлек бег Дейноних. Его ученик Грегори С. Пол изображен нептичий манирапторан динозавры с перьями и прародителями, начиная с конца 1970-х годов. В 1975 г. Элеонора Киш начал рисовать точные изображения динозавров, ее Гипакрозавр быть первым показанным с его камуфляж.[9]
До открытия окаменелостей пернатых динозавров доказательства были ограничены исследованиями Хаксли и Острома. сравнительная анатомия. Некоторые основные орнитологи, включая Смитсоновский институт куратор Сторрс Л. Олсон, оспорили ссылки, в частности, сославшись на отсутствие окаменелостей пернатых динозавров.[нужна цитата ] Однако к 1990-м годам большинство палеонтологов считали птиц выжившими динозаврами и относили их к `` нептичьим динозаврам '' (все вымершие), чтобы отличить их от птиц (Avialae ).
Открытия окаменелостей
Первый известный образец Археоптерикс, на основании которого был назван род, представлял собой изолированное перо, независимо от того, принадлежит ли оно к Археоптерикс был спорным.[10][11] Одним из самых ранних открытий возможных отпечатков перьев у нептичьих динозавров является ихнофоссил (Fulicopus lyellii ) возрастом 195-199 млн лет Формация Портленда на северо-востоке США. Герлински (1996, 1997, 1998) и Кундрат (2004) интерпретировали следы между двумя следами на этой окаменелости как отпечатки перьев на животе сидящего на корточках. дилофозаврид. Хотя некоторые обозреватели подняли вопросы о названии и интерпретации этой окаменелости, если это так, то это раннее Юрский период окаменелость - самое старое известное свидетельство существования перьев, оно почти на 30 миллионов лет старше следующего по возрасту из известных свидетельств.[12][13][14]

После столетия гипотез без убедительных доказательств в течение 1990-х годов были обнаружены хорошо сохранившиеся окаменелости пернатых динозавров, и их продолжают находить. Окаменелости сохранились в Lagerstätte —А осадочный месторождение, демонстрирующее удивительное богатство и полноту окаменелостей - в Ляонин, Китай. Район неоднократно задыхался вулканическим пеплом, образовавшимся в результате извержений в Внутренняя Монголия 124 миллиона лет назад, во время Раннемеловой период эпоха. Мелкозернистый пепел в мельчайших деталях сохранил погребенные живые организмы. Местность кипела жизнью, с миллионами листьев, покрытосеменные (самый старый из известных), насекомые, рыбы, лягушки, саламандры, млекопитающие, черепахи, и ящерицы обнаружено на сегодняшний день.
Самыми важными открытиями в Ляонине стали окаменелости пернатых динозавров с постоянным потоком новых находок, дополняющих картину связи между динозаврами и птицами и дополняющих теории эволюционного развития перьев и полета. Тернер и другие. (2007) сообщили о перьях на локтевой кости Велоцираптор монгольский, и они сильно коррелируют с крупными и хорошо развитыми вторичными перьями.[15]

Поведенческие доказательства в виде овирапторозавр на своем гнезде показал еще одну связь с птицами. Его предплечья были сложены, как у птицы.[16] Хотя нет перья были сохранены, вероятно, они присутствовали для изоляции яиц и молоди.[17]

Однако не все китайские открытия окаменелостей оказались достоверными. В 1999 году предполагаемая окаменелость предположительно пернатого динозавра по имени Археораптор liaoningensis, нашел в Провинция Ляонин, Северо-Восточный Китай, оказался подделкой. Сравнивая фотографию экземпляра с другой находкой, китайский палеонтолог Сюй Син пришли к выводу, что он состоит из двух частей разных ископаемых животных. Его требование сделано Национальная география просмотрите свои исследования, и они тоже пришли к такому же выводу.[18] Нижняя часть "Археораптор "композит произошел от законного пернатого дромеозаврида, теперь известного как Микрораптор, а верхняя часть от ранее известной примитивной птицы по имени Янорнис.
В 2011 г. образцы Янтарь были обнаружены сохранившиеся перья от 75 до 80 миллионов лет назад во время Меловой эпохи, с доказательствами, что они были от динозавров и птиц. Первоначальный анализ предполагает, что некоторые перья использовались для изоляции, а не для полета.[19][20] Было обнаружено, что более сложные перья имеют вариации окраски, похожие на окраску современных птиц, в то время как более простые перья были преимущественно темными. В настоящее время известно всего 11 экземпляров. Образцы слишком редки, чтобы их можно было вскрыть, чтобы изучить их. меланосомы, но есть планы использовать неразрушающие рентгеновские снимки с высоким разрешением.[21] Меланосомы - это клетки, содержащие пигмент, которые окрашивают перья; Поскольку меланосомы разной формы дают разные цвета, последующие исследования меланосом, сохранившихся в образцах пернатых динозавров, привели к реконструкции облик нескольких видов динозавров. К ним относятся Анхиорнис,[22] Синозауроптерикс,[23] Микрораптор,[24] и Археоптерикс.[11]
В 2016 году было объявлено об открытии хвоста пернатого динозавра, сохранившегося в янтаре, возраст которого оценивается в 99 миллионов лет. Лида Син, исследователь из Китайский университет наук о Земле в Пекин, нашел образец на янтарном рынке в Мьянма. Это первое окончательное открытие материала динозавров в Янтарь.[25][26][27][28]
Текущие знания
Сохранились нептичьи виды динозавров со следами перьев




Сейчас известно, что у нескольких динозавров, не являющихся птицами, были перья. Прямые доказательства наличия перьев существуют у нескольких видов.. Во всех примерах описанные доказательства состоят из отпечатков перьев, за исключением тех родов, которые, как предполагается, имели перья на основании скелетных или химических доказательств, таких как наличие гусиных шишек (точки привязки перьев крыльев на передних конечностях) или пигостиль (сросшиеся позвонки на конце хвоста, которые часто поддерживают большие перья).[29]
Примитивные типы перьев
Покровные структуры, из которых выросли перья птиц, видны на спинных шипах рептилий и рыб. Подобный этап их эволюции со сложной оболочкой птиц и млекопитающих можно наблюдать у живых рептилий, таких как игуаны и Гоноцефал агамиды. Считается, что перьевые структуры произошли от простых полых нитей через несколько стадий возрастающей сложности, заканчивая большими, глубоко укоренившимися перьями с прочными перьями (рахис ), зазубрины и бородки, которые птицы показывают сегодня.[30]
Согласно модели, предложенной Prum (1999), на стадии I фолликул образует цилиндрическое эпидермальное углубление вокруг основания перьевого сосочка. Первое перо появилось, когда недифференцированный трубчатый воротник фолликула развился из старых кератиноцитов, выталкиваемых наружу. На стадии II внутренний, базилярный слой воротничка фолликула дифференцировался на продольные гребешки с неразветвленными кератиновыми волокнами, а тонкий периферический слой воротника стал листопадным влагалищем, образовав пучок неразветвленных зубцов с базальным аиром. Стадия III состоит из двух нововведений в развитии, IIIa и IIIb, каждая из которых могла произойти первой. Стадия IIIa включает в себя спиральное смещение зубчатых гребней, возникающих внутри воротника. Зубчатые гребни на передней средней линии фолликула сливаются вместе, образуя рахис. Далее следует создание заднего локуса заусениц, что дает неопределенное количество зазубрин. В результате получилось перо с симметричной, в первую очередь разветвленной структурой с рахисом и неразветвленными зазубринами. На стадии IIIb бородки, спаренные внутри периферийных бородковых пластин гребешков, образуют разветвленные бородки с ветвями и бородками. В результате получается перо с пучком разветвленных зубцов без рахиса. На стадии IV дифференцированные дистальные и проксимальные бородки образуют замкнутую пластинчатую лопатку (a контурное перо ). Закрытая лопасть образуется, когда гребешки на дистальных бородках образуют крючковатую форму для прикрепления к более простым проксимальным бородкам соседней бородки. Нововведения в развитии стадии V привели к дополнительному структурному разнообразию в замкнутом перистом перо. Здесь несимметричные маховые перья, двулепестковые перистый перья[требуется разъяснение ] филоплюмы,[требуется разъяснение ] порошок вниз,[требуется разъяснение ] и щетина[требуется разъяснение ] эволюционировал.[31]
Некоторые данные свидетельствуют о том, что первоначальной функцией простых перьев была изоляция. В частности, сохранившиеся участки кожи на крупных, производных, тираннозавроиды Показать щитки а у более мелких и примитивных - перья. Это может указывать на то, что у более крупных форм была сложная кожа с щитками и нитями, или что тираннозавроиды может быть как носороги и слоны, имея нити при рождении, а затем теряя их по мере развития до зрелости.[32] Взрослый тиранозавр Рекс весил примерно столько же Африканский слон. Если большой тираннозавроиды мы эндотермы, они должны были бы эффективно излучать тепло.[33] Однако из-за различных структурных свойств перьев по сравнению с мехом,[34] а также большей площади поверхности на кубический метр, это крайне маловероятно даже для самых больших тероподы будет страдать от перегрева из-за обширной перьевой шерсти.[нужна цитата ]
Некоторые данные также свидетельствуют о том, что более производные типы перьев могли служить целям изоляции. Например, исследование Овираптор перья на крыльях и положение гнезд позволяют предположить, что удлиненные перья на крыльях, очевидно, могли служить для заполнения промежутков между изоляционной камерой тела насиженных особей и внешней средой. Эта «стена» из перьев крыльев могла защитить яйца от перепадов температур.[35]
Появляется все больше доказательств, подтверждающих гипотезу демонстрации, которая утверждает, что ранние перья были окрашены и повысили репродуктивный успех.[36][37] Окраска могла обеспечить первоначальную адаптацию перьев, подразумевая, что все последующие функции перьев, такие как терморегуляция и полет, были кооптированный.[36] Эта гипотеза была подтверждена открытием пигментированных перьев у многих видов.[38][39][40] [41] Гипотезу о демонстрации поддерживает тот факт, что ископаемые перья наблюдались в кладе наземных травоядных динозавров, поэтому маловероятно, что перья использовались как хищные орудия или средство полета.[42] Кроме того, у некоторых экземпляров перья радужные.[43] Пигментированные и переливающиеся перья, возможно, обеспечивали большую привлекательность для самок, обеспечивая более высокий репродуктивный успех по сравнению с неокрашенными перьями.[44] Текущие исследования показывают, что вполне вероятно, что у теропод была острота зрения, необходимая для того, чтобы видеть изображения. В исследовании Стивенса (2006) бинокулярное поле зрения для Велоцираптор по оценкам, от 55 до 60 градусов, что примерно соответствует температуре современных сов. Острота зрения для Тиранозавр было предсказано, что оно будет примерно от человеческого до 13 раз больше, чем у людей.[45] Однако, поскольку оба Велоцираптор и Тиранозавр имеют довольно обширные эволюционные отношения с более базальными тероподами, неясно, какая часть этих данных по остроте зрения может быть экстраполирована.[нужна цитата ]
Идея о том, что предшественники перьев появились до того, как они были использованы для изоляции, уже высказана в Gould and Vrba, 1982.[46] Первоначальная польза могла быть метаболической. Перья в основном состоят из кератинового белкового комплекса, который имеет дисульфидные связи между аминокислотами, которые придают ему стабильность и эластичность. Метаболизм аминокислот, содержащих серу, может быть токсичным; однако, если серные аминокислоты не катаболизируются в конечных продуктах мочевины или мочевой кислоты, а вместо этого используются для синтеза кератина, высвобождение сероводорода значительно снижается или предотвращается. Для организма, метаболизм которого происходит при высоких внутренних температурах 40 ° C или выше, может быть чрезвычайно важно предотвратить избыточное производство сероводорода. Эта гипотеза может быть совместима с необходимостью высокой скорости метаболизма тероподных динозавров.[47][48]
Точно неизвестно, в какой момент архозавр филогения самые ранние простые «праотцы» возникали либо один раз, либо независимо несколько раз. Нитчатые структуры явно присутствуют в птерозавры,[49] и длинные, полые иглы были зарегистрированы в образцах орнитисхий динозавры Пситтакозавр и Тяньюлонг.[50][51] В 2009 году Xu et al. отметил, что полые, неразветвленные, жесткие покровные структуры, обнаруженные на образце Бейпиаозавр были поразительно похожи на покровные структуры Пситтакозавр и птерозавры. Они предположили, что все эти структуры могли быть унаследованы от общего предка намного раньше в эволюции архозавры, возможно, в орнитодир из Среднего Триасовый или раньше.[52] Совсем недавно находки в России базальных неорнитишиан Кулиндадромей сообщают, что, хотя голень и хвост казались покрытыми чешуей, «различные покровные структуры были обнаружены, непосредственно связанные с элементами скелета, что подтверждает гипотезу о том, что простые нитевидные перья, а также сложные перьевидные структуры, сопоставимые с таковыми у теропод, были широко распространены среди людей. вся клады динозавров ".[53]
Отображаемые перья также известны от динозавров, которые являются очень примитивными представителями рода птиц, или Avialae. Самый примитивный пример: Эпидексиптерикс, у которого был короткий хвост с очень длинными лентообразными перьями. Как ни странно, окаменелость не сохраняет перья крыльев, что позволяет предположить, что Эпидексиптерикс был либо вторично нелетающим, либо этот показ перьев развился раньше летных перьев в птичьей линии.[54] Плюшевые перья встречаются почти во всех линиях Тероподы распространены в северном полушарии, а перистые перья засвидетельствованы так далеко вниз по дереву, как Орнитомимозаврия. Дело в том, что только взрослый Орнитомим имели похожие на крылья структуры, что говорит о том, что перья из меловых перьев эволюционировали для спаривания.[55]
Филогения и вывод перьев у других динозавров
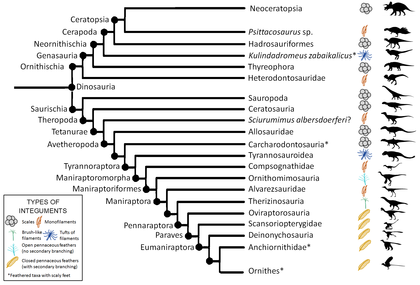
Ископаемые отпечатки перьев крайне редки, и для их формирования требуются исключительные условия хранения. Таким образом, только несколько нептичьих пернатых динозавров роды были идентифицированы. Было обнаружено, что все ископаемые образцы пера демонстрируют определенное сходство. Из-за этого сходства и благодаря исследованиям развития многие ученые считают, что перья у динозавров эволюционировали только один раз.[нужна цитата ] Затем перья передавались бы всем более поздним, более производным видам, если бы некоторые линии не потеряли перья вторично. Если динозавр попадает в точку эволюционного древа в пределах известных перьевых родословных, значит, у его предков были перья, и вполне возможно, что они тоже были.[нужна цитата ] Эта техника, называемая филогенетический брекетинг, также можно использовать для определения типа перьев, которые могли иметь вид, поскольку история развития перьев в настоящее время достаточно хорошо известна. У всех видов пернатых были нитевидные или перистые (пуховые) перья, при этом перья с мелкими перьями встречаются у более похожих на птиц групп. Следующее кладограмма адаптировано из Godefroit и другие., 2013.[56]
Филогенетический брекетинг также может использоваться для доказательства отсутствия пернатых покровов путем вывода. Например, наличие чешуйчатых покровов в определенной кладе будет сильным показателем того, что члены этой клады будут иметь одинаковые покровы, поскольку независимая эволюция перьев несколько раз маловероятна, независимо от того, присутствуют ли ископаемые свидетельства для всех родов в кладе.
Серый обозначает кладу, о которой на момент написания не было известно ни одного экземпляра с перьями, некоторые из которых имеют ископаемые остатки чешуи. Присутствие или отсутствие пернатых экземпляров в данной кладе не подтверждает, что все члены в кладе имеют указанные покровы, если только не подтверждено репрезентативными ископаемыми свидетельствами членов этой клады.
| Neotheropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Следующая кладограмма сделана Сюй (2020).[57]
- Тонкий монофиламентный покров
- Широкий монофиламентный покров
- Основное соединение нитчатого пера
- Основное соединение стержня нитевидного пера
- Нитевидное перо с радиально разветвленным стержнем
- Двусторонне разветвленное нитчатое перо
- Основное соединение разветвленного нитчатого пера
- Основное соединение нитчатого пера на основе перепончатой
- Симметричное перо с открытыми лопастями
- Симметричное перо с закрытыми лопастями
- Асимметричное перо с закрытыми лопастями
- Проксимально ленточное перо с закрытыми лопастями
- Рахис-доминантное перо с закрытыми лопастями
| Орнитодира |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Смотрите также
Рекомендации
- ^ Браун, Джозеф В .; Ван Туйнен, М. (2011). «Эволюция представлений о древности современного птичьего дерева». Живые динозавры. С. 306–324. Дои:10.1002 / 9781119990475.ch12. ISBN 9781119990475.
- ^ Фараго, Джейсон (7 марта 2019 г.). "T. Rex, как вы его не видели: с перьями". Нью-Йорк Таймс. Получено 7 марта 2019.
- ^ Барон, Мэтью Дж .; Норман, Дэвид Б.; Барретт, Пол М. (23 марта 2017 г.). «Новая гипотеза взаимоотношений динозавров и ранней эволюции динозавров». Природа. 543 (7646): 501–506. Bibcode:2017Натура.543..501Б. Дои:10.1038 / природа21700. PMID 28332513.
- ^ Ян, Цзысяо; Цзян, Баоюй; Макнамара, Мария Э .; Кирнс, Стюарт Л .; Питтман, Майкл; Kaye, Thomas G .; Орр, Патрик Дж .; Сюй, Син; Бентон, Майкл Дж. (17 декабря 2018 г.). «Покровные структуры птерозавров со сложным перьевидным ветвлением» (PDF). Природа Экология и эволюция. 3 (1): 24–30. Дои:10.1038 / с41559-018-0728-7. HDL:1983 / 1f7893a1-924d-4cb3-a4bf-c4b1592356e9. PMID 30568282.
- ^ Гринволд, Мэтью Дж .; Сойер, Роджер Х. (сентябрь 2013 г.). «Молекулярная эволюция и экспрессия β-кератинов архозавров: диверсификация и расширение β-кератинов архозавров и происхождение β-кератинов пера». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития. 320 (6): 393–405. Дои:10.1002 / jez.b.22514. PMID 23744807.
- ^ Хаксли, Т. (1868). «О животных, которые занимают промежуточное положение между птицами и рептилиями». Летопись и журнал естественной истории. 4-й. 2: 66–75.
- ^ Фостер, Майкл; Ланкестер, Э. Рей 1898–1903. Научные мемуары Томаса Генри Хаксли. 4 тома и приложение. Лондон: Макмиллан.[страница нужна ]
- ^ Хаксли, Томас Генри (1877). «Гипотеза эволюции. Нейтральные и благоприятные свидетельства».. В Хаксли, Томас Генри (ред.). Американские адреса, с лекцией по биологии. Нью-Йорк: Д. Эпплтон и компания.
- ^ "1970-е годы: новый дизайн динозавров". Палеоартистия. Получено 15 июн 2017.
- ^ Kaye, T.G .; Pittman, M .; Mayr, G .; Schwarz, D .; Сюй, X. (2019). "Обнаружение потерянного аира бросает вызов идентичности изолированного Археоптерикс пух Перо". Научные отчеты. 9 (1): 1182. Дои:10.1038 / s41598-018-37343-7.
- ^ а б Carney, R.M .; Tischlinger, H .; Шоуки, доктор медицины (2020). "Доказательства подтверждают идентичность изолированного окаменелого пера как прикрытия крыла Археоптерикс". Научные отчеты. 10: 15593. Дои:10.1038 / с41598-020-65336-у.
- ^ Герлинский, Г. (1996). «Перьевидные отпечатки на остатках теропод из нижней юры Массачусетса». Бюллетень Музея Северной Аризоны. 60: 179–184.
- ^ Кундрат, Мартин (15 июля 2004 г.). «Когда тероподы стали пернатыми? - свидетельство наличия перистых придатков до археоптерикса». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития. 302B (4): 355–364. Дои:10.1002 / jez.b.20014. PMID 15287100.
- ^ Зоология четвероногих @ScienceBlogs
- ^ Тернер, А. Х .; Makovicky, P.J .; Норелл М.А. (21 сентября 2007 г.). "Перья гусиные ручки в динозавре велоцирапторе". Наука. 317 (5845): 1721. Bibcode:2007Научный ... 317.1721Т. Дои:10.1126 / science.1145076. PMID 17885130.
- ^ Норелл М.А., Кларк Дж. М., Чиаппе Л. М., Дашзевег Д.; Кларк; Чиаппе; Дашзевег (1995). «Гнездовой динозавр». Природа. 378 (6559): 774–776. Bibcode:1995Натура 378..774Н. Дои:10.1038 / 378774a0.CS1 maint: несколько имен: список авторов (связь)
- ^ Хопп, Томас П .; Орсен, Марк Дж. (2004). «Наследственное поведение динозавров и происхождение перьев полета». В Currie, Philip J .; Koppelhus, Eva B .; Шугар, Мартин А .; Райт, Джоанна Л. (ред.). Пернатые драконы: исследования перехода от динозавров к птицам. Издательство Индианского университета. С. 234–250. ISBN 978-0-253-34373-4.
- ^ «Стенограмма: Динозавр, обманувший мир». BBC. Получено 22 декабря 2006.
- ^ Бок, Уолтер Дж. (Август 2000 г.). «Толковая история происхождения перьев1». Американский зоолог. 40 (4): 478–485. Дои:10.1668 / 0003-1569 (2000) 040 [0478: ehotoo] 2.0.co; 2.
- ^ Эмили Чанг (12 сентября 2011 г.). «Перья динозавров, найденные в янтаре Альберты». CBC. Получено 16 сентября 2011.
- ^ Свитек, Брайан (15 сентября 2011 г.). «Вкрапления янтаря демонстрируют доисторические перья». Природа. Дои:10.1038 / новости.2011.539.
- ^ Li, Q .; Gao, K.-Q .; Vinther, J .; Shawkey, M.D .; Clarke, J. A .; D'Alba, L .; Meng, Q .; Briggs, D. E. G .; Прум, Р. О. (2010). "Цветовые узоры оперения вымершего динозавра" (PDF). Наука. 327 (5971): 1369–1372. Bibcode:2010Sci ... 327.1369L. Дои:10.1126 / science.1186290. PMID 20133521.
- ^ Smithwick, F.M .; Nicholls, R .; Cuthill, I.C .; Винтер, Дж. (2017). «Затенение и полосы у тероподовых динозавров Sinosauropteryx выявляют гетерогенные среды обитания в раннемеловой биоте Джехол». Текущая биология. 27 (21): 3337–3343.e2. Дои:10.1016 / j.cub.2017.09.032. PMID 29107548.
- ^ Li, Q .; Gao, K.-Q .; Meng, Q .; Clarke, J.A .; Shawkey, M.D .; D’Alba, L .; Pei, R .; Ellison, M .; Norell, M.A .; Винтер, Дж. (2012). "Реконструкция Микрораптор и эволюция радужного оперения " (PDF). Наука. 335 (6073): 1215–1219. Bibcode:2012Научный ... 335.1215L. Дои:10.1126 / science.1213780. PMID 22403389.
- ^ Санкт-Флер, Николас (8 декабря 2016 г.). «Та штука с перьями в янтаре? Это был хвост динозавра». Нью-Йорк Таймс. Получено 8 декабря 2016.
- ^ Роми, Кристин (8 декабря 2016 г.). "Первый хвост динозавра, найденный в янтаре". Национальное географическое общество. Получено 12 декабря 2016.
- ^ Ринкон, Пол (8 декабря 2016 г.). "'В янтаре найден красивый хвост динозавра ». Новости BBC. Получено 8 декабря 2016.
- ^ Син, Лида; McKellar, Ryan C .; Сюй, Син; Ли, банда; Бай, Мин; Персоны, В. Скотт; Мияшита, Тецуто; Бентон, Майкл Дж .; Чжан, Цзяньпин; Вулф, Александр П .; Йи, Циру; Ценг, Куовей; Ран, Хао; Карри, Филип Дж. (Декабрь 2016 г.). "Хвост пернатого динозавра с примитивным оперением в янтаре среднего мелового периода". Текущая биология. 26 (24): 3352–3360. Дои:10.1016 / j.cub.2016.10.008. PMID 27939315.
- ^ Стивен Л. Брусатт, Грэм Т. Ллойд, Стив К. Ван, Марк А. Норелл (2014). «Постепенное построение птичьего строения тела привело к быстрым темпам эволюции перехода от динозавра к птице». Текущая биология. 24 (20): 2386–2392. Дои:10.1016 / j.cub.2014.08.034. PMID 25264248.CS1 maint: использует параметр авторов (связь)
- ^ Prum, R. & Brush A.H .; Кисть (2002). «Эволюционное происхождение и разнообразие перьев». Ежеквартальный обзор биологии. 77 (3): 261–295. Дои:10.1086/341993. PMID 12365352.
- ^ Прум, Р. (1999). «Развитие и эволюционное происхождение перьев». Журнал экспериментальной зоологии. 285 (4): 291–306. Дои:10.1002 / (SICI) 1097-010X (19991215) 285: 4 <291 :: AID-JEZ1> 3.0.CO; 2-9. PMID 10578107.
- ^ Сюй, Син; Норелл, Марк А .; Куанг, Сюэвэнь; Ван, Сяолинь; Чжао, Ци; Цзя, Чэнкай (октябрь 2004 г.). «Базальные тиранозавроиды из Китая и свидетельства праотцов у тираннозавроидов». Природа. 431 (7009): 680–684. Bibcode:2004Натура 431..680X. Дои:10.1038 / природа02855. PMID 15470426.
- ^ Норелл, М. Сюй, X. (2005) "Разновидности тираннозавров "Журнал" Естествознание ", май 2005 г.
- ^ Dawson, Tj; Мэлони, Словакия (2013). «Мех против перьев: разные роли меха красного кенгуру и перьев эму в терморегуляции в засушливой зоне Австралии». Австралийская маммология. 26 (2): 145. Дои:10.1071 / am04145.
- ^ Орсен, М. Дж .; Хопп, Т. П. (2004). Пернатые драконы: исследования перехода от динозавров к птицам. Издательство Индианского университета. С. 234–350.
- ^ а б Даймонд, К. К., Р. Дж. Кэбин и Дж. С. Брукс (2011). «Перья, динозавры и поведенческие сигналы: определение гипотезы визуального отображения для адаптивной функции перьев у нептичьих теропод». BIOS. 82 (3): 58–63. Дои:10.1893/011.082.0302.CS1 maint: использует параметр авторов (связь)
- ^ Sumida, S. S .; К. А. Брочу (2000). «Филогенетический контекст происхождения перьев». Американский зоолог. 40 (4): 485–503. Дои:10.1093 / icb / 40.4.486.
- ^ Лингхэм-Солиар, Т. (2011). «Эволюция пера: Sinosauropteryx, красочный хвост». Журнал орнитологии. 152 (3): 567–577. Дои:10.1007 / s10336-010-0620-у.
- ^ Vinther, J .; Д. Э. Г. Бриггс; Р. О. Прам и В. Саранатан (2008). «Цвет ископаемых перьев». Письма о биологии. 4 (5): 522–525. Дои:10.1098 / рсбл.2008.0302. ЧВК 2610093. PMID 18611841.
- ^ Чжан, Ф. К., С. Л. Кернс, П. Дж. Орр, М. Дж. Бентон, З. Х. Чжоу, Д. Джонсон, X. Сюй X. Л. Ван (2010). «Окаменелые меланосомы и окраска меловых динозавров и птиц» (PDF). Природа. 463 (7284): 1075–1078. Bibcode:2010 Натур.463.1075Z. Дои:10.1038 / природа08740. PMID 20107440.CS1 maint: использует параметр авторов (связь)
- ^ Li, Q .; Gao, K .; Vinther, J .; Shawkey, M.D .; Clarke, J. A .; D'Alba, L .; Мэн К. "Цветовые узоры оперения вымершего динозавра". Наука. 327: 1369–1372.
- ^ Зеленицкий, Дарла К .; Терриен, Франсуа; Эриксон, Грегори М .; DeBuhr, Christopher L .; Кобаяси, Ёсицугу; Eberth, David A .; Хэдфилд, Франк (2012). «Пернатые динозавры, не являющиеся птицами из Северной Америки, дают представление о происхождении крыльев». Наука. 338 (6106): 510–514. Bibcode:2012Научный ... 338..510Z. Дои:10.1126 / наука.1225376. PMID 23112330.
- ^ Ли, К. Г., К. К. Гао, К. Дж. Мэн, М. Д. Шоуки, Л. Д'Альба, Р. Пей, М. Эллисон, М. А. Норелл и Дж. Винтер (2012). "Реконструкция Микрораптор и эволюция радужного оперения ". Наука. 335 (6073): 1215–1219. Bibcode:2012Научный ... 335.1215L. Дои:10.1126 / science.1213780. PMID 22403389. S2CID 206537426.CS1 maint: использует параметр авторов (связь)
- ^ Dimond, C.C .; Cabin, R.J .; Брукс, Дж. С. «Перья, динозавры и поведенческие сигналы: определение гипотезы визуального отображения для адаптивной функции перьев у теропод, не являющихся птицами». BIOS. 82 (3): 58–63.
- ^ Rauhut, Oliver W. M .; Фот, Кристиан; Тишлингер, Гельмут; Норелл, Марк А. (17 июля 2012 г.). «Исключительно сохранившийся молодой мегалозавроидный динозавр-теропод с нитчатым покровом из поздней юры в Германии». Труды Национальной академии наук Соединенных Штатов Америки. 109 (29): 11746–11751. Bibcode:2012PNAS..10911746R. Дои:10.1073 / pnas.1203238109. ЧВК 3406838. PMID 22753486.
- ^ Гулд, Стивен Дж .; Врба, Элизабет С. (1982). «Экзаптация: недостающий термин в науке о форме» (PDF). Палеобиология. 8 (1): 4–15. Дои:10.1017 / S0094837300004310.
- ^ Райххольф, Дж. Х. (1996). "Die Feder, die Mauser und der Ursprung der Vögel. Ein neure Sicht zur Evolution der Vögel" [Перо, линька и происхождение птиц. Новый взгляд на эволюцию птиц. Археоптерикс (на немецком). 14: 27–38.
- ^ Бок, Уолтер Дж. (1 августа 2000 г.). «Толковая история происхождения перьев». Американский зоолог. 40 (4): 478–485. CiteSeerX 10.1.1.497.1279. Дои:10.1093 / icb / 40.4.478.
- ^ Ян, З .; Jiang, B .; McNamara, M.E .; Kearns, S.L .; Pittman, M .; Kaye, T. G .; Орр, П. Дж .; Сюй, X .; Бентон, М. Дж. (2019). «Покровные структуры птерозавров со сложным перьевидным ветвлением». Природа Экология и эволюция. 3: 24–30.
- ^ Майр, Джеральд; Петерс, Стефан; Плодовски, Герхард; Фогель, Олаф (2002). "Щетиноподобные покровные структуры на хвосте рогатого динозавра. Пситтакозавр". Naturwissenschaften. 89 (8): 361–365. Bibcode:2002NW ..... 89..361M. Дои:10.1007 / s00114-002-0339-6. PMID 12435037.
- ^ Чжэн, Сяо-Тин; Ты, Хай-Лу; Сюй, Син; Донг, Чжи-Мин (2009). "Раннемеловой гетеродонтозаврид динозавр с нитчатыми покровными структурами". Природа. 458 (7236): 333–336. Bibcode:2009Натура.458..333Z. Дои:10.1038 / природа07856. PMID 19295609.
- ^ Сюй, X .; Чжэн, X .; Вы, Х. (2009). «Новый тип перьев у тероподов-нонавийцев и ранняя эволюция перьев». Труды Национальной академии наук. 106 (3): 832–4. Bibcode:2009PNAS..106..832X. Дои:10.1073 / pnas.0810055106. ЧВК 2630069. PMID 19139401.
- ^ Godefroit, P; Синица, С; Дуайи, Д; Болотский, У; Сизов, А (2013). Перьевидные структуры и чешуя у юрского неорнитишского динозавра из Сибири.
- ^ Чжан, Фучэн; Чжоу, Чжунхэ; Сюй, Син; Ван, Сяолинь; Салливан, Корвин (2008). «Причудливый юрский манирапторан из Китая с удлиненными ленточными перьями» (PDF). Природа. 455 (7216): 1105–1108. Bibcode:2008 Натур.455.1105Z. Дои:10.1038 / природа07447. PMID 18948955.
- ^ Зеленицкий, Д.К .; Therrien, F .; Эриксон, Г. М .; DeBuhr, C.L .; Кобаяши, Ю .; Eberth, D. A .; Хэдфилд, Ф. (25 октября 2012 г.). «Пернатые динозавры, не являющиеся птицами из Северной Америки, дают представление о происхождении крыльев». Наука. 338 (6106): 510–514. Bibcode:2012Научный ... 338..510Z. Дои:10.1126 / наука.1225376. PMID 23112330.
- ^ Годфруа, Паскаль; Кау, Андреа; Ху, Дун-Ю; Эскилье, Франсуа; Ву, Венхао; Дайк, Гарет (2013). «Юрский птичий динозавр из Китая решает раннюю филогенетическую историю птиц». Природа. 498 (7454): 359–362. Bibcode:2013Натура.498..359G. Дои:10.1038 / природа12168. PMID 23719374.
- ^ Сюй, Син (2020), Фот, Кристиан; Раухут, Оливер В. М. (ред.), «Нитчатые покровы тероподов Nonavialan и их родственников: достижения и перспективы понимания эволюции перьев», Эволюция перьев: от их происхождения до наших дней, Увлекательные науки о жизни, Cham: Springer International Publishing, стр. 67–78, Дои:10.1007/978-3-030-27223-4_5, ISBN 978-3-030-27223-4, получено 24 октября 2020
внешняя ссылка
- Цвет динозавра и перья, Бристольский университет, 12 февраля 2019
- DinoBuzz, объяснение противоречия между динозаврами и птицами в Калифорнийском университете в Беркли.
- Пернатые динозавры на В наше время на BBC