Альбертозавр - Википедия - Albertosaurus
| Альбертозавр | |
|---|---|
 | |
| Установлен литой в Научный центр Айовы в Де-Мойне, штат Айова | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Tyrannosauridae |
| Подсемейство: | †Альбертозаврины |
| Род: | †Альбертозавр Осборн, 1905 |
| Разновидность: | †А. саркофаг |
| Биномиальное имя | |
| †Саркофаг альбертозавра Осборн, 1905 г. | |
| Другие виды | |
| |
| Синонимы | |
| |
Альбертозавр (/æлˌбɜːrтəˈsɔːrəs/; означает "ящерица Альберты") является род из тираннозаврид теропод динозавры которые жили на западе Северной Америки во время Поздний мел Период, около 70миллион лет назад. В типовой вид, А. саркофаг, очевидно, был ограничен в классифицировать к современности Канадская провинция из Альберта, в честь которого назван род, хотя и неопределенный вид ("ср. Альбертозавр sp. ") была обнаружена в Corral de Enmedio и Packard Formations в Мексика.[1] Ученые расходятся во мнениях относительно содержания рода, при этом некоторые признают Горгозавр libratus как второй вид.
Как тираннозаврид, Альбертозавр был двуногий хищник с крошечными двупалыми руками и массивной головой с множеством больших острых зубов. Возможно, он был в верхней части пищевая цепочка в своем местном экосистема. Пока Альбертозавр был большим для теропод, он был намного меньше, чем его более крупный и известный родственник тиранозавр Рекс, ростом от 9 до 10 м (от 30 до 33 футов) и, возможно, весом 2,5 тонны (2,8 коротких тонны) или меньше.
С момента первого открытия в 1884 г. окаменелости из более чем 30 человек были восстановлены, что дает ученым более подробную информацию о Альбертозавр анатомии, чем у большинства других тираннозавридов. Обнаружение 26 человек на одном участке свидетельствует о том, что пакет поведение и позволяет изучать онтогенез и популяционная биология, что невозможно с менее известными динозаврами.
История открытия
Именование

Альбертозавр был назван Генри Фэрфилд Осборн в примечании на одну страницу в конце его описания 1905 г. тиранозавр Рекс.[2]:265 Имя почести Альберта, то Канадская провинция установлен в том же году, в котором были обнаружены первые останки. Общее название также включает в себя Греческий срок σαυρος/sauros («ящерица»), самый распространенный суффикс в именах динозавров. В типовой вид является Саркофаг альбертозавра; то конкретное имя происходит от древнегреческого σαρκοφάγος (саркофаги), что означает "поедание плоти" и имеющий то же этимология как погребальный контейнер с которым он разделяет свое название: сочетание греческих слов σαρξ /саркс («плоть») и φαγειν /фагеин ("есть").[2] Науке известно более 30 особей всех возрастов.[3][4]
Ранние открытия

В типовой образец частичный череп, собранный летом 1884 г. обнажение из Формация каньона Подкова рядом с Ред Дир Ривер, в Альберте. Этот экземпляр, найденный 9 июня 1884 г., был обнаружен экспедицией Геологическая служба Канады во главе с известным геолог Джозеф Берр Тиррелл. Из-за отсутствия специального оборудования почти весь череп можно было закрепить только частично. В 1889 году коллега Тиррелла Томас Чесмер Уэстон нашел неполный череп меньшего размера, связанный с некоторым скелетным материалом в месте поблизости.[5] Два черепа были отнесены к ранее существовавшим видам Laelaps incrassatus к Эдвард Дринкер Коуп в 1892 г.,[6] хотя имя Лаэлапс был озабоченный родом клещ и был изменен на Дриптозавр в 1877 г. Отниэль Чарльз Марш. Коуп отказался признать новое имя, придуманное его заклятым соперником Маршем. Тем не мение, Лоуренс Ламбе использовал имя Dryptosaurus incrassatus вместо Laelaps incrassatus когда он подробно описал останки в 1903 и 1904 годах,[7][8] комбинация, впервые придуманная Оливер Перри Хэй в 1902 г.[9]
Вскоре Осборн отметил, что D. incrassatus был основан на типичных зубах тираннозавридов, поэтому два черепа в каньоне Подкова нельзя было с уверенностью отнести к этому виду. Черепа каньона Подкова также заметно отличались от останков D. aquilunguis, типовые виды Дриптозавр, поэтому Осборн создал новое имя Саркофаг альбертозавра для них в 1905 году. Он не описал останки во всех подробностях, сославшись на полное описание Ламбе годом ранее.[2] Оба экземпляра ( голотип CMN 5600 и паратип CMN 5601) хранятся в Канадский музей природы в Оттава. К началу двадцать первого века возникли некоторые опасения, что из-за поврежденного состояния голотипа Альбертозавр может быть номен дубиум, «сомнительное имя», которое могло быть использовано только для самого типового образца, потому что другие окаменелости не могли быть достоверно присвоены ему. Однако в 2010 г. Томас Карр установили, что голотип, паратип и сопоставимые более поздние находки имеют одну общую уникальную черту или аутапоморфия: наличие увеличенного пневматического отверстия в задней кромке боковой части небная кость, доказывая, что Альбертозавр был действительным таксон.[10]
Костное ложе Dry Island

11 августа 1910 г. американский палеонтолог Барнум Браун обнаружил останки большой группы Альбертозавр в другом карьер рядом с рекой Ред-Дир. Из-за большого количества костей и ограниченного времени группа Брауна не собирала все образцы, а старалась собрать останки всех людей, которых они могли идентифицировать в костяк. Среди костей, отложенных в Американский музей естественной истории коллекции в Нью-Йорк семь наборов правильных плюсневые кости вместе с двумя изолированными костями пальца, которые не соответствовали размеру плюсневых костей. Это указывало на присутствие в карьере не менее девяти человек. Палеонтолог Филип Дж. Карри из Королевский музей палеонтологии Тиррелла заново открыл костяной пласт в 1997 году и возобновил полевые работы на участке, который сейчас находится внутри Провинциальный парк Dry Island Buffalo Jump.[11] В ходе дальнейших раскопок с 1997 по 2005 год были обнаружены останки еще 13 человек разного возраста, в том числе миниатюрного двухлетнего ребенка и очень старого человека, длина которого оценивалась более чем в 10 метров (33 фута). Ни один из этих людей не известен по полным скелетам, и большинство из них представлено останками в обоих музеях.[3][12] Раскопки продолжались до 2008 года, когда было установлено минимальное количество присутствующих особей - 12 человек на основе сохранившихся элементов, которые встречаются только один раз в скелете, и 26, если учитывались зеркальные элементы, различающиеся по размеру из-за онтогенез. Всего 1128 Альбертозавр кости были закреплены, это была самая большая концентрация окаменелостей крупных теропод, известная в меловом периоде.[13]
Другие открытия

В 1911 году Барнум Браун во время второго года работы Американского музея естественной истории в Альберте обнаружил фрагментарный частичный Альбертозавр череп на реке Ред-Дир у моста Толмена, экз. AMNH 5222.[14]
Уильям Паркс описал новый вид в 1928 г., Альбертозавр arctunguis, основанный на частичном скелете без черепа, раскопанном Гасом Линдбладом и Ральфом Хорнеллом у реки Ред-Дир в 1923 году,[15] но этот вид считался идентичным А. саркофаг с 1970 г.[16] Образец Парка (ROM 807) находится в Королевский музей Онтарио в Торонто.
Между 1926 и 1972 годами нет Альбертозавр окаменелости вообще были обнаружены; но, начиная с семидесятых годов, количество известных материалов постоянно увеличивается. Помимо костного ложа на Сухом острове, с тех пор в Альберте были обнаружены еще шесть черепов и скелетов, которые хранятся в различных канадских музеях: образцы RTMP 81.010.001, найденные в 1978 году палеонтологом-любителем Морисом Стефанюком; RTMP 85.098.001, обнаруженный Стефанюком 16 июня 1985 г .; RTMP 86.64.001 (декабрь 1985 г.); РТМП 86.205.001 (1986); РТМП 97.058.0001 (1996); и CMN 11315. Однако из-за вандализма и несчастных случаев среди этих находок не удалось найти неповрежденных и целых черепов.[5] Об окаменелостях также сообщалось из американских штатов Монтана, Нью-Мексико, Вайоминг, и Миссури, но они, вероятно, не представляют А. саркофаг и может даже не принадлежать к роду Альбертозавр.[17][4]
Два экземпляра из ("ср. Альбертозавр ".sp") были найдены в Мексика (Packard Formation и Формация Корраль-де-Энмедио ).[1]
Горгозавр libratus

В 1913 г. палеонтолог Чарльз Х. Штернберг восстановил еще один скелет тиранозаврида от немного более старого Формирование парка динозавров в Альберте. Лоуренс Ламбе назвал этого динозавра Горгозавр libratus в 1914 г.[18] Другие образцы позже были обнаружены в Альберте и американском штате Монтана. Находка, во многом из-за отсутствия хороших Альбертозавр материал черепа, нет существенных различий для разделения двух таксонов, Дейл Рассел объявил имя Горгозавр а младший синоним из Альбертозавр, который был назван первым, и G. libratus был переименован Альбертозавр libratus в 1970 г. Различие видов сохранялось из-за разницы в возрасте. Это дополнение расширило временной диапазон рода Альбертозавр назад на несколько миллионов лет и его ареал на юг на сотни километров.[16]
В 2003 г. Филип Дж. Карри, воспользовавшись гораздо более обширными находками и общим увеличением анатомических знаний теропод, сравнили несколько черепов тираннозавров и пришли к выводу, что эти два вида более различны, чем считалось ранее. Решение об использовании одного или двух родов довольно произвольно, поскольку эти два вида сестринские таксоны, более тесно связанные друг с другом, чем с любыми другими видами. Признавая это, Карри, тем не менее, рекомендовал Альбертозавр и Горгозавр быть сохранены как отдельные роды, поскольку он пришел к выводу, что они не более похожи, чем Дасплетозавр и Тиранозавр, которые почти всегда разделены. Кроме того, несколько образцов альбертозаврина были извлечены из Аляска и Нью-Мексико, и Карри предположил, что Альбертозавр-Горгозавр ситуация может быть прояснена после их полного описания.[4] Большинство авторов следовали рекомендации Карри,[17][12][19] но некоторые нет.[20]
Другие виды
Помимо А. саркофаг, A. arctunguis и A. libratus, несколько других видов Альбертозавр были названы. Все они сегодня рассматриваются как более молодые синонимы других видов или как Номина Дубия, и не закреплены за Альбертозавр.
В 1930 г. Анатолий Николаевич Рябинин названный Альбертозавр периколозус основанный на зубе из Китая, который, вероятно, принадлежал Тарбозавр.[21] В 1932 г. Фридрих фон Хюене переименован Dryptosaurus incrassatus, не считается номен дубиум им, чтобы Альбертозавр incrassatus.[22] Потому что он определил Горгозавр с Альбертозавр, в 1970 году Рассел также переименовал Горгозавр стернберги (Мэтью и Браун 1922) в Альбертозавр sternbergi и Горгозавр лансенсис (Гилмор 1946) в Альбертозавр лансенсис.[16] Первый вид сегодня рассматривается как ювенильная форма Горгозавр libratus, последний как идентичный Тиранозавр или представляющие отдельный род Нанотираннус. В 1988 г. Грегори С. Пол основан Альбертозавр мегаграцилис на скелете небольшого тираннозаврида, экземпляр LACM 28345, из Формация Адского ручья Монтаны.[23] Он был переименован Динотираннус в 1995 г.[24] но теперь считается, что представляет собой несовершеннолетнего тиранозавр Рекс.[25] Также в 1988 году Пол переименовал Алектрозавр Olseni (Гилмор 1933) в Альбертозавр olseni;[23] это не нашло всеобщего признания. В 1989 г. Горгозавр novojilovi (Малеев 1955) был переименован Брин Мадер и Роберт Брэдли как Альбертозавр novojilovi;[26] сегодня это рассматривается как синоним Тарбозавр.
В двух случаях виды на основе действительных Альбертозавр материал перенесли в другой род: в 1922 г. Уильям Диллер Мэтью переименован А. саркофаг в Дейнодон саркофаг[27] а в 1939 г. немецкий палеонтолог Оскар Кун переименован A. arctunguis в Deinodon arctunguis.[28]
Описание
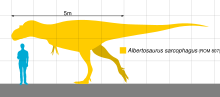
Альбертозавр был крупнее большинства тираннозавридов, но меньше Тарбозавр и Тиранозавр. Типичный Альбертозавр взрослые достигли длины до 9 м (30 футов),[16][17] в то время как редкие особи преклонного возраста могут вырасти до более 10 метров (33 футов) в длину.[3] Несколько независимых оценок массы, полученных разными методами, позволяют предположить, что взрослый Альбертозавр весил от 1,3тонны и 2,5 тонны (2,8 тонны).[29][12][30] В 2016 году Молина-Перес и Ларраменди оценили образец (CMN 5600) в 9,7 метра (32 фута) и 4 тонны (4,4 коротких тонны).[31]
Альбертозавр имели схожий внешний вид со всеми другими тираннозавридами. Обычно у тероподов Альбертозавр был двуногим и уравновешивал тяжелую голову и торс с длинным хвостом. Однако передние конечности тираннозавров были чрезвычайно малы для своего размера и сохраняли только два пальца. Задние конечности были длинными и оканчивались четырехпалой ступней, на которой первый палец, называемый Hallux, был коротким и не достиг земли. Третья цифра была длиннее остальных.[17] Альбертозавр могли развить скорость ходьбы 14-21 км / час (8-13 миль / час).[32] По крайней мере, для более молодых людей высокая скорость бега правдоподобна.[11]
Два слепка кожи от Альбертозавр известны, оба показывают напольные весы.[33] На одном участке обнаружены ребра желудка и отпечаток длинной неизвестной кости, что указывает на то, что это пятно с живота. Чешуя галечная, постепенно увеличивается в размерах и имеет несколько шестиугольную форму. Также сохранились две более крупные шкалы, расположенные на расстоянии 4,5 см друг от друга.[33] Еще один отпечаток кожи - с неизвестной части тела. Эти чешуи мелкие, ромбовидные, расположены рядами.[33]
Череп и зубы

Массивный череп из Альбертозавр, который сидел на короткой S-образной шее, у самых крупных взрослых особей был около 1 метра (3,3 фута) в длину.[25] Широкие отверстия в черепе (фенестры ) уменьшил вес головы, а также предоставил место для мышца привязанность и органы чувств. Его длинные челюсти содержали, вместе взятые, 58 или более зубов в форме банана; у более крупных тираннозавридов было меньше зубов; Горгозавр в 62 раза больше. В отличие от большинства теропод, Альбертозавр и другие тираннозавриды были гетеродонт, с зубами разной формы в зависимости от их положения во рту. В предчелюстной зубы на кончике верхней челюсти, по четыре с каждой стороны, были намного меньше остальных, плотнее прилегали и имели D-образную форму. поперечное сечение.[17] Как с Тиранозавр, верхнечелюстные (щечные) зубы Альбертозавр были адаптированы в общем виде, чтобы противостоять боковым силам со стороны сопротивляющейся добычи. Сила укуса Альбертозавр был менее грозным, однако, с максимальной силой на задних зубах, достигающей 3413 ньютонов.[34] Над глазами были короткие костлявые гребни, которые, возможно, были ярко окрашены при жизни и использовались во время ухаживания для привлечения партнера.[35]

Уильям Эблер заметил в 2001 году, что Альбертозавр зубцы напоминать трескаться в зубе, оканчивающемся круглой пустотой, называемой ампулой.[36] Зубы тиранозавров использовались в качестве опор для отрыва плоти от тела, поэтому, когда тираннозавр тянул назад кусок мяса, натяжение могло привести к тому, что зубцы, похожие на трещины, распространились по зубу.[36] Однако наличие ампулы распределяло эти силы на большую площадь поверхности, и уменьшил риск повреждения зуба при растяжении.[36] Наличие разрезов, заканчивающихся пустотами, имеет параллели в человеческой инженерии. Производители гитар используют надрезы, оканчивающиеся пустотами, чтобы, как описывает Аблер, «придавать чередующиеся области гибкости и жесткости» дереву, с которым они работают.[36] Использование дрель для создания своего рода «ампулы» и предотвращения распространения трещин в материале также используется для защиты поверхностей самолетов.[36] Abler продемонстрировал, что стержень из оргстекла с надрезами, называемыми «пропилами», и просверленными отверстиями был более чем на 25% прочнее, чем стержень с регулярными надрезами.[36] В отличие от тираннозавров, древние хищники любят фитозавры и Диметродон не имели приспособлений для предотвращения распространения трещиноподобных зубцов на их зубах под воздействием сил кормления.[36]
Классификация и систематика
Альбертозавр является членом теропод семья Tyrannosauridae, в подсемействе Albertosaurinae. Ближайший родственник - немного старше Горгозавр libratus (иногда называют Альбертозавр libratus; Смотри ниже).[37] Эти два вида являются единственными описанными альбертозавринами; могут существовать другие неописанные виды.[4] Томас Хольц найденный Аппалачиозавр быть альбертозаврином в 2004 году,[17] но его более поздняя неопубликованная работа находит его недалеко от Tyrannosauridae,[38] по согласованию с другими авторами.[20]
Другое крупное подсемейство тираннозаврид - Tyrannosaurinae, которое включает Дасплетозавр, Тарбозавр и Тиранозавр. По сравнению с этими крепкими тираннозавринами, альбертозаврины имели стройное телосложение, с пропорционально меньшими черепами и более длинными костями голени (большеберцовая кость ) и ступни (плюсневые кости и фаланги ).[25][37]

Ниже представлена кладограмма тираннозавров, основанная на филогенетический анализ дирижирует Loewen и другие. в 2013.[39]
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
Схема роста

Большинство возрастных категорий Альбертозавр представлены в Окаменелости. Использование кости гистология часто можно определить возраст отдельного животного на момент смерти, что позволяет оценить темпы роста и сравнить с другими видами. Самый молодой из известных Альбертозавр - двухлетний ребенок, обнаруженный на костном ложе Сухого острова, который весил бы около 50 килограммов (110фунт ) и имели длину чуть более 2 метров (6,6 футов). Образец 10 метров (33 фута) из того же карьера - самый старый и самый крупный из известных, возраст 28 лет. Когда образцы среднего возраста и размера нанесены на график, SВ результате получается кривая роста в форме, причем наиболее быстрый рост происходит в четырехлетний период, заканчивающийся примерно на шестнадцатом году жизни, что также наблюдается у других тираннозавридов. Скорость роста на этом этапе составляла 122 килограмма (269 фунтов) в год, из расчета на 1,3 взрослого человека.тонны. Другие исследования предполагали более высокий вес взрослых; это повлияет на величину темпа роста, но не на общую картину. Тираннозавриды размером с Альбертозавр имели аналогичные темпы роста, хотя гораздо больше тиранозавр Рекс выросли почти в пять раз больше (601 килограмм [1325 фунтов] в год) на пике.[3] Окончание фазы быстрого роста предполагает начало половая зрелость в Альбертозавр, хотя рост продолжался более медленными темпами на протяжении всей жизни животных.[3][12] Половое созревание при активном росте, по-видимому, является общей чертой среди мелких[40] и большой[41] динозавров, а также крупных млекопитающих, таких как люди и слоны.[41] Этот образец относительно раннего полового созревания разительно отличается от образца у птиц, у которых половая зрелость откладывается до тех пор, пока они не закончат расти.[19][41]
Во время роста из-за утолщения морфология зубов так сильно изменилась, что, если бы ассоциация скелетов молодых и взрослых на костном дне Сухого острова не была доказана, что они принадлежат к одному таксону, зубы молодых, вероятно, были бы идентифицированы статистическим анализом как зубы другой вид.[42]
История жизни
Самый известный Альбертозавр люди были в возрасте 14 лет и старше на момент смерти. Молодые животные редко встречаются в окаменелостях по нескольким причинам, в основном предвзятость сохранения, где меньшие кости более молодых животных с меньшей вероятностью будут сохранены путем окаменения, чем более крупные кости взрослых особей, и систематическая ошибка сбора, когда более мелкие окаменелости с меньшей вероятностью будут замечены коллекционерами в полевых условиях.[43] Молодой Альбертозавр относительно велики для молодых животных, но их останки все еще редки в летописи окаменелостей по сравнению с взрослыми особями. Было высказано предположение, что это явление является следствием история жизни, а не предвзятость, и что окаменелости молодых Альбертозавр редки, потому что они просто не умирали так часто, как взрослые.[3]
Гипотеза Альбертозавр история жизни постулирует, что детеныши погибли в большом количестве, но не сохранились в летописи окаменелостей из-за их небольшого размера и хрупкой конструкции. Спустя всего два года молодые особи были крупнее любого другого хищника в регионе, за исключением взрослых особей. Альбертозавр, и более быстрым, чем большинство их хищных животных. Это привело к резкому снижению их смертности и соответствующей редкости ископаемых останков. Уровень смертности удвоился в возрасте двенадцати лет, возможно, в результате физиологических требований фазы быстрого роста, а затем снова удвоился с наступлением половой зрелости в возрасте от четырнадцати до шестнадцати лет. Этот повышенный уровень смертности сохранялся на протяжении всей взрослой жизни, возможно, из-за высоких физиологических требований к деторождению, включая стресс и травмы, полученные во время внутривидовой конкуренции за партнеров и ресурсы, и, в конечном итоге, постоянно усиливающиеся эффекты старение. Более высокий уровень смертности взрослых может объяснить их более частую сохранность. Очень большие животные были редкостью, потому что немногие особи выживали достаточно долго, чтобы достичь таких размеров. Высокий уровень младенческой смертности, за которым следует снижение смертности среди молоди и внезапное увеличение смертности после половой зрелости, когда очень немногие животные достигают максимального размера, - это закономерность, наблюдаемая у многих современных крупных млекопитающих, включая слонов, Африканский буйвол, и носорог. Та же картина наблюдается и у других тираннозавридов. Сравнение с современными животными и другими тираннозавридами подтверждает эту гипотезу истории жизни, но предвзятость в летописи окаменелостей все еще может играть большую роль, тем более что более двух третей всех Альбертозавр экземпляры известны из одного местонахождения.[3][19][44]
Поведение упаковки

Костное ложе Сухого острова, обнаруженное Барнумом Брауном и его командой, содержит останки 26 человек. Альбертозавр, наибольшее количество особей, обитающих в одном месте среди крупных тероподов мелового периода, и второе место среди крупных тероподных динозавров после Аллозавр сборка на Карьер динозавров Кливленд-Ллойд в Юта. Кажется, что группа состоит из одного очень старого взрослого; восемь взрослых в возрасте от 17 до 23 лет; семь суб-взрослых, переживающих фазы быстрого роста в возрасте от 12 до 16 лет; и шесть молодых особей в возрасте от 2 до 11 лет, которые еще не достигли фазы роста.[3]
Практически полное отсутствие травоядное животное остается и аналогичное состояние сохранности, общее для многих людей в Альбертозавр Костяной карьер привел Карри к выводу, что это место не было ловушкой для хищников, как La Brea Tar Pits в Калифорния, и что все сохранившиеся животные погибли одновременно. Карри утверждает, что это свидетельство поведения стаи.[11] Другие ученые настроены скептически, отмечая, что животные могли быть сбиты вместе из-за засухи, наводнения или по другим причинам.[3][43][45]

Существует множество свидетельств стадного поведения травоядных динозавров, в том числе цератопсы и гадрозавры.[46] Однако очень редко в одном и том же месте можно найти так много хищников-динозавров. Маленькие теропод, такие как Дейноних[47] и Целофиз[48] были найдены в скоплениях, как и более крупные хищники, такие как Аллозавр и Мапузавр.[49] Есть некоторые свидетельства стадное поведение и у других тираннозавридов. Рядом с "Сью ", Тиранозавр установлен в Полевой музей естественной истории в Чикаго, и костяк в Формирование двух лекарств Монтаны содержит не менее трех экземпляров Дасплетозавр, сохранились рядом с несколькими гадрозаврами.[50] Эти результаты могут подтвердить данные о социальном поведении в Альбертозавр, хотя некоторые или все вышеперечисленные местонахождения могут представлять собой временные или неестественные скопления.[11] Другие предполагают, что вместо социальных групп, по крайней мере, некоторые из этих находок представляют Дракон Комодо -подобное преследование туш, когда агрессивная конкуренция приводит к гибели некоторых хищников и каннибализированный.[43]
Карри также высказал предположение об охотничьих привычках Альбертозавр. Пропорции ног у более мелких особей были сопоставимы с пропорциями ног. орнитомимиды, которые, вероятно, были одними из самых быстрых динозавров. Моложе Альбертозавр вероятно, были столь же быстроногими или, по крайней мере, быстрее своей добычи. Карри предположил, что младшие члены стаи могли быть ответственны за то, чтобы направить свою добычу к взрослым, которые были крупнее и мощнее, но также медленнее.[11] Молодые особи, возможно, вели другой образ жизни, чем взрослые, что является хищником. ниши между огромными взрослыми особями и более мелкими современными тероподами, самыми большими из которых были два порядки величины меньше взрослого Альбертозавр в массе.[17] Похожая ситуация наблюдается у современных драконов Комодо, когда вылупившиеся детеныши начинают жизнь маленькими. насекомоядные прежде чем вырасти, чтобы стать доминирующими хищниками на своих островах.[51] Однако, поскольку сохранение поведения в летописи окаменелостей чрезвычайно редко, эти идеи нелегко проверить. В 2010 году Карри, по-прежнему поддерживая гипотезу охотничьих стай, признал, что концентрация могла быть вызвана другими причинами, такими как медленное повышение уровня воды во время продолжительного наводнения.[52]
Палеопатология

В 2009 году исследователи выдвинули гипотезу, что отверстия с гладкими краями, найденные в окаменелых челюстях тираннозаврид динозавры, такие как Альбертозавр были вызваны паразитом, похожим на Трихомонада галлины, которым заражаются птицы.[53] Они предположили, что тиранозавриды передают инфекцию, кусая друг друга, и что инфекция ухудшает их способность есть пищу.[53]
В 2001, Брюс Ротшильд и другие опубликовали исследование, посвященное изучению доказательств стрессовые переломы и отрыв сухожилия в теропод динозавры и последствия для их поведения. Они обнаружили, что только один из 319 Альбертозавр Кости стопы, проверенные на наличие стрессовых переломов, на самом деле имели их, а ни одна из четырех костей руки не обнаружила. Ученые обнаружили, что стрессовые переломы «значительно» менее распространены в Альбертозавр чем в карнозавр Аллозавр.[54] ROM 807, голотип A. arctunguis (теперь упоминается А. саркофаг), имел глубокое отверстие 2,5 на 3,5 см (0,98 на 1,38 дюйма) в подвздошная лопатка, хотя описатель вида не признал это патологическим. Образец также содержит некоторые экзостоз на четвертой левой плюсне. В 1970 году двое из пяти Саркофаг альбертозавра Дейл Рассел сообщил, что образцы с плечевыми костями имеют патологические повреждения.[55]
В 2010 г. здоровье Сухого острова Альбертозавр о сборке не поступало. Большинство образцов не показывали никаких признаков болезни. На трех фалангах стопы странные костные шпоры, состоящие из аномальных окостенений сухожилий, так называемых энтезофиты, присутствовали, их причина неизвестна. Два ребра и ребро на животе показали признаки перелома и заживления. У одного взрослого экземпляра была левая нижняя челюсть с колотой раной и как зажившими, так и незажившими следами укусов. Небольшое количество аномалий выгодно отличается от состояния здоровья Маджунгазавр популяции, из которой в 2007 г. было установлено, что 19% лиц имеют костные патологии.[56]
Палеоэкология

Большинство окаменелостей Саркофаг альбертозавра известны с верхних Формация каньона Подкова в Альберте. Эти более молодые единицы этого геологическая формация свидание с ранним Маастрихтский этап Поздний мел Период с 70 по 68 год Ма (миллион лет назад). Сразу под этой формацией находится Медвежья лапа, а морской формация, представляющая часть Западный внутренний морской путь. Морской путь отступал по мере того, как климат остывал, а уровень моря понижался к концу мелового периода, обнажая землю, которая ранее была под водой. Однако это не был гладкий процесс, и морской путь периодически поднимался, чтобы покрыть части региона по всему каньону Подкова, прежде чем, наконец, полностью отступить в последующие годы. Из-за изменения уровня моря в формации Хорсшу-Каньон представлено множество различных сред, включая прибрежные и прибрежные морские среды обитания, а также прибрежные среды обитания, такие как лагуны, эстуарии и приливные отмели. Многочисленные каменный уголь швы представляют собой древние торф болота. Как и большинство других позвоночное животное окаменелости из формации, Альбертозавр останки найдены в отложениях, заложенных в дельты и поймы крупных рек во второй половине времен каньона Подкова.[57]
В фауна формации Хорсшу-Каньон хорошо известны, так как окаменелости позвоночных, в том числе динозавров, довольно распространены. Акулы, лучи, осетровые, Bowfins, Гарс и гар-подобный Аспидоринх составили ихтиофауну. Включены млекопитающие многотуберкулезные и сумчатое животное Дидельфодон. Соленая вода плезиозавр Левоспондил был обнаружен в морских отложениях в каньоне Подкова, в то время как в пресноводных средах обитали черепахи, Шампсозавр, и крокодилы подобно Leidyosuchus и Стангерочампса. Динозавры доминируют в фауне, особенно гадрозавры, которые составляют половину всех известных динозавров, включая роды. Эдмонтозавр, Зауролоф и Гипакрозавр. Цератопсы и орнитомимиды были также очень распространены, вместе составляя еще треть известной фауны. Наряду с гораздо более редкими анкилозавры и пахицефалозавры, все эти животные были бы добычей разнообразных плотоядных теропод, включая троодонтиды, дромеозавриды, и ценагнатиды.[57][58] В сочетании с Альбертозавр остатки костного ложа Сухого острова, кости малого теропод Альбертоникус были найдены.[59] Взрослый Альбертозавр были высшие хищники в этой среде с промежуточными нишами, возможно, заполненными молодыми альбертозаврами.[57]
Смотрите также
Рекомендации
- ^ а б Перечислено как "ср. Альбертозавр sp. »« Corral De Enmedio and Packard Formations, Cabollona Group, Sonora, Mexico »в Sullivan and Lucas (2006). Стр. 16.
- ^ а б c Осборн, Генри Ф. (1905). «Тираннозавр и другие хищные динозавры мелового периода». Бюллетень Американского музея естественной истории. 21 (3): 259–265. Дои:10.1111 / j.1468-5965.2007.00735_17.x. HDL:2246/1464.
- ^ а б c d е ж грамм час я Эриксон, Грегори М .; Карри, Филип. J .; Inouye, Brian D .; Винн, Алиса А. (2006). «Таблицы дожития тираннозавров: пример популяционной биологии неавианских динозавров» (PDF). Наука. 313 (5784): 213–217. Bibcode:2006Научный ... 313..213E. Дои:10.1126 / science.1125721. PMID 16840697. S2CID 34191607. Архивировано из оригинал (PDF) на 2010-07-18. Получено 2010-08-29.
- ^ а б c d Карри, Филип Дж. (2003). "Анатомия черепа тираннозаврид из позднего мела Альберты" (PDF). Acta Palaeontologica Polonica. 48 (2): 191–226.
- ^ а б Танке, Даррен Х .; Карри, Филип Дж. (2010). "История Альбертозавр открытия в Альберте, Канада ". Канадский журнал наук о Земле. 47 (9): 1197–1211. Bibcode:2010CaJES..47.1197T. Дои:10.1139 / e10-057.
- ^ Коуп, Эдвард Д. (1892). "На черепе динозавра Laelaps incrassatus Справиться". Американское философское общество, Труды. 30: 240–245.
- ^ Ламбе, Л. М. (1903). "На нижней челюсти Dryptosaurus incrassatus (Справиться)". Оттавский натуралист. 17: 134.
- ^ Ламбе, Лоуренс М. (1904). "На Dryptosaurus incrassatus (Cope) из серии Эдмонтон Северо-Западного края ». Вклад в канадскую палеонтологию. 3: 1–27. Получено 2010-08-29.
- ^ Хэй, Оливер Перри, 1902 г. Библиография и каталог ископаемых позвоночных Северной Америки.. Бюллетень Геологической службы США, № 117, Государственная типография. стр. 868
- ^ Карр, Томас Д. (2010). "Таксономическая оценка типовой серии Саркофаг альбертозавра и идентичность Tyrannosauridae (Dinosauria, Coelurosauria) в Альбертозавр костяной пласт из формации Хорсшу-Каньон (кампан – маастрихт, поздний мел ». Канадский журнал наук о Земле. 47 (9): 1213–1226. Bibcode:2010CaJES..47.1213C. Дои:10.1139 / e10-035.
- ^ а б c d е Карри, Филип Дж. (1998). «Возможное свидетельство стадного поведения тираннозавридов» (PDF). Гайя. 15: 271–277. Архивировано из оригинал (PDF) на 2009-03-26. Получено 2009-05-03. (не печатается до 2000 г.)
- ^ а б c d е Эриксон, Грегори М .; Маковицкий, Питер Дж .; Карри, Филип Дж .; Норелл, Марк А .; Yerby, Scott A .; Брошу, Кристофер А. (2004). «Гигантизм и сравнительные параметры жизненного цикла динозавров-тираннозавров» (PDF). Природа. 430 (7001): 772–775. Bibcode:2004Натура 430..772E. Дои:10.1038 / природа02699. PMID 15306807. S2CID 4404887. Архивировано из оригинал (PDF) на 2011-10-06. Получено 2010-08-29.
- ^ Eberth, David A .; Карри, Филип Дж. (2010). «Стратиграфия, седиментология и тафономия Альбертозавр костное дно (верхняя часть формации каньона Подкова; маастрихт), южная часть Альберты, Канада ". Канадский журнал наук о Земле. 47 (9): 1119–1143. Bibcode:2010CaJES..47.1119E. Дои:10.1139 / e10-045.
- ^ Карпентер, К., 1992, "Тираннозавриды (динозавры) Азии и Северной Америки", В: Н. Матир и П.-Дж. Чен (ред.) Аспекты неморской геологии мелового периода. China Ocean Press, Пекин, Китай, стр. 250–268.
- ^ Парки, Уильям А. (1928). "Альбертозавр arctunguis, новый вид тернистых динозавров из формации Эдмонтон в Альберте ". Исследования Университета Торонто, серия геологических исследований. 25: 1–42.
- ^ а б c d Рассел, Дейл А. (1970). «Тираннозавры из позднего мела западной Канады». Публикации по палеонтологии Национального музея естественных наук. 1: 1–34.
- ^ а б c d е ж грамм Хольц, Томас Р. (2004). «Тиранносавроидеа». В Weishampel, Дэвид Б.; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (Второе изд.). Беркли: Калифорнийский университет Press. С. 111–136. ISBN 978-0-520-24209-8.
- ^ Ламбе, Лоуренс М. (1914). "О новом роде и виде плотоядных динозавров из формации Белли-Ривер в Альберте, с описанием черепа Стефанозавр маргинальный с того же горизонта ». Оттавский натуралист. 28: 13–20.
- ^ а б c Риклефс, Роберт Э. (2007). «Старение тираннозавра». Письма о биологии. 3 (2): 214–217. Дои:10.1098 / rsbl.2006.0597. ЧВК 2375931. PMID 17284406.
- ^ а б Карр, Томас Д .; Уильямсон, Томас Э .; Швиммер, Дэвид Р. (2005). «Новый род и вид тираннозавроидов из позднемеловой (средний кампан) формации Демополис в Алабаме». Журнал палеонтологии позвоночных. 25 (1): 119–143. Дои:10.1671 / 0272-4634 (2005) 025 [0119: ANGASO] 2.0.CO; 2.
- ^ Рябинин, А. (1930). «[На русском]« О возрасте и фауне пластов динозавров на реке Амур ». Воспоминания Русского минералогического общества. 59 (2): 41–51.
- ^ Фон Хюене, Ф., 1932 Die fossile Reptil-Ordnung Saurischia: ihre Entwicklung und Geschichte. Monographie für Geologie und Palaeontologie, части I и II, сер. I, 4: 1-361
- ^ а б Пол, Грегори С. (1988). Хищные динозавры мира. Нью-Йорк: Саймон и Шустер. ISBN 978-0-671-61946-6.
- ^ Ольшевский, Георгий. (1995). «(Происхождение и эволюция тираннозавридов.)». Кёрюгаку Сайдзенсен (Дино Фронтлайн) (на японском языке). 9: 92–119.
- ^ а б c Карри, Филип Дж. (2003). «Аллометрический рост тираннозаврид (Dinosauria: Theropoda) из верхнего мела Северной Америки и Азии». Канадский журнал наук о Земле. 40 (4): 651–665. Bibcode:2003CaJES..40..651C. Дои:10.1139 / e02-083.
- ^ Мадер, Б .; Брэдли, Р. (1989). "Новое описание и уточненный диагноз синтипов монгольского тираннозавра. Alectrosaurus olseni". Журнал палеонтологии позвоночных. 9 (1): 41–55. Дои:10.1080/02724634.1989.10011737.
- ^ Мэтью, W.D .; Браун, Б. (1922). «Семейство Deinodontidae с указанием нового рода из мелового периода Альберты». Бюллетень Американского музея естественной истории. 46 (6): 367–385.
- ^ Кун, О., 1939 Saurischia - Fossilium catalogus I, Animalia, Pars 87. 's-Gravenhage, W. Junk, 1939, 124 стр.
- ^ Кристиансен, Пер; Фаринья, Ричард А. (2004). «Массовое предсказание у динозавров-теропод». Историческая биология. 16 (2–4): 85–92. Дои:10.1080/08912960412331284313. S2CID 84322349.
- ^ Пол, Г.С. (2016). Принстонский полевой путеводитель по динозаврам (Второе изд.). Издательство Принстонского университета. п. 112. ISBN 978-0-691-16766-4.
- ^ Молина-Перес и Ларраменди (2016). Records y curiosidades de los dinosaurios Terópodos y otros dinosauromorfos. Барселона, Испания: Ларусс. п. 266.
- ^ Талборн, Ричард А. (1982). «Скорости и походки динозавров». Палеогеография, палеоклиматология, палеоэкология. 38 (3–4): 227–256. Bibcode:1982ППП .... 38..227Т. Дои:10.1016/0031-0182(82)90005-0.
- ^ а б c Белл, Фил; Э. Кампионе, Николас; Скотт Персонс, Вт; Дж. Карри, Филип; Ларсон, Питер; Танке, Даррен; Т. Баккер, Роберт (2017-06-01). «Покровы тираннозавроидов демонстрируют противоречивые закономерности гигантизма и эволюции перьев». Письма о биологии. 13 (6): 20170092. Дои:10.1098 / рсбл.2017.0092. ЧВК 5493735. PMID 28592520.
- ^ Райхель, Мириам (2010). "Гетеродонность Саркофаг альбертозавра и тиранозавр Рекс: биомеханические последствия, полученные с помощью трехмерных моделей ". Канадский журнал наук о Земле. 47 (9): 1253–1261. Bibcode:2010CaJES..47.1253R. Дои:10.1139 / e10-063.
- ^ «Альбертозавр». В: Додсон, Питер; Бритт, Брукс; Карпентер, Кеннет; Forster, Catherine A .; Gillette, David D .; Норелл, Марк А .; Ольшевский, Георгий; Пэрриш, Дж. Майкл; И Weishampel, Дэвид Б. Эпоха динозавров. Линкольнвуд, Иллинойс: Publications International, Ltd., 1993. С. 106–107. ISBN 0-7853-0443-6.
- ^ а б c d е ж грамм Abler, W.L. 2001. Модель зубцов тираннозавра в виде пропилов и сверлений. п. 84–89. В: Мезозойская жизнь позвоночных. Под ред. Танке, Д. Х., Карпентер, К., Скрепник, М. В. Издательство Индианского университета.
- ^ а б Карри, Филип Дж.; Hurum, Jørn H; Сабат, Кароль (2003). «Строение черепа и эволюция в филогении тираннозаврид» (PDF). Acta Palaeontologica Polonica. 48 (2): 227–234. В архиве (PDF) из оригинала 26 марта 2009 г.. Получено 2009-05-03.
- ^ Хольц, Томас Р. (2005-09-20). "RE: Burpee Conference (LONG)". Получено 2007-06-18.
- ^ Лёвен, М.А.; Ирмис, Р.; Сертич, J.J.W.; Карри, П. Дж.; Сэмпсон, С.Д. (2013). Эванс, Дэвид С (ред.). «Эволюция динозавров-тиранов отслеживает подъем и падение океанов позднего мелового периода». PLoS ONE. 8 (11): e79420. Bibcode:2013PLoSO ... 879420L. Дои:10.1371 / journal.pone.0079420. ЧВК 3819173. PMID 24223179.
- ^ Эриксон, Грегори М .; Карри Роджерс, Кристи; Варриккио, Дэвид Дж .; Норелл, Марк .; Сюй, Син (2007). «Модели роста у высиживающих динозавров показывают время наступления половой зрелости у нептичьих динозавров и генезис птичьего состояния». Письма о биологии. 3 (5): 558–561. Дои:10.1098 / rsbl.2007.0254. ЧВК 2396186. PMID 17638674.
- ^ а б c Ли, Эндрю Х .; Вернинг, Сара (2008). «Половая зрелость у растущих динозавров не соответствует моделям роста рептилий». PNAS. 105 (2): 582–587. Дои:10.1073 / pnas.0708903105. ЧВК 2206579. PMID 18195356.
- ^ Бакли, Лиза Дж .; Ларсон, Дерек У .; Райхель, Мириам; Самман, Таня (2010). "Количественная оценка вариации зубов в одной популяции Саркофаг альбертозавра (Theropoda: Tyrannosauridae) и значение для идентификации изолированных зубов тираннозаврид ". Канадский журнал наук о Земле. 47 (9): 1227–1251. Bibcode:2010CaJES..47.1227B. Дои:10.1139 / e10-029.
- ^ а б c Роуч, Брайан Т .; Бринкман, Дэниел Т. (2007). "Переоценка совместной охоты стаей и общительности в Deinonychus antirrhopus и другие динозавры-тероподы, не являющиеся морскими животными ". Бюллетень музея естественной истории Пибоди. 48 (1): 103–138. Дои:10.3374 / 0079-032X (2007) 48 [103: AROCPH] 2.0.CO; 2.
- ^ Эриксон, Грегори М .; Карри, Филип Дж .; Inouye, Brian D .; Винн, Алиса А. (2010). "Пересмотренная таблица дожития и кривая выживаемости для Саркофаг альбертозавра основан на сборке массовых смертей на Сухом острове ". Канадский журнал наук о Земле. 47 (9): 1269–1275. Bibcode:2010CaJES..47.1269E. Дои:10.1139 / e10-051.
- ^ Eberth, David A .; МакКри, Ричард Т. (2001). «Были ли крупные тероподы стадными?». Журнал палеонтологии позвоночных. 21 (Приложение к 3 - Тезисы статей 61-го ежегодного собрания Общества палеонтологии позвоночных): 46A. Дои:10.1080/02724634.2001.10010852. S2CID 220414868.(только опубликованная аннотация)
- ^ Хорнер, Джон Р. (1997). "Поведение". В Карри, Филип Дж.; Падиан, Кевин (ред.). Энциклопедия динозавров. Сан-Диего: Academic Press. С. 45–50. ISBN 978-0-12-226810-6.
- ^ Максвелл, У. Десмонд; Остром, Джон Х. (1995). "Тафономия и палеобиологические последствия Тенонтозавр-Дейноних ассоциации ». Журнал палеонтологии позвоночных. 15 (4): 707–712. Дои:10.1080/02724634.1995.10011256.
- ^ Раат, Майкл А. (1990). «Морфологическая изменчивость мелких теропод и ее значение в систематике: данные Syntarsus rhodesiensis". В Карпентер, Кеннет; Карри, Филип Дж. (Ред.). Систематика динозавров: подходы и перспективы. Кембридж: Издательство Кембриджского университета. С. 91–105. ISBN 978-0-521-43810-0.
- ^ Кориа, Родольфо А.; Карри, Филип Дж. (2006). "A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina" (PDF). Геодиверситас. 28 (1): 71–118. Архивировано из оригинал (PDF) на 2009-03-26. Получено 2009-05-03.
- ^ Карри, Филип Дж.; Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; Murphy, Nate (2005). "An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA)". В Карпентер, Кеннет (ред.). The Carnivorous Dinosaurs. Блумингтон: Издательство Индианского университета. pp. 313–324. ISBN 978-0-253-34539-4.
- ^ Auffenberg, Walter (2000). The Behavioral Ecology of the Komodo Monitor. Гейнсвилл: Издательство Университета Флориды. ISBN 978-0-8130-0621-5.
- ^ Currie, Philip J.; Eberth, David A. (2010). "On gregarious behavior in Альбертозавр". Канадский журнал наук о Земле. 47 (9): 1277–1289. Bibcode:2010CaJES..47.1277C. Дои:10.1139/e10-072.
- ^ а б Wolff, Ewan D. S.; Salisbury, Steven W.; Horner, John R.; Varricchio, David J. (2009). Hansen, Dennis Marinus (ed.). "Common avian infection plagued the tyrant dinosaurs". PLOS ONE. 4 (9): e7288. Bibcode:2009PLoSO...4.7288W. Дои:10.1371/journal.pone.0007288. ЧВК 2748709. PMID 19789646.
- ^ Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331–336.
- ^ Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337–363.
- ^ Bell, Phil R. (2010). "Palaeopathological changes in a population of Саркофаг альбертозавра from the Upper Cretaceous Horseshoe Canyon Formation of Alberta, Canada". Канадский журнал наук о Земле. 47 (9): 1263–1268. Bibcode:2010CaJES..47.1263B. Дои:10.1139/e10-030.
- ^ а б c Эберт, Дэвид А. (1997). "Edmonton Group". В Карри, Филип Дж.; Падиан, Кевин (ред.). Энциклопедия динозавров. Сан-Диего: Academic Press. С. 199–204. ISBN 978-0-12-226810-6.
- ^ Ларсон, Дерек У .; Brinkman, Donald B.; Bell, Phil R. (2010). "Faunal assemblages from the upper Horseshoe Canyon Formation, an early Maastrichtian cool-climate assemblage from Alberta, with special reference to the Саркофаг альбертозавра bonebed". Канадский журнал наук о Земле. 47 (9): 1159–1181. Bibcode:2010CaJES..47.1159L. Дои:10.1139/e10-005.
- ^ Лонгрич, Николас Р .; Карри, Филип Дж. (2009). "Albertonykus borealis, новый альваресзавр (Dinosauria: Theropoda) из раннего маастрихта провинции Альберта, Канада: значение для систематики и экологии Alvarezsauridae ". Меловые исследования. 30 (1): 239–252. Дои:10.1016 / j.cretres.2008.07.005.
внешняя ссылка
 СМИ, связанные с Альбертозавр в Wikimedia Commons
СМИ, связанные с Альбертозавр в Wikimedia Commons













