Цератозавр - Ceratosaurus
| Цератозавр | |
|---|---|
 | |
| В ролях Цератозавр от Кливленд Ллойд Карьер, выставленные на Музей естественной истории Юты | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Ceratosauridae |
| Род: | †Цератозавр Марш, 1884 |
| Типовой вид | |
| †Ceratosaurus nasicornis Марш, 1884 г. | |
| Другие виды | |
| Синонимы | |
| |
Цератозавр /ˌsɛrəтoʊˈsɔːrəs/ (из Греческий κέρας / κέρατος, керас / кератос что означает "рог" и σαῦρος sauros означает «ящерица») был плотоядным теропод динозавр в Поздняя юра период (Кимериджский к Титонский ). Этот род впервые описан в 1884 году американским палеонтологом. Отниэль Чарльз Марш основанный на почти полном скелете, обнаруженном в Гарден Парк, Колорадо, в породах, принадлежащих Формация Моррисон. В типовой вид является Ceratosaurus nasicornis.
Образец Garden Park остается наиболее полным скелетом, известным из рода, и с тех пор было описано лишь несколько дополнительных экземпляров. Два дополнительных вида, Ceratosaurus dentisulcatus и Ceratosaurus magnicornis, были описаны в 2000 г. по двум фрагментарным скелетам из Кливленд-Ллойд Карьер из Юта и из окрестностей Фруита, Колорадо. В срок действия Однако эти дополнительные виды были подвергнуты сомнению, и все три скелета, возможно, представляют разные стадии роста одного и того же вида. В 1999 г. было сообщено об обнаружении первого экземпляра молоди. С 2000 г. частичный экземпляр был раскопан и описан из Формация Лориньян из Португалия, что свидетельствует о присутствии рода за пределами Северной Америки. Сообщалось также о фрагментарных останках Танзания, Уругвай, и Швейцария, хотя их отнесение к Цератозавр в настоящее время не принимается большинством палеонтологи.
Цератозавр был тероподом среднего размера. Исходный образец оценивается в 5,3 м (17 футов) или 5,69 м (18,7 фута) в длину, в то время как образец, описанный как C. dentisulcatus был больше, около 7 м (23 фута) в длину. Цератозавр был характеризует глубокими челюстями, которые поддерживали пропорционально очень длинные лезвийные зубы, выступающий гребневидный рог на средней линии морды и пару рогов над глазами. Передние конечности были очень короткими, но оставались полностью функциональными; на руке было четыре пальца. Хвост был глубоким сверху вниз. Ряд малых остеодермы (кости кожи) присутствовали посередине шеи, спины и хвоста. Дополнительные остеодермы присутствовали в неизвестных местах на теле животного.
Цератозавр дает название Цератозаврия, а клады динозавров-теропод, которые рано отклонились от эволюционной линии, ведущей к современному птицы. Некоторые палеонтологи предположили, что внутри Ceratosauria он наиболее близок к Genyodectes из Аргентины, у которого общие сильно удлиненные зубы. Геологически более древний род Процератозавр из Англия, хотя первоначально описывался как предполагаемый антецедент Цератозавр, позже выяснилось, что они не связаны. Цератозавр разделял свою среду обитания с другими крупными родами теропод, включая Торвозавр и Аллозавр, и было высказано предположение, что эти тероподы занимали разные экологические ниши уменьшить конкуренцию. Цератозавр возможно, охотились на динозавров, питающихся растениями, хотя некоторые палеонтологи предполагали, что они охотились на водную добычу, например, на рыбу. Носовой рог, вероятно, не использовался в качестве оружия, как первоначально предлагал Марш, но, скорее всего, использовался исключительно для демонстрации.
Описание

Цератозавр последовал за план тела типичен для крупных динозавров-теропод.[1] А двуногий, он передвигался на мощных задних лапах, а его руки были уменьшены в размерах. Образец USNM 4735, первый обнаруженный скелет и голотип из Ceratosaurus nasicornis, был индивидуумом длиной 5,3 м (17 футов) или 5,69 м (18,7 футов), согласно отдельным источникам.[2]:115[3] Выросло ли это животное полностью, не ясно.[4]:66 Отниэль Чарльз Марш в 1884 г., предположил, что этот образец весил примерно вдвое меньше современного Аллозавр.[5] В более поздних отчетах это было пересмотрено до 418 кг (922 фунтов), 524 кг (1155 фунтов) или 670 кг (1480 фунтов).[6] Еще три скелета, обнаруженные во второй половине 20 века, были значительно больше. Первый из них, UMNH VP 5278, по неофициальной оценке Джеймса Мэдсена, имел длину около 8,8 м (29 футов),[7] но позже был оценен в 7 м (23 фута) в длину.[8] Его вес был рассчитан на 980 кг (2160 фунтов), 452 кг (996 фунтов) и 700 кг (1540 фунтов) в отдельных работах.[3][8][9] Второй скелет, MWC 1, был несколько меньше UMNH VP 5278 и мог весить 275 кг (606 фунтов).[9] Третий, еще не описанный, образец BYUVP 12893 был заявлен как самый крупный из обнаруженных, хотя оценки не были опубликованы.[10]:192 Другой образец (ML 352), обнаруженный в Португалии в 2000 году, оценивался в длину 6 м (20 футов) и вес в 600 кг (1320 фунтов).[8]

Точное количество позвонков неизвестно из-за нескольких щелей в позвоночнике Ceratosaurus nasicornis голотип. Не менее 20 позвонков образовали шею и спину перед крестец. В средней части шеи центр (тела) позвонков были такими же длинными, как и высокие, в то время как в передней и задней частях шеи центры были короче своей высоты. Восходящая проекция нервные отростки были сравнительно большими и в спинных (задних) позвонках были такой же высоты, как и тела позвонков. Крестец, состоящий из шести сросшихся крестцовые позвонки, был выгнут вверх, с сильно уменьшенными в высоте центрами позвонков в средней части, как в некоторых других цератозавры.[4]:55–58 Хвост составлял около 50 хвостовые позвонки и составлял около половины всей длины животного; на голотипе он оценивается в 2,84 м (9,33 фута).[5][2]:115 Хвост был глубоким сверху вниз из-за высоких нервных шипов и удлиненной формы. шевроны, кости, расположенные ниже центральных позвонков. Как и у других динозавров, он уравновешивал тело и содержал массивные каудофеморальный мышца, которая отвечала за толчок вперед во время движения, тянет верхнюю часть бедра назад, когда контракт.[4]:55–58

В лопатка (лопатка) слилась с коракоидный, образуя единую кость без видимой границы между двумя исходными элементами.[4]:58 В C. nasicornis был обнаружен голотип с сочлененной левой передней конечностью, включая неполный манус (рука). Несмотря на то что расчлененный Во время подготовки из окаменелости была сделана слепка, чтобы задокументировать первоначальное относительное положение костей. Кости запястья не были известны ни по одному экземпляру, что побудило некоторых авторов предположить, что они потерялись в этом роде. В статье 2016 года Мэтью Каррано и Джона Чуанььер предположили, что один или несколько хрящевой (не костные) запястья, вероятно, присутствовали, о чем свидетельствует зазор между костями предплечья и пястной кости, а также текстура поверхности внутри этого зазора, видимая в гипсе.[11] В отличие от большинства другихполученный теропод, у которых на каждой кисти было всего по три пальца (цифры I – III), Цератозавр сохранены четыре цифры, цифра IV уменьшена в размерах. Первый и четвертый пястные кости были короткими, а второй был немного длиннее третьего. Пястные кости и особенно первая фаланги были пропорционально очень короткими, в отличие от большинства других базальный теропод. На голотипе сохранились только первые фаланги пальцев II, III и IV; общее количество фаланг и ногтевые (кости когтей) неизвестно. Анатомия I пястной кости указывает на то, что изначально фаланги присутствовали и на этом пальце. В пес (ступня) состояла из трех опорных пальцев, пронумерованных II – IV. Цифра I, которая у теропод обычно сводится к dewclaw что не касается земли, не сохранилось в голотипе. Марш в своем первоначальном описании 1884 года предположил, что эта цифра была потеряна в Цератозавр, но Чарльз Гилмор в своей монографии 1920 г. отметил область прикрепления на втором плюсневой демонстрируя наличие этой цифры.[2]:112
Уникально среди теропод, Цератозавр обладал маленькими, удлиненными и неправильной формы остеодермы (кожные кости) по средней линии его тела. Такие остеодермы были обнаружены над нервными отростками шейных позвонков 4 и 5, а также хвостовых позвонков с 4 по 10 и, вероятно, образовывали непрерывный ряд, который мог тянуться от основания черепа до большей части хвоста. Как предположил Гилмор в 1920 году, их положение в матрице горных пород, вероятно, отражает их точное положение в живом животном. Было обнаружено, что остеодермы над хвостом отделены от нервных шипов на 25 мм (0,98 дюйма) до 38 мм (1,5 дюйма), что, возможно, составляет кожу и мышцы, присутствующие между ними, в то время как мышцы шеи были гораздо ближе к нервным шипам. Помимо средней линии тела, кожа содержала дополнительные остеодермы, на что указывает крупная четырехугольная пластинка размером 58 мм (2,3 дюйма) на 70 мм (2,8 дюйма), обнаруженная вместе с голотипом; положение этой пластины на теле неизвестно.[2]:113–114 Образец UMNH VP 5278 был также обнаружен с рядом остеодерм, которые были описаны как аморфные по форме. Хотя большинство из них косточки были найдены на расстоянии не более 5 м от скелета, они не были напрямую связаны с какими-либо позвонками, в отличие от C. nasicornis голотип, поэтому их исходное положение на теле не может быть определено по этому экземпляру.[12]:32
Череп

В череп был довольно большим по сравнению с остальной частью его тела.[1] Его длина составляет 55 см (22 дюйма). C. nasicornis голотип, измеренный от кончика морды до затылочный мыщелок, который соединяется с первым шейным позвонком.[2]:88 Ширину этого черепа трудно реконструировать, так как он сильно искажен, а реконструкция Гилмора 1920 года была позже признана слишком широкой.[13] Довольно полный череп образца MWC 1, по оценкам, имел длину 60 см (24 дюйма) и ширину 16 см (6,3 дюйма); этот череп был несколько более вытянутым, чем у голотипа.[12]:3 Задняя часть черепа была построена легче, чем у некоторых других более крупных теропод, из-за обширных отверстий в черепе, однако челюсти были глубокими, чтобы поддерживать пропорционально большие зубы.[3]:277 В слезная кость формируется не только задний край анторбитальное окно, большое отверстие между глазом и костлявая ноздря, но также и часть его верхнего поля, в отличие от элементов связанных Abelisauridae. В квадратная кость, который был соединен с нижней челюстью своим нижним концом для образования челюстного сустава, был наклонен так, что челюстной сустав был смещен назад по отношению к затылочному мыщелку. Это также привело к расширению базы латеральное височное отверстие, большое отверстие за глазами.[4]:53
Наиболее отличительной особенностью был выступающий рог, расположенный на средней линии черепа за костными ноздрями, который образовался из сросшихся выступов левого и правого. носовые кости.[2]:82 По окаменелостям известно только костяное ядро рога - у живого животного это ядро могло бы поддерживать ороговевший оболочка. В то время как основание рогового ядра было гладким, его верхние две трети были морщинистыми и выстланы бороздками, которые могли содержать кровеносный сосуд когда жив. В голотипе ядро рога имеет длину 13 см (5,1 дюйма) и ширину 2 см (0,79 дюйма) у основания, но быстро сужается до 1,2 см (0,47 дюйма) дальше вверх; его высота составляет 7 см (2,8 дюйма).[2]:82 Он длиннее и ниже в черепе MWC 1.[12]:3 У живого животного рог, вероятно, был бы более удлиненным из-за его ороговевшей оболочки.[14] За носовым рогом носовые кости образовывали овальную бороздку; и эта борозда, и носовой рог служат особенности различать Цератозавр из родственных родов.[10]:192 Помимо большого носового рога, Цератозавр обладал меньшими полукруглыми костными выступами перед каждым глазом, подобными таковым у Аллозавр. Эти гребни образованы слезными костями.[9] У молоди все три рога были меньше, чем у взрослых, и две половинки сердцевины носового рога еще не срослись.[15]
В предчелюстные кости, который образовывал кончик рыла, имел всего по три зуба с каждой стороны, меньше, чем у большинства других теропод.[4]:52 В верхнечелюстные кости верхней челюсти на голотипе выстланы по 15 пластинчатых зубов с каждой стороны. Первые восемь из этих зубов были очень длинными и крепкими, но начиная с девятого зуба они постепенно уменьшались в размерах. Как типично для теропод, они имели мелкие зубчатый ребер, которых в голотипе было около 10 зубчики на 5 мм (0,20 дюйма).[2]:92 Образец MWC 1 показал всего от 11 до 12, а образец UMNH VP 5278 - 12 зубов в каждой верхней челюсти; у последнего экземпляра зубы были более крепкими и более изогнутыми.[12]:3,27 Во всех экземплярах зубные коронки верхних челюстей были исключительно длинными. В образце УМНХ ВП 5278 они имели длину до 9,3 см (3,7 дюйма), что равно минимальной высоте нижней челюсти. На голотипе они имеют длину 7 см (2,8 дюйма), что даже превышает минимальную высоту нижней челюсти. У других теропод сравнимая длина зубов известна только по возможно близкородственным Genyodectes.[16] Напротив, некоторые представители Abelisauridae имеют очень короткие коронки зубов.[4]:92 В голотипе каждая половина зубной, зубная кость нижняя челюсть, был оснащен 15 зубами, которые, однако, плохо сохранились. Оба образца MWC 1 и UMNH VP 5278 показывают только 11 зубов в каждой зубной части, которые, как показывает последний образец, были немного более прямыми и менее прочными, чем зубы верхней челюсти.[12]:3,21
История открытия
Образец голотипа C. nasicornis
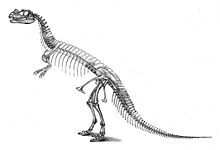
Первый образец, голотип USNM 4735, был обнаружен и раскопан фермером Маршаллом Паркером Фельчем в 1883 и 1884 годах.[17] Найденный в сочленении, с костями, все еще соединенными друг с другом, он был почти готов, включая череп. Значительные недостающие части включают неизвестное количество позвонков; все, кроме последних ребер туловища; в плеча (кости плеча); дистальные кости пальцев обеих рук; большая часть правой передней конечности; большая часть левой задней конечности; и большая часть ног.[2]:77 Образец был найден заключенным в твердый песчаник; череп и позвоночник были сильно деформированы во время окаменелость.[2]:2,114 Место открытия, расположенное в Садовый парк область к северу от Каньон-Сити, Колорадо, и известный как карьер Фелч 1, считается одним из самых богатых ископаемых Формация Моррисон. Многочисленные окаменелости динозавров были обнаружены в этом карьере еще до открытия Цератозавр, в первую очередь почти полный образец Аллозавр (USNM 4734) в 1883 и 1884 годах.[2]:7,114
После раскопок образец был отправлен на Музей естественной истории Пибоди в Новый рай, где он был изучен Маршем, который описал его как новый род и вид Ceratosaurus nasicornis в 1884 г.[5][2]:114 Название Цератозавр переводится как «рог ящерицы» (от Греческий κερας / κερατος, керас / кератос- «рог» и σαυρος /sauros- «ящерица»),[7] и носикорнис с "носовым рогом" (от латинский нос- «нос» и Cornu-"Рог").[18] Учитывая полноту экземпляра, недавно описанный род был в то время самым известным тероподом, обнаруженным в Америке. В 1898 и 1899 гг. Экземпляр был передан в Национальный музей естественной истории в Вашингтон, округ Колумбия вместе со многими другими окаменелостями, первоначально описанными Маршем. Только часть этого материала была полностью подготовлена, когда прибыла в Вашингтон; последующая подготовка длилась с 1911 по конец 1918 года. Упаковка и доставка из Нью-Хейвена в Вашингтон причинили некоторый ущерб машине. Цератозавр образец.[2]:2,114 В 1920 г. Чарльз Гилмор опубликовал обширное переописание этого и других образцов теропод, полученных из Нью-Хейвена, включая почти полное Аллозавр экземпляр извлечен из того же карьера.[2]:2

В статье 1892 года Марш опубликовал первую реконструкцию скелета Цератозавр, на котором изображено животное ростом 22 фута (6,7 м) в длину и 12 футов (3,7 м) в высоту.[1] Как отмечал Гилмор в 1920 году, туловище в этой реконструкции было изображено слишком длинным, включая по крайней мере шесть спинных позвонков. Эта ошибка повторялась в нескольких последующих публикациях, в том числе в первой реконструкции жизни, которую в 1899 году нарисовал Фрэнк Бонд под руководством Чарльз Р. Найт, но не публиковался до 1920 года. Более точная реконструкция жизни, опубликованная в 1901 году, была произведена Джозеф М. Глисон, снова под наблюдением Найта. Голотип был установлен Гилмором в 1910 и 1911 годах, и с тех пор выставлялся в Национальном музее естественной истории. Самые ранние реконструкции показывают Цератозавр в вертикальном положении, волоча хвостом по земле.[2]:115–116 Гилмор на голотипе, напротив, опередил свое время:[3]:276 Вдохновленный костями верхней части бедра, которые были обнаружены под углом к голени, он изобразил лошадь как бегущее животное с горизонтальной, а не вертикальной позой и хвостом, который не касался земли. Из-за сильного сплющивания окаменелостей Гилмор установил образец не как отдельно стоящий скелет, а как барельеф внутри искусственной стены.[2]:114 Поскольку кости были частично встроены в бляшку, доступ к исследованиям был ограничен. В ходе обновления экспозиции динозавров в музее в период с 2014 по 2019 год образец был демонтирован и освобожден от обшивки.[19][20] На новой выставке, которая откроется в 2019 году, монтировку планируется заменить отдельно стоящей, а оригинальные кости будут храниться в коллекции музея, чтобы обеспечить полный доступ для ученых.[20]

Дополнительные находки в Северной Америке
После открытия голотипа C. nasicornis, значительный Цератозавр Эта находка была сделана только в начале 1960-х годов, когда палеонтолог Джеймс Мэдсен и его команда обнаружили фрагментарный, разрозненный скелет, включая череп (UMNH VP 5278), в карьере динозавров Кливленд-Ллойд в штате Юта. Эта находка представляет собой одну из самых известных Цератозавр образцы.[12]:21 Второй шарнирный образец, включающий череп (MWC 1), был обнаружен в 1976 году Тором Эриксоном, сыном палеонтолога Ланса Эриксона, недалеко от Фруита, Колорадо.[7] Достаточно полный экземпляр, у него отсутствуют нижняя челюсть, предплечья и гастралия. Череп, хотя и довольно полный, был обнаружен разрозненным и сильно уплощенным с боков. Несмотря на то, что он был крупным, он еще не достиг взрослого размера, о чем свидетельствуют открытые швы между костями черепа.[12]:2–3 Научно точные трехмерные реконструкции черепа для использования в музейных экспонатах были произведены с использованием сложного процесса, включающего формование и отливку отдельных оригинальных костей, исправление деформаций, реконструкцию недостающих частей, сборку слепков костей в их правильное положение и покраска под оригинальный цвет костей.[21]
Образцы как Фруита, так и Кливленд-Ллойд были описаны Мэдсеном и Сэмюэл Пол Уэллс в монографии 2000 г., при этом экземпляр из Юты был отнесен к новому виду C. dentisulcatus и образец Колорадо к новому виду C. magnicornis.[12] Название зубной камень относится к параллельным бороздкам, имеющимся на внутренней стороне предчелюстных зубов и первых трех зубов нижней челюсти в этом образце; магникорнис указывает на больший носовой рог.[12]:2,21 В срок действия обоих видов, однако, подвергался сомнению в последующих публикациях. Брукс Бритт и его коллеги в 2000 году утверждали, что C. nasicornis голотип был фактически молодой особью, причем два более крупных вида представляли взрослую особь одного вида.[22] Оливер Раухут в 2003 г. и Мэтью Каррано и Скотт Сэмпсон в 2008 г. рассмотрели анатомические различия, приведенные Мэдсеном и Уэллсом, чтобы поддержать эти дополнительные виды для представления онтогенетический (возрастные) или индивидуальные вариации.[23][10]:192

Еще один образец (BYUVP 12893) был обнаружен в 1992 году в карьере Агатовой котловины к юго-востоку от г. Мур, Юта, но все еще ждет описания. Образец, который считается самым крупным из представителей этого рода, включает переднюю половину черепа, семь фрагментов тазовых спинных позвонков, а также сочлененный таз и крестец.[10]:192[12]:36 В 1999 году Бритт сообщил об открытии Цератозавр скелет молодой особи. Обнаружен в Костяной карьер в Вайоминге он на 34% меньше, чем C. nasicornis голотип и состоит из полного черепа, а также 30% остальной части скелета, включая полный таз.[15]
Помимо этих пяти скелетных находок, фрагментарные Цератозавр останки были зарегистрированы в различных местах из стратиграфических зон 2 и 4-6 формации Моррисон,[24] включая некоторые из основных ископаемых участков формации. Национальный памятник динозаврам, Юта, получена изолированная правая предчелюстная кость (номер образца DNM 972); о большой лопатке (скапулокоракоид) сообщалось от Como Bluff в Вайоминг. Другой образец происходит из Карьер Dry Mesa, Колорадо, и включает левый скапулокоракоид, а также фрагменты позвонков и костей конечностей. В карьере Майгатт Мур, штат Колорадо, этот род известен по зубам.[12]:36
Находки за пределами Северной Америки
С 1909 по 1913 год немецкие экспедиции Берлин Museum für Naturkunde обнаружил разнообразную фауну динозавров из Формация Тендагуру в Германская Восточная Африка, в том, что сейчас Танзания.[25] Хотя обычно считается самым важным местом обитания африканских динозавров,[25] крупные динозавры-тероподы известны лишь по немногим и очень фрагментарным останкам.[26] В 1920 году немецкий палеонтолог Вернер Яненш присвоено несколько спинных позвонков из карьера «TL» Цератозавр, так как Цератозавр sp. (неопределенных видов). В 1925 году Яненш назвал новый вид Цератозавр, C. roechlingi, основанный на фрагментарных останках из карьера «Mw», включающих квадратную кость, малоберцовую кость, фрагменты хвостовых позвонков и другие фрагменты. Этот экземпляр происходит от особи, значительно крупнее, чем C. nasicornis голотип.[26]

В своей монографии 2000 года Мэдсен и Уэллс подтвердили принадлежность этих находок к Цератозавр. Кроме того, они отнесли несколько зубов к роду, который первоначально был описан Яненшем как возможный вид Лаброзавр, Лаброзавр (?) Stechowi.[12] Другие авторы поставили под сомнение принадлежность какой-либо из находок Тендагуру к Цератозавр, отметив, что ни один из этих экземпляров не обнаруживает признаков, диагностических для этого рода.[27][4]:66[10]:192[28] В 2011 году Раухут обнаружил как C. roechlingi и Лаброзавр (?) Stechowi быть возможными цератозавридами, но обнаружили, что они не являются диагностическими на уровне рода, и поэтому обозначили их как Номина Дубия (сомнительные имена).[28] В 1990 году Тимоти Роу и Жак Готье упомянул еще один Цератозавр виды из Тендагуру, Ceratosaurus ingens, который якобы был установлен Яненшем в 1920 году и основывался на 25 отдельных очень больших зубах до 15 см (5,9 дюйма) в длину.[27][26] Однако Яненш отнес этот вид к Мегалозавр, а не Цератозавр; следовательно, это имя может быть простой ошибкой копирования.[12]:37[26] Раухут в 2011 году показал, что Мегалозавр Ingens не был тесно связан ни с Мегалозавр или же Цератозавр, но, возможно, представляет собой кархародонтозаврид, вместо.[28]
В 2000 и 2006 гг. Палеонтологи под руководством Октавио Матеус описал находку из Формация Лориньян в центре-западе Португалия (ML 352) как новый образец Цератозавр, состоящий из правого бедренная кость (верхняя кость бедра), левая большеберцовая кость (большеберцовая кость) и несколько изолированных зубов, найденных на скалах пляжа Валмитан, между муниципалитетами Lourinhã и Торриш Ведраш.[29][30] Кости были обнаружены в мелкозернистых песчаниках от желтого до коричневого цвета, которые были отложены реками в виде пойменных отложений и относятся к нижним уровням пачки Порто-Ново, которая считается поздней. Кимериджский в возрасте. Дополнительные кости этого человека (SHN (JJS) -65), включая левую бедренную кость, правую большеберцовую кость и частично левую малоберцовая кость (кость голени), с тех пор были обнажены из-за прогрессирования эрозия обрыва. Хотя изначально эти дополнительные элементы были частью частной коллекции, эти дополнительные элементы стали официально храниться после того, как частная коллекция была передана Sociedade de História Natural в Торрес-Ведрас, и были подробно описаны в 2015 году.[31] Экземпляр отнесен к виду Ceratosaurus dentisulcatus Матеус и его коллеги в 2006 году.[30] Обзор 2008 года, проведенный Каррано и Сэмпсоном, подтвердил назначение Цератозавр, но пришел к выводу, что отнесение к какому-либо конкретному виду в настоящее время невозможно.[10]:192 В 2015 году Элизабет Малафайя и его коллеги поставили под сомнение обоснованность C. dentisulcatus, присвоил образец Цератозавр aff. Ceratosaurus nasicornis.[31]
В других сообщениях упоминается один зуб, найденный в Мутье, Швейцария. Первоначально названный Яненшем в 1920 году как Лаброзавр мериани, позже зуб был назначен Цератозавр sp. (неизвестного вида) Мэдсена и Уэллса.[12]:35–36 В 2008 году Матиас Сото и Даниэль Переа описали зубы из Формация Такуарембо в Уругвай, включая предполагаемую коронку предчелюстного зуба.На его внутренней стороне видны вертикальные бороздки, а на переднем крае отсутствуют зубчики; эти особенности в этой комбинации известны только из Цератозавр. Авторы, однако, подчеркнули, что поручение Цератозавр невозможно, так как останков мало, и, кроме того, обратите внимание, что отнесение европейских и африканских материалов к Цератозавр следует рассматривать с осторожностью.[32] В 2020 году Сото и его коллеги описали дополнительные Цератозавр зубы из того же образования, что дополнительно подтверждает их более раннюю интерпретацию.[33]
Классификация

В его первоначальном описании Ceratosaurus nasicornis голотипа и последующих публикаций Марш отметил ряд характеристики которые не были известны у всех других известных в то время теропод.[10]:185 Две из этих черт, сросшийся таз и сросшийся плюсневой кость, были известны у современных птиц и, по словам Марша, ясно демонстрируют тесную связь между последними и динозаврами.[34] Чтобы отделить род от Аллозавр, Мегалозавр, и целурозавры, Марш сделал Цератозавр единственный член как нового семья, то Ceratosauridae, и новый инфраотряд, цератозаврия.[10]:185 Это было поставлено под сомнение в 1892 г. Эдвард Дринкер Коуп, Соперник Марша в Bone Wars, которые утверждали, что такие отличительные особенности, как носовой рог, просто показывают, что C. nasicornis был отдельным видом, но этого было недостаточно, чтобы оправдать отдельный род. Следовательно, он назначил C. nasicornis к роду Мегалозавр, создавая новую комбинацию Мегалозавр носикорнис.[35]
Несмотря на то что Цератозавр был сохранен как отдельный род во всех последующих анализах,[2]:76 его отношения оставались спорными в течение следующего столетия. И Ceratosauridae, и Ceratosauria не получили широкого признания, были идентифицированы лишь несколько и малоизвестных дополнительных членов. С годами отдельные авторы классифицировали Цератозавр в пределах Deinodontidae, то Megalosauridae, то Целурозаврия, то Карнозаврия, а Deinodontoidea.[12]:2 В своей редакции 1920 года Гилмор утверждал, что этот род был наиболее базальным тероподом, известным после Триасовый, поэтому не были тесно связаны с какими-либо другими современными тероподами, известными в то время; таким образом, он имеет собственное семейство Ceratosauridae.[2]:76 Так продолжалось до создания кладистический анализ однако в 1980-х годах первоначальное заявление Марша о Ceratosauria как об отдельной группе получило поддержку. В 1985 году недавно открытые южноамериканские роды Абелизавр и Карнотавр оказались тесно связаны с Цератозавр. Готье в 1986 году признал Coelophysoidea быть тесно связанным с Цератозавр, хотя согласно последним исследованиям эта клада выходит за рамки Ceratosauria. С тех пор были признаны многие дополнительные члены Ceratosauria.[10]:185
Ceratosauria рано отделилась от эволюционной линии, ведущей к современным птицам, поэтому считается базальный внутри теропод.[36] Сама цератозаврия содержит группу производных (небазальных) членов семейств Noasauridae и Abelisauridae, заключенные в скобки внутри клады Абелизавройда, а также ряд базальных членов, таких как Элафрозавр, Deltadromeus, и Цератозавр. Положение Цератозавр внутри базальных цератозавров обсуждается. Некоторые анализы рассмотрели Цератозавр как наиболее производный из базальных членов, образующих сестринский таксон Абелизавройда.[10]:187[37] Оливер Раухут в 2004 году предложил Genyodectes как родственный таксон Цератозавр, так как оба рода характеризуются исключительно длинными зубами на верхней челюсти.[16] Раухут сгруппированы Цератозавр и Genyodectes внутри семейства Ceratosauridae,[16] за которым последовали несколько более поздних отчетов.[38][39][40][14]
Шуо Ван и его коллеги в 2017 году пришли к выводу, что Noasauridae не гнездились внутри Abelisauroida, как предполагалось ранее, а вместо этого были более базальными, чем Цератозавр. Поскольку ноазавриды использовались в качестве фиксированной точки для определения клад Abelisauroida и Abelisauridae, эти клады, следовательно, будут включать гораздо больше таксонов по определению, включая Цератозавр. В последующем исследовании 2018 года Рафаэль Делькур согласился с этими результатами, но указал, что, как следствие, Abelisauroida необходимо заменить на более старый синоним Ceratosauroida, который до сих пор использовался редко. Для Abelisauridae Делькур предложил новое определение, исключающее Цератозавр, что позволяет использовать имя в его традиционном смысле. Ван и его коллеги также обнаружили, что Цератозавр и Genyodectes образуют кладу с аргентинским родом Эоабелизавр.[40] Делькур использовал название Ceratosauridae для обозначения этой же клады и предложил определить Ceratosauridae как содержащие все таксоны, которые более тесно связаны с Цератозавр чем к абелизавриду Карнотавр.[14]

Следующее кладограмма показывая отношения Цератозавр основан на филогенетический Анализ, проведенный Диего Полом и Оливером Раухутом в 2012 году:[38]
| Цератозаврия |
| |||||||||||||||||||||||||||||||||||||||||||||
Череп из Средняя юра в Англии, по-видимому, имеет носовой рог, похожий на рог Цератозавр. В 1926 г. Фридрих фон Хюене описал этот череп как Процератозавр (что означает "до Цератозавр"), предполагая, что это был предшественник позднеюрского периода. Цератозавр.[41] Сегодня, Процератозавр считается базальным членом Тираннозавройда, гораздо более производная клада динозавров-теропод;[42] носовой рог, следовательно, развился независимо у обоих родов.[10]:185 Оливер Раухут и его коллеги в 2010 году сгруппировали Процератозавр внутри своей семьи Proceratosauridae. Эти авторы также отметили, что носовой рог сохранился не полностью, что открывает возможность того, что он представляет собой переднюю часть более обширного гребня головы, как это видно у некоторых других процедурных заврид, таких как Гуаньлун.[42]
Палеобиология
Экология и кормление

Внутри формации Моррисон, Цератозавр окаменелости часто встречаются вместе с таковыми других крупных теропод, в том числе мегалозаврид Торвозавр и аллозаврид Аллозавр. В районе Гарден-Парк в Колорадо, кроме Цератозавр, окаменелости, приписываемые Аллозавр. Карьер Dry Mesa в Колорадо, а также карьер Кливленд-Ллойд и Национальный памятник динозавров в штате Юта содержат останки по крайней мере трех крупных теропод: Цератозавр, Аллозавр, и Торвозавр.[13][43] Аналогичным образом, в Комо-Блафф и близлежащих районах в Вайоминге были обнаружены останки Цератозавр, Аллозавр, и по крайней мере один крупный мегалозаврид.[44] Цератозавр был редким элементом фауны теропод; его численно меньше Аллозавр в среднем от 7,5 к 1 на сайтах, где они встречаются одновременно.[45]
Несколько исследований попытались объяснить, как эти сочувствующий виды могли уменьшить прямую конкуренцию. Дональд Хендерсон в 1998 году утверждал, что Цератозавр совместно с двумя отдельными потенциальными видами Аллозавр, которые он обозначил как «морфы»: морф с укороченной мордой, высоким и широким черепом и короткими, выступающими назад зубами и морф, характеризующийся более длинной мордой, нижней частью черепа и длинными вертикальными зубами. Вообще говоря, чем больше сходство между симпатрическими видами в отношении их морфологии, физиологии и поведения, тем более интенсивной будет конкуренция между этими видами. Хендерсон пришел к выводу, что коротконорый Аллозавр морфинг занимал другой экологическая ниша от морфа с длинным носом и Цератозавр: Более короткий череп в этой морфе уменьшил бы изгибающие моменты, возникающие во время укуса, таким образом увеличив силу укуса, сравнимую с состоянием, наблюдаемым у кошек. Цератозавр и другие Аллозавр morph, однако, имел черепа с длинными мордами, которые лучше по сравнению с черепами собак: более длинные зубы использовались в качестве клыков для быстрого и резкого укуса, при этом сила укуса концентрировалась на меньшей площади из-за более узкого черепа. По словам Хендерсона, большое сходство в форме черепа между Цератозавр и длинноносый Аллозавр морфы указывают на то, что эти формы участвуют в прямой конкуренции друг с другом. Следовательно, Цератозавр мог быть вытеснен из мест обитания, где преобладали длиннорые морфы. В самом деле, Цератозавр очень редко встречается в карьере Кливленд-Ллойд, где обитают длинноносые Аллозавр морф, но, по-видимому, более распространен как в Гарден-парке, так и в карьере Сухой Мезы, где он встречается вместе с морфом с коротким носом.[13]

Кроме того, Хендерсон предположил, что Цератозавр мог избежать конкуренции, отдавая предпочтение разным объектам добычи; эволюция его чрезвычайно удлиненных зубов могла быть прямым результатом конкуренции с длинноносыми Аллозавр превращаться. Оба вида могли также отдавать предпочтение различным частям туши, выступая в качестве падальщиков. Удлиненные зубы Цератозавр могли служить визуальными сигналами, облегчающими распознавание представителей одного и того же вида или для других социальных функций. Кроме того, большой размер этих теропод имел тенденцию к снижению конкуренции, так как количество возможных объектов добычи увеличивается с размером.[13]
Фостер и Дэниел Чуре в исследовании 2006 года согласились с Хендерсоном в том, что Цератозавр и Аллозавр обычно жили в одних и тех же средах обитания и охотились на одни и те же типы добычи, поэтому, вероятно, имели разные стратегии питания, чтобы избежать конкуренции. По мнению этих исследователей, об этом также свидетельствуют разные пропорции черепа, зубов и передней конечности.[45] Различие между двумя Аллозавр морфы, однако, подверглись сомнению в некоторых более поздних исследованиях. Кеннет Карпентер, в исследовании 2010 г. обнаружили, что коротконорые особи Аллозавр из карьера Кливленд-Ллойд представляют собой случаи крайней индивидуальной изменчивости, а не отдельный таксон.[46] Более того, череп USNM 4734 из местности Гарден-Парк, который лег в основу анализа Хендерсоном морфы с коротким носом, позже оказался реконструированным слишком коротким.[47]
В исследовании 2004 г. Роберт Баккер и Гэри Бир предположил, что Цератозавр в первую очередь специализировалась на водной добыче, такой как двоякодышащая рыба, крокодилы и черепахи. Как показал статистический анализ выпавших зубов из 50 отдельных мест в районе Комо-Блафф и его окрестностях, зубы обоих Цератозавр и мегалозавриды были наиболее распространены в местообитаниях в и вокруг водных источников, таких как влажные поймы, окраины озер и болота. Цератозавр также изредка встречался в наземных точках. Однако аллозавриды были одинаково распространены в наземных и водных средах обитания. Из этих результатов Баккер и Бир пришли к выводу, что Цератозавр и мегалозавриды, должно быть, преимущественно охотились вблизи водоемов и внутри них, с Цератозавр также иногда поедает туши более крупных динозавров. Кроме того, исследователи отметили длинное, низкое и гибкое тело Цератозавр и мегалозавриды. По сравнению с другими тероподами Моррисона, Цератозавр показали более высокие нервные отростки на передних хвостовых позвонках, которые были вертикальными, а не наклонными к спине. Вместе с глубокими шевронными костями на нижней стороне хвоста они указывают на глубокий, «крокодиловий» хвост, возможно, приспособленный для плавания. Напротив, у аллозавридов более короткое, более высокое и жесткое тело с более длинными ногами. Они были бы приспособлены для быстрого бега по открытой местности и для охоты на крупных травоядный динозавры, такие как зауроподы и стегозавры, но, как предполагают Баккер и Бир, сезонно переключались на водную добычу, когда отсутствовали крупные травоядные.[44] Однако в 2019 году Юн поставил под сомнение эту теорию, предположив, что цератозавр был просто более способен охотиться на водную добычу, чем другие тероподы формации Моррисон, чем его современники, чем быть полностью полуводным.[48]

В своей популярной книге 1986 года Ереси динозавров Баккер утверждал, что кости верхней челюсти были только слабо прикреплены к окружающим костям черепа, что допускало некоторую степень движения внутри черепа, состояние, называемое черепной кинезис. Точно так же кости нижней челюсти могли бы двигаться друг против друга, а квадратная кость отклоняться наружу, раздвигая нижнюю челюсть в суставе челюсти. Взятые вместе, эти особенности позволили животному расширить свои челюсти, чтобы глотать большие пищевые продукты.[49] В исследовании 2008 года Кейси Холлидей и Лоуренс Уитмер повторно оценили аналогичные утверждения, сделанные в отношении других динозавров, и пришли к выводу, что наличие мышечного черепного кинезиса не может быть доказано ни для одного вида динозавров и, вероятно, отсутствовало у большинства из них.[50]
Функция носового рога и остеодермы
В 1884 году Марш считал носовой рог Цератозавр быть «самым мощным оружием» как для наступательных, так и для оборонительных целей, и Гилмор в 1920 году согласился с этой интерпретацией.[5]:331[2]:82 Однако в настоящее время использование рога в качестве оружия считается маловероятным.[7] В 1985 году Дэвид Норман считал, что рог был «вероятно, не для защиты от других хищников», но вместо этого мог использоваться для внутривидовой борьбы между самцами цератозавров, борющихся за право размножения.[51] Грегори С. Пол в 1988 г. предложил аналогичную функцию и проиллюстрировал два Цератозавр участвовал в состязании несмертельных ударов.[3] В 1990 году Роу и Готье пошли еще дальше, предположив, что носовой рог Цератозавр "вероятно, использовались только для демонстрации" и не играли никакой роли в физических столкновениях.[27] Если бы его использовали для демонстрации, рог, вероятно, был бы ярко окрашен.[9] Также была предложена функция отображения ряда остеодерм, идущих по средней линии тела.[27]
Функция передних конечностей

Сильно укороченные пястные кости и фаланги Цератозавр возникает вопрос, сохранила ли кисть руки захватывающую функцию, принятую у других базальных теропод. В пределах Ceratosauria еще более резкое сокращение кисти можно наблюдать у абелизаврид, у которых передняя конечность потерял свою первоначальную функцию,[52] И в Лимузавр. В статье 2016 года об анатомии Цератозавр manus, Каррано и Иона Шуаньер подчеркнули большое морфологическое сходство кисти с таковыми у других базальных теропод, предполагая, что она по-прежнему выполняла свою первоначальную захватывающую функцию, несмотря на ее укорочение. Хотя сохранились только первые фаланги, вторые фаланги были бы подвижными, на что указывало хорошо развитые суставные поверхности, и пальцы, вероятно, позволяли бы такую же степень движения, как и у других базальных теропод. Как и у других теропод, кроме абелизаврид, палец I был бы слегка повернут, когда согнутый.[11]
Мозг и чувства

Отливка полости мозга голотипа была сделана под наблюдением Марша, вероятно, во время подготовки черепа, что позволило Маршу сделать вывод, что мозг «был среднего размера, но сравнительно намного больше, чем у травоядных динозавров». Однако кости черепа впоследствии были скреплены вместе, поэтому точность этой модели не могла быть подтверждена более поздними исследованиями.[5][2]:93
Вторая, хорошо сохранившаяся черепная коробка была обнаружена с образцом MWC 1 во Фруите, Колорадо, и была обнаружена. КТ-сканирование палеонтологами Кентом Сандерсом и Дэвидом Смитом, что позволило реконструировать внутреннее ухо, крупные области мозга и черепные пазухи транспортировка крови от мозга. В 2005 году исследователи пришли к выводу, что Цератозавр обладал полостью мозга, типичной для базальных теропод и подобной полости мозга Аллозавр. Впечатления для обонятельные луковицы, обладающие обонянием, хорошо сохранились. Хотя они похожи на Аллозавр, они были меньше, чем в Тиранозавр, который, как считается, обладал очень острым обонянием. В полукружные каналы, которые отвечают за Чувство равновесия и, следовательно, позволяют делать выводы о привычной ориентации головы и передвижении, аналогичны тем, которые обнаруживаются у других теропод. У теропод эти структуры обычно консервативны, что позволяет предположить, что функциональные требования во время передвижения были одинаковыми для разных видов. Передний из полукружных каналов был расширен, что обычно наблюдается у двуногих животных. Ориентация бокового полукружного канала указывает на то, что голова и шея находились горизонтально в нейтральном положении.[53]
Слияние плюсневых костей и палеопатологии
Голотип C. nasicornis был обнаружен со слиянием его левых плюсневых костей II-IV.[54] Марш в 1884 году посвятил небольшую статью этой в то время неизвестной особенности динозавров, отметив близкое сходство с состоянием, наблюдаемым у современных птиц.[34] Наличие этой функции в Цератозавр стало спорным в 1890 году, когда Георг Баур предположил, что слияние голотипа было результатом исцеления перелом. Это утверждение повторил в 1892 году Коп, утверждая, что C. nasicornis следует отнести к разновидностям Мегалозавр из-за недостаточных анатомических различий между этими родами.[35] Однако с тех пор описаны примеры сросшихся плюсневых костей динозавров, которые не имеют патологического происхождения, включая таксоны более базальные, чем у динозавров. Цератозавр.[54] Осборн в 1920 году объяснил, что аномального роста костей не наблюдается и что сращение является необычным, но, вероятно, не патологическим.[2]:112 Рональд Раткевич в 1976 году утверждал, что это слияние ограничивало беговые способности животного, но это утверждение было отвергнуто Полом в 1988 году, который отметил, что та же особенность встречается у многих современных быстро передвигающихся животных, включая наземных птиц и копытных. .[3] Анализ 1999 года, проведенный Дарреном Танке и Брюсом Ротшильдом, показал, что слияние действительно было патологическим, подтверждая более раннее заявление Баура.[54] Другие сообщения о патологиях включают стрессовый перелом кости стопы, относящийся к этому роду,[55] а также сломанный зуб неустановленного вида Цератозавр что показывает признаки дальнейшего носить получил после перерыва.[54]
Палеосреда и палеобиогеография
Все североамериканские Цератозавр находки происходят из формации Моррисон, последовательности мелководных морских и аллювиальный осадочные породы в западной части Соединенных Штатов, и самый плодородный источник костей динозавров на континенте. В соответствии с радиометрическое датирование, возраст образования колеблется от 156,3 млн. годы старый (Mya) в основании,[56] и 146,8 миллиона лет на вершине,[57] что помещает его в конец Оксфордский, Кимериджский и ранний Титонский этапы позднеюрского периода. Цератозавр известен из кимериджских и титонских пластов свиты.[4]:49 Формация Моррисон интерпретируется как полузасушливый среда с четкими смачивать и сухие сезоны. Бассейн Моррисон простирался от Нью-Мексико до Альберты и Саскачевана и был сформирован, когда предшественники Передний диапазон Скалистых гор начали продвигаться на запад. Отложения с восточной стороны дренажные бассейны несли ручьи и реки и депонирован в болотистый низменности, озера, русла рек и поймы рек.[58] Эта формация похожа по возрасту на формацию Lourinhã в Португалии и формацию Tendaguru в Танзании.[59]

Формация Моррисон фиксирует среду и время, в которых доминировали гигантские динозавры-завроподы.[60] Другие динозавры, известные от Моррисона, включают тероподов. Копарион, Стокезозавр, Орнитолестес, Аллозавр, и Торвозавр; зауроподы Апатозавр, Брахиозавр, Камаразавр, и Диплодок; и орнитисхианы Камптозавр, Дриозавр, Отниелия, Горгульозавр, и Стегозавр.[61] Аллозавр, на долю которых приходилось от 70 до 75% всех особей теропод, находилась на вершине трофический уровень Моррисона пищевой сети.[43] Среди других позвоночных, которые разделяли эту палеосреду, были рыбы с луговыми плавниками, лягушки, саламандры, черепахи подобно Дорсеточелис, сфенодонты, ящерицы, наземные и водные крокодиломорфоны Такие как Hoplosuchus, и несколько видов птерозавры Такие как Harpactognathus и Мезадактиль. Снаряды из двустворчатые моллюски и водные улитки также распространены. Флора этого периода была обнаружена по окаменелостям зеленые водоросли, грибы, мхи, хвощи, саговники, гинкго, и несколько семейств хвойные породы. Растительность разнообразна от прибрежных лесов древовидные папоротники и папоротники (галерея леса ) папоротник саванны с редкими деревьями, такими как Араукария -подобное хвойное дерево Брахифиллум.[62]
Частичный Цератозавр Образец указывает на присутствие этого рода в португальском порто-ново, члене формации Лориньян. Многие из динозавров формации Лоуринья принадлежат к тем же родам, что и те, что встречаются в формации Моррисон, или имеют близких аналогов.[59] Помимо Цератозавр, исследователи также отметили наличие Аллозавр и Торвозавр в португальских породах известны в первую очередь от Моррисона, а Лоуринханозавр пока что поступили сообщения только из Португалии. Травоядные динозавры из члена Порто-Ново включают, среди прочего, зауроподов. Динхейрозавр и Zby, так же хорошо как стегозавр Miragaia.[63][29][30] Во время поздней юры Европа была отделена от Северной Америки еще узким Атлантический океан, и Португалия, как часть Пиренейский полуостров, все еще был отделен от других частей Европы. По мнению Матеуса и его коллег, сходство между португальской и североамериканской фаунами теропод указывает на наличие временного сухопутный мост, позволяющий обмен фауной.[29][30] Однако Малафая и его коллеги отстаивали более сложный сценарий, поскольку другие группы, такие как зауроподы, черепахи и крокодилы, демонстрируют явно различный видовой состав в Португалии и Северной Америке. Таким образом, зарождающееся разделение этих фаун могло привести к обмену некоторыми, но аллопатрическое видообразование в других группах.[31]
Рекомендации
- ^ а б c Марш, О. (1892). "Реставрация Клаозавр и Цератозавр". Американский журнал науки. 44 (262): 343–349. Bibcode:1892AmJS ... 44..343M. Дои:10.2475 / ajs.s3-44.262.343. HDL:2027 / hvd.32044107356040. S2CID 130216318.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты Гилмор, К. (1920). "Остеология плотоядных динозавров в Национальном музее США, с особым упором на роды Антродем (Аллозавр) и Цератозавр" (PDF). Бюллетень Национального музея США. 110 (110): 1–154. Дои:10.5479 / si.03629236.110.i. HDL:2027 / uiug.30112032536010.
- ^ а б c d е ж Пол, Грегори С. (1988). «Цератозавры». Хищные динозавры мира. Саймон и Шустер. стр.274–279. ISBN 978-0-671-61946-6.
- ^ а б c d е ж грамм час я Tykoski, R.S .; Роу, Т. (2004). «Цератозаврия». In Weishampel, D.B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия: второе издание. Калифорнийский университет Press. С. 47–70. ISBN 978-0-520-24209-8.
- ^ а б c d е Марш, О. (1884). «Основные персонажи американских динозавров юрского периода, часть VIII: Отряд тероподы» (PDF). Американский журнал науки. 27 (160): 329–340. Bibcode:1884AmJS ... 27..329M. Дои:10.2475 / ajs.s3-27.160.329. S2CID 131076004.
- ^ Therrien, F .; Хендерсон, Д. (2007). «Мой теропод больше вашего… или нет: размер тела определяется по длине черепа теропод». Журнал палеонтологии позвоночных. 27 (1): 108–115. Дои:10.1671 / 0272-4634 (2007) 27 [108: mtibty] 2.0.co; 2.
- ^ а б c d Глют, Д.Ф. (1997). "Цератозавр". Динозавры: Энциклопедия. McFarland & Company. С. 266–270. ISBN 978-0-89950-917-4.
- ^ а б c Пол, Грегори С. (2016). Принстонский полевой путеводитель по динозаврам (2-е изд.). Издательство Принстонского университета. п. 84. ISBN 978-0-691-16766-4.
- ^ а б c d Фостер, Дж. (2007). «От Гаргантюа до Меньшего: Зверинец Моррисона, Часть II». Юрский Запад: Динозавры формации Моррисон и их мир. Издательство Индианского университета. С. 162–242. ISBN 978-0-253-34870-8.
- ^ а б c d е ж грамм час я j k Carrano, M.T .; Сэмпсон, С. (2008). «Филогения Ceratosauria (Dinosauria: Theropoda)». Журнал систематической палеонтологии. 6 (2): 183–236. Дои:10.1017 / S1477201907002246. ISSN 1477-2019. S2CID 30068953.
- ^ а б Carrano, M.T .; Шойнер, Дж. (2016). "Новая информация о предплечье и кисти Ceratosaurus nasicornis Marsh, 1884 (Dinosauria, Theropoda) с последствиями для эволюции передних конечностей теропод ". Журнал палеонтологии позвоночных. 36 (2): –1054497. Дои:10.1080/02724634.2015.1054497. S2CID 88089084.
- ^ а б c d е ж грамм час я j k л м п о Madsen, J.H .; Уэллс, С.П. (2000). Цератозавр (Dinosauria, Theropoda): новая остеология. Геологическая служба Юты. С. 1–80. ISBN 978-1-55791-380-7.
- ^ а б c d Хендерсон, Д. (1998). «Морфология черепа и зубов как индикаторы разделения ниши у симпатрических тероподов Моррисонской формации». Гайя (15): 219–226.
- ^ а б c Делькур, Рафаэль (2018). «Палеобиология цератозавров: новые взгляды на эволюцию и экологию южных правителей». Научные отчеты. 8 (1): 9730. Bibcode:2018НатСР ... 8.9730D. Дои:10.1038 / с41598-018-28154-х. ЧВК 6021374. PMID 29950661.
- ^ а б Britt, B.B .; Miles, CA; Cloward, K.C .; Мэдсен, Дж. (1999). "Несовершеннолетний Цератозавр (Theropoda, Dinosauria) из карьера Bone Cabin West (верхняя юра, формация Моррисон), Вайоминг ". Журнал палеонтологии позвоночных. 19 (Приложение к № 3): 33А. Дои:10.1080/02724634.1999.10011202.
- ^ а б c Раухут, О.В.М. (2004). "Происхождение и анатомия Genyodectes serus, зубастый цератозавр (Dinosauria: Theropoda) из Патагонии ". Журнал палеонтологии позвоночных. 24 (4): 894–902. Дои:10.1671 / 0272-4634 (2004) 024 [0894: paaogs] 2.0.co; 2.
- ^ Бринкман, П. (2010). Вторая атака динозавров юрского периода. Музеи и палеонтология в Америке на рубеже ХХ века. Издательство Чикагского университета. п. 10. ISBN 978-0-226-07472-6.
- ^ Симпсон, Д. (1979) [1854]. Латинский словарь Кассела (5-е изд.). Лондон: Cassell Ltd., стр. 153, 387. ISBN 978-0-304-52257-6.
- ^ Жаклин Т. (3 мая 2012 г.). «Дэвид Кох пожертвовал 35 миллионов долларов Национальному музею естественной истории для строительства зала динозавров». Вашингтон Пост. Получено 4 декабря, 2017.
- ^ а б "Хорошее, плохое и уродливое: демонтаж исторических окаменелостей, часть вторая". Раскопки летописи окаменелостей: палеобиология в Смитсоновском институте. Смитсоновский национальный музей естественной истории. Архивировано из оригинал 10 октября 2017 г.. Получено 4 декабря, 2017.
- ^ Тидвелл, В.(1996). «Восстановление раздробленных черепов динозавров юрского периода для демонстрации». Континентальная юра. Бюллетень Музея Северной Аризоны. 60.
- ^ Britt, B.B .; Chure, D.J .; Holtz, T.R., Jr .; Miles, CA; Штадтман, К. (2000). "Повторный анализ филогенетического сродства Цератозавр (Theropoda, Dinosauria) на основе новых образцов из Юты, Колорадо и Вайоминга ". Журнал палеонтологии позвоночных. 20 (доп.): 32А. Дои:10.1080/02724634.2000.10010765. S2CID 220412294.
- ^ Раухут, О.В.М. (2003). «Взаимоотношения и эволюция базальных динозавров-теропод». Специальные статьи по палеонтологии: 25.
- ^ Фостер, Дж. (2007). «Приложение». Юрский Запад: Динозавры формации Моррисон и их мир. Издательство Индианского университета. С. 327–329. ISBN 978-0-253-34870-8.
- ^ а б Zils, C.W .; Moritz, A .; Саанане, К. (1995). «Тендагуру, самое известное место обитания динозавров в Африке. Обзор, обзор и перспективы на будущее». Documenta Naturae. 97: 1–41.
- ^ а б c d Яненш, В. (1925). "Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas". Palaeontographica (на немецком). Дополнение VIII: 1–100.
- ^ а б c d Rowe, T .; Готье, Дж. (1990). «Цератозаврия». In Weishampel, D.B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия. Калифорнийский университет Press. С. 151–168. ISBN 978-0-520-06726-4.
- ^ а б c Раухут, О.В.М. (2011). «Тероподные динозавры из поздней юры Тендагуру (Танзания)». Специальные статьи по палеонтологии. 86: 195–239.
- ^ а б c Mateus, O .; Антунес, М. (2000). "Цератозавр (Dinosauria: Theropoda) в поздней юре Португалии ». 31-й Международный геологический конгресс, тезисы. Рио де Жанейро.
- ^ а б c d Mateus, O .; Вален, А .; Антунес, М. (2006). "Крупная фауна теропод формации Лориньян (Португалия) и ее сходство с формацией Моррисон, с описанием нового вида Аллозавр". In Foster, J.R .; Lucas, S.G. (ред.). Палеонтология и геология верхнеюрской свиты Моррисон. Музей естественной истории и науки Нью-Мексико, Бюллетень 36.
- ^ а б c Malafaia, E .; Ортега, Ф .; Escaso, F .; Сильва, Б. (3 октября 2015 г.). "Новое свидетельство Цератозавр (Dinosauria: Theropoda) из поздней юры Лузитанской котловины, Португалия ". Историческая биология. 27 (7): 938–946. Дои:10.1080/08912963.2014.915820. ISSN 0891-2963. S2CID 129349509.
- ^ Сото, М .; Переа, Д. (2008). «Цератозаврид (Dinosauria, Theropoda) из поздней юры – раннего мела Уругвая». Журнал палеонтологии позвоночных. 28 (2): 439–444. Дои:10.1671 / 0272-4634 (2008) 28 [439: acdtft] 2.0.co; 2.
- ^ Сото, Матиас; Ториньо, Пабло; Переа, Даниил (1 ноября 2020 г.). "Цератозавр (Theropoda, Ceratosauria) зубы из формации Такуарембо (поздняя юра, Уругвай) ». Журнал южноамериканских наук о Земле. 103: 102781. Дои:10.1016 / j.jsames.2020.102781. ISSN 0895-9811.
- ^ а б Марш, О. (1884). "На соединенных плюсневых костях Цератозавр". Американский журнал науки. 28 (164): 161–162. Bibcode:1884AmJS ... 28..161M. Дои:10.2475 / ajs.s3-28.164.161. S2CID 131737380.
- ^ а б Коуп, Э. (1892). "На черепе динозавра Lælaps incrassatus Справиться". Труды Американского философского общества. 30 (138): 240–245. ISSN 0003-049X. JSTOR 983173.
- ^ Hendrickx, C .; Hartman, S.A .; Матеус, О. (2015). «Обзор открытий и классификации нептичьих теропод». Журнал PalArch по палеонтологии позвоночных. 12 (1): 1–73.
- ^ Сюй, X .; Clark, J.M .; Mo, J .; Choiniere, J .; Forster, C.A .; Эриксон, G.M .; Hone, D.W .; Салливан, К .; Eberth, D.A .; Nesbitt, S .; Чжао, Q. (2009). «Юрский цератозавр из Китая помогает прояснить цифровую гомологию птиц» (PDF). Природа. 459 (7249): 940–944. Bibcode:2009Натура.459..940X. Дои:10.1038 / природа08124. PMID 19536256. S2CID 4358448.
- ^ а б Pol, D .; Раухут, О.В.М. (2012). «Среднеюрский абелизаврид из Патагонии и раннее разнообразие динозавров-теропод». Труды Королевского общества B: биологические науки. 279 (1804): 3170–5. Дои:10.1098 / rspb.2012.0660. ЧВК 3385738. PMID 22628475.
- ^ Делькур, Р. (2017). "Пересмотренная морфология Pycnonemosaurus nevesi Kellner & Campos, 2002 (Theropoda: Abelisauridae) и его филогенетические отношения ". Zootaxa. 4276 (1): 1–45. Дои:10.11646 / zootaxa.4276.1.1. PMID 28610214.
- ^ а б Wang, S .; Stiegler, J .; Amiot, R .; Ван, X .; Du, G.-H .; Clark, J.M .; Сюй, X. (2017). «Экстремальные онтогенетические изменения у теропод цератозавров» (PDF). Текущая биология. 27 (1): 144–148. Дои:10.1016 / j.cub.2016.10.043. PMID 28017609. S2CID 441498.
- ^ Huene, F.v. (1926). «О нескольких известных и неизвестных рептилиях отряда Saurischia из Англии и Франции». Летопись и журнал естественной истории. Серия 9 (17): 473–489. Дои:10.1080/00222932608633437.
- ^ а б Rauhut, O.W.M .; Milner, A.C .; Мур-Фэй, С. (2010). «Черепная остеология и филогенетическое положение тероподового динозавра. Proceratosaurus bradleyi (Woodward, 1910) из средней юры Англии ». Зоологический журнал Линнеевского общества. 158 (1): 155–195. Дои:10.1111 / j.1096-3642.2009.00591.x.
- ^ а б Фостер, Дж. Р. (2003). Палеоэкологический анализ фауны позвоночных формации Моррисон (верхняя юра), регион Скалистых гор, США. Бюллетень Музея естественной истории и науки Нью-Мексико. 23. Альбукерке, Нью-Мексико: Музей естественной истории и науки Нью-Мексико. п. 29.
- ^ а б Баккер, Роберт Т .; Бир, Гэри (2004). «Расследование на месте преступления динозавров: поведение теропод в Комо-Блафф, Вайоминг, и эволюция птицеводства». В Currie, P.J .; Koppelhus, E.B .; Шугар, M.A .; Райт, Дж. Л. (ред.). Пернатые драконы: исследования перехода от динозавров к птицам. Блумингтон и Индианаполис: Издательство Индианского университета. С. 301–342. ISBN 978-0-253-34373-4.
- ^ а б Foster, J.R .; Чуре, Д.Дж. (2006). "Аллометрия задних конечностей у позднеюрских тероподных динозавров. Аллозавр, с комментариями о его изобилии и распространении ». Бюллетень Музея естественной истории и науки Нью-Мексико. 36: 119–122.
- ^ Карпентер, Кеннет (2010). "Изменчивость в популяции теропод (динозавров): Аллозавр из карьера Кливленд-Ллойд (верхняя юра), штат Юта, США ». Палеонтологические исследования. 14 (4): 250–259. Дои:10.2517/1342-8144-14.4.250. S2CID 84635714.
- ^ Карпентер, Кеннет; Пол, Грегори С. (2015). "Комментировать Аллозавр Marsh, 1877 (Dinosauria, Theropoda): предложено сохранение использования путем обозначения неотипа для своего типового вида. Аллозавр ломкий Марш, 1877 г. ». Вестник зоологической номенклатуры. 72: 1.
- ^ Чаню Юнь (2019). »Комментарии к экологии юрского динозавра теропод. Цератозавр (Dinosauria: Theropoda) с критической переоценкой предполагаемого полуводного образа жизни ». Volumina Jurassica. в прессе. Архивировано из оригинал 20 июля 2019 г.. Получено 19 сентября, 2019.
- ^ Баккер, Р. (1986). Ереси динозавров. ПАЛАИ. 2. Уильям Морроу и компания. п. 523. Bibcode:1987 Палай ... 2..523G. Дои:10.2307/3514623. ISBN 978-0-688-04287-5. JSTOR 3514623.
- ^ Холлидей, C.M .; Витмер, Л.М. (12 декабря 2008 г.). «Краниальный кинезис у динозавров: внутричерепные суставы, транспортирные мышцы и их значение для развития черепа и функции в диапсидах». Журнал палеонтологии позвоночных. 28 (4): 1073–1088. Дои:10.1671/0272-4634-28.4.1073. S2CID 15142387.
- ^ Норман, Д. (1985). «Карнозавры». Иллюстрированная энциклопедия динозавров. Саламандра Букс Лтд., Стр. 62–67. ISBN 978-0-517-46890-6.
- ^ Сентер, П. (2010). «Рудиментарные скелетные структуры динозавров». Журнал зоологии. 280 (4): 60–71. Дои:10.1111 / j.1469-7998.2009.00640.x.
- ^ Сандерс, Р.К .; Смит, Д.К. (2005). "Эндокраниум динозавра теропод Цератозавр учился компьютерной томографии ». Acta Palaeontologica Polonica. 50 (3).
- ^ а б c d Мольнар, Р. (2001). «Палеопатология теропод: обзор литературы». In Tanke, D.H .; Карпентер, К. (ред.). Мезозойская жизнь позвоночных. Издательство Индианского университета. С. 337–363.
- ^ Ротшильд, В .; Tanke, D.H .; Форд, Т. (2001). «Стрессовые переломы теропод и отрывы сухожилий как ключ к активности». In Tanke, D.H .; Карпентер, К. (ред.). Мезозойская жизнь позвоночных. Издательство Индианского университета. С. 331–336.
- ^ Трухильо, К.С.; Чемберлен, К.Р .; Стрикленд, А. (2006). «Оксфордский возраст U / Pb из анализа SHRIMP для верхнеюрской формации Моррисон на юго-востоке Вайоминга с последствиями для биостратиграфических корреляций». Рефераты Геологического общества Америки с программами. 38 (6): 7.
- ^ Билби, С.А. (1998). "Кливленд-Ллойд Карьер динозавров - возраст, стратиграфия и условия осадконакопления". В Carpenter, K .; Chure, D .; Киркланд, Дж. (ред.). Формация Моррисон: междисциплинарное исследование. Современная геология 22. Группа Тейлор и Фрэнсис. С. 87–120. ISSN 0026-7775.
- ^ Рассел, Д.А. (1989). Одиссея во времени: динозавры Северной Америки. Миноква, Висконсин: NorthWord Press. С. 64–70. ISBN 978-1-55971-038-1.
- ^ а б Матеус, О. (2006). «Динозавры юрского периода из формации Моррисон (США), формации Лоуринья и Алкобаса (Португалия) и пластов Тендагуру (Танзания): сравнение». In Foster, J.R .; Лукас, С.Г. (ред.). Палеонтология и геология верхнеюрской свиты Моррисон. Бюллетень Музея естественной истории и науки Нью-Мексико. 36. Альбукерке, Нью-Мексико: Музей естественной истории и науки Нью-Мексико. С. 223–231.
- ^ Фостер, Дж. (2007). «Приложение». Юрский Запад: Динозавры формации Моррисон и их мир. Издательство Индианского университета. С. 327–329.
- ^ Chure, D.J .; Litwin, R .; Hasiotis, S.T .; Evanoff, E .; Карпентер, К. (2006). «Фауна и флора формации Моррисон: 2006». In Foster, J.R .; Лукас, С.Г. (ред.). Палеонтология и геология верхнеюрской свиты Моррисон. Бюллетень Музея естественной истории и науки Нью-Мексико. 36. Альбукерке, Нью-Мексико: Музей естественной истории и науки Нью-Мексико. С. 233–248.
- ^ Карпентер, К. (2006). "Крупнейшее из большого: критическая переоценка мега-зауроподов Amphicoelias fragillimus". In Foster, J.R .; Lucas, S.G. (ред.). Палеонтология и геология верхнеюрской свиты Моррисон. Бюллетень Музея естественной истории и науки Нью-Мексико. 36. Альбукерке, Нью-Мексико: Музей естественной истории и науки Нью-Мексико. С. 131–138.
- ^ Mateus, O .; Mannion, P.D .; Апчерч, П. (2014). "Зби атлантик, новый туриазаврический зауропод (Dinosauria, Eusauropoda) из поздней юры Португалии ». Журнал палеонтологии позвоночных. 34 (3): 618–634. Дои:10.1080/02724634.2013.822875. S2CID 59387149.
внешняя ссылка
 СМИ, связанные с Цератозавр в Wikimedia Commons
СМИ, связанные с Цератозавр в Wikimedia Commons
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||













