Карнотавр - Carnotaurus
| Карнотавр | |
|---|---|
 | |
| Конный слепок в музее Хлупач в Прага | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Семья: | †Abelisauridae |
| Clade: | †Фурилеузаурия |
| Племя: | †Карнотаурини |
| Род: | †Карнотавр Бонапарт, 1985 |
| Разновидность: | †C. sastrei |
| Биномиальное имя | |
| †Carnotaurus sastrei Бонапарт, 1985 | |
Карнотавр /ˌkɑːrпoʊˈтɔːrəs/ это род из теропод динозавр который жил в Южная Америка вовремя Поздний мел период между 84 и 72 миллионами лет назад. Единственный разновидность является Carnotaurus sastrei. Известный по единственному хорошо сохранившемуся скелету, это один из наиболее изученных тероподов Южное полушарие. Скелет, найденный в 1984 г., был обнаружен в Провинция Чубут из Аргентина из скал Формация Ла Колония. Карнотавр является производным членом Abelisauridae, группа крупных теропод, населявшая большую хищный ниша на южных территориях Гондвана в конце Меловой. Внутри Abelisauridae этот род часто считается членом Brachyrostra, клады короткорылых форм, обитающих в Южной Америке.
Карнотавр был легко сложен, двуногий хищник размером от 7,5 до 9 м (от 24,6 до 29,5 футов) в длину и весом не менее 1,35 метрических тонн (1,33 длинных тонны; 1,49 коротких тонны). Как теропод, Карнотавр был узкоспециализированным и самобытным. Он был толстым рога над глазами, черта, невидимая ни в одном другом плотоядный динозавров и очень глубокий череп, сидящий на мускулистой шее. Карнотавр далее характеризовался небольшим, рудиментарный передние конечности и длинные тонкие задние конечности. Скелет сохранился с обширными отпечатками кожи, на которых видна мозаика небольших неперекрывающихся напольные весы примерно 5 мм в диаметре. Мозаика прервана большими шишками по бокам животного, на них нет намека на перья.
Отличительные рога и мускулистая шея, возможно, использовались в боевых действиях. сородичи. Согласно отдельным исследованиям, соперничающие люди могли сражаться друг с другом быстрыми ударами по голове, медленными толчками верхней стороной черепа или тараном друг друга, используя свои рога в качестве амортизаторов. Привычки кормления Карнотавр остаются неясными: некоторые исследования показали, что животное могло охотиться на очень крупную добычу, такую как зауроподы, в то время как другие исследования показали, что он охотился в основном на относительно мелких животных. Полость мозга свидетельствует об остром обонянии, в то время как слух и зрение были менее развиты. Карнотавр вероятно, был хорошо приспособлен для бега и, возможно, был одним из самых быстрых крупных теропод.
Открытие
Единственный скелет (голотип MACN-CH 894) был обнаружен в 1984 году экспедицией под руководством аргентинского палеонтолога. Хосе Бонапарт.[A] Эта экспедиция также вернула своеобразный колючий зауропод Амаргазавр.[3] Это была восьмая экспедиция в рамках проекта «Наземные позвоночные юрского и мелового периодов Южной Америки», который стартовал в 1976 году и спонсировался Национальное географическое общество.[3][B] Скелет хорошо сохранился и сочлененный (все еще соединены вместе), причем только две трети задней части хвоста, большая часть голени и задние лапы разрушены выветривание.[C][5] Скелет принадлежал взрослой особи, на что указывает слитный швы в черепная коробка.[6] Он был найден лежащим на правом боку, показывая типичный поза смерти с отогнутой назад шеей над туловищем.[7] Необычно сохранились обширные кожные вмятины.[D] Ввиду важности этих отпечатков была начата вторая экспедиция для повторного исследования исходного места раскопок, что привело к обнаружению нескольких дополнительных участков кожи.[7] Череп был деформирован во время окаменения, при этом кости морды левой стороны смещены вперед относительно правой стороны, носовые кости сдвинуты вверх, а предчелюстные кости толкнул назад на носовые кости. Деформация также преувеличивала искривление верхней челюсти вверх.[E] Морда была подвержена деформации сильнее, чем задняя часть черепа, возможно, из-за большей жесткости последней. На виде сверху или снизу верхние челюсти имели менее U-образную форму, чем нижние, что приводило к очевидному несоответствию. Это несоответствие является результатом деформации, действующей с боков, которая затронула верхние челюсти, но не нижние челюсти, возможно, из-за большей гибкости суставов внутри последних.[1]

Скелет был собран на ферме "Почо Састре" недалеко от Бахада Морено в Отделение Тельсен из Провинция Чубут, Аргентина.[5] Потому что он был встроен в большой гематит конкреция, очень твердый камень, подготовка к нему была сложной и продвигалась медленно.[9][5] В 1985 году Бонапарт опубликовал заметку, в которой Carnotaurus sastrei как новый род и вид и кратко описывая череп и нижнюю челюсть.[5] Общее название Карнотавр происходит от латинского carno [carnis] («плоть») и taurus («бык») и может быть переведено как «мясоедный бык», что является намеком на бычьи рога животного.[10] В конкретное имя састрей чтит Анхеля Састре, владельца ранчо, где был найден скелет.[11] Всестороннее описание всего скелета последовало в 1990 году.[4] После Абелизавр, Карнотавр был вторым обнаруженным членом семейства Abelisauridae.[12] В течение многих лет он был самым понятым членом своего семейства, а также самым понимаемым тероподом из Южное полушарие.[13][14] Так продолжалось до 21 века, когда подобные хорошо сохранившиеся абелизавриды были описаны, в том числе Ауказавр, Маджунгазавр и Скорпиовенатор, что позволяет ученым по-новому оценить некоторые аспекты анатомии Карнотавр.[F] Скелет голотипа отображается в Аргентинский музей естественных наук, Бернардино Ривадавиа;[ГРАММ] точные копии можно увидеть в этом и других музеях мира.[15] Скульпторы Стивен и Сильвия Черкас изготовили скульптуру в натуральную величину. Карнотавр который ранее был выставлен на Музей естественной истории округа Лос-Анджелес. Эта скульптура, заказанная музеем в середине 1980-х годов, вероятно, является первой реставрацией теропода с точной кожей.[7][16]
Описание

Карнотавр был крупным, но легко сложенным хищником.[17] Единственная известная особь была около 7,5–9 м (24,6–29,5 футов) в длину,[ЧАС][Я][19] изготовление Карнотавр один из крупнейших абелизаврид.[J][K][19] Экриксинатозавр и возможно Абелизавр, которые очень неполные, могли быть похожими или большими по размеру.[L][M][N] Исследование 2016 года показало, что только Пикнонемозавр на высоте 8,9 м (29,2 фута) был длиннее, чем Карнотавр; это было оценено в 7,8 м (25,6 футов).[21] Его масса оценивается в 1350 кг (1,33 длинных тонны; 1,49 коротких тонн).[O] 1500 кг (1,5 длинных тонны; 1,7 коротких тонны),[П] 2000 кг (2,0 длинных тонны; 2,2 коротких тонны),[19] 2100 кг (2,1 длинных тонны; 2,3 коротких тонны)[Q], и 1,306–1,743 кг (1,285–1,715 длинных тонн; 1,440–1,921 коротких тонн)[24] в отдельных исследованиях, в которых использовались разные методы оценки. Карнотавр был узкоспециализированным теропод, что особенно заметно по характеристикам череп, то позвонки и передние конечности.[Р] В таз а задние конечности, с другой стороны, оставались относительно консервативными, напоминая таковые более базальный Цератозавр. И таз, и задние конечности были длинными и тонкими. Слева бедренная кость (бедренная кость) человека имеет длину 103 см, но средний диаметр составляет всего 11 см.[S]
Череп

Череп размером 59,6 см (23,5 дюйма) в длину был пропорционально короче и глубже, чем у любого другого крупного плотоядного динозавра.[T][U] В морда был умеренно широким, не таким сужающимся, как у более базальных теропод, таких как Цератозавр, а челюсти были загнуты вверх.[25] Над глазами наискось торчала пара рогов. Эти рога, образованные лобной кости[V] были толстыми и конусообразными, внутренне твердыми, несколько уплощенными по вертикали в поперечном сечении и имели длину 15 см (5,9 дюйма).[6][1] Бонапарт в 1990 году предположил, что эти рога, вероятно, образовывали костные ядра гораздо более длинных ороговевших оболочек.[W] Маурисио Черрони и его коллеги в 2020 году согласились с тем, что рога поддерживают ороговевшие оболочки, но утверждали, что эти оболочки не были бы намного длиннее костных стержней.[1]
Как и у других динозавров, череп был пробит шестью основные отверстия черепа с каждой стороны. Переднее из этих отверстий внешний нарис (костная ноздря), была субпрямоугольной и была направлена в стороны и вперед, но не была наклонной при виде сбоку, как у некоторых других цератозавров, таких как Цератозавр. Это отверстие было образовано только носовой и предчелюстной частью, тогда как у некоторых родственных цератозавров верхняя челюсть также способствовала этому отверстию. Между костной ноздрей и орбита (открыв глаза) был анторбитальное окно. В Карнотавр, это открытие было больше, чем длинное, в то время как оно было больше, чем у связанных форм, таких как Скорпиовенатор и Маджунгазавр. Анторбитальное отверстие ограничивалось более крупным углублением, анторбитальная ямка, который был образован утопленными частями верхней челюсти спереди и слезный позади. Как и у всех абелизаврид, это углубление было небольшим в Карнотавр. В нижнем переднем углу анторбитальной ямки было отверстие меньшего размера, Промаксиллярное отверстие, который вел в заполненную воздухом полость внутри верхней челюсти.[1] Глаз располагался в верхней части глазницы в форме замочной скважины.[ИКС] Эта верхняя часть была пропорционально маленькой и субциркульной и отделялась от нижней части орбиты выступающим вперед. заглазничная кость.[1] Он был слегка повернут вперед, что, вероятно, позволяло бинокулярное зрение.[Y] Форма глазницы, похожая на замочную скважину, вероятно, была связана с заметным укорочением черепа, а также встречается у родственных коротконорых абелизаврид.[1] Как и у всех абелизавридов, лобная кость (на крыше черепа между глазами) исключен из орбиты. За орбитой было два отверстия, подвисочное отверстие на стороне и надвисочное окно на вершине черепа. Подвисочное отверстие было высоким, коротким и почковидным, а надвисочное отверстие коротким и квадратным. Еще одно открытие, нижнечелюстное отверстие, располагалась в нижней челюсти - в Карнотавр, это отверстие было сравнительно большим.[1]

На каждой стороне верхней челюсти было по четыре предчелюстной и двенадцать верхнечелюстной зубы,[Z] в то время как нижняя челюсть были оснащены пятнадцатью зубной зубов с каждой стороны.[AA][1] Зубы были описаны как длинные и тонкие,[9] в отличие от очень коротких зубов, наблюдаемых у других абелизаврид.[25] Однако Черрони и его коллеги в своем описании черепа 2020 года заявили, что все извергнутый зубы были серьезно повреждены во время раскопок и позже были реконструированы с помощью гипса (Бонапарт в 1990 году только отметил, что некоторые зубы нижней челюсти были фрагментированы).[1][AB] Таким образом, надежная информация о форме зубов ограничивается замещающими зубами и корнями зубов, которые все еще закрыты челюстью, и ее можно изучить с помощью компьютерной томографии.[1] Зубы для замены были низкие, уплощенные. короны, были близко расположены и наклонены вперед примерно на 45 °.[1] В своем описании 1990 года Бонапарт отметил, что нижняя челюсть неглубокая и слабо построенная, с зубной (передняя кость челюсти) соединена с задними костями челюсти всего двумя точками контакта; это контрастирует с массивным черепом.[9][AC] Серрони и его коллеги вместо этого обнаружили множественные, но неплотные связи между зубной частью и костями задней челюсти. Таким образом, эта артикуляция была очень гибкой, но не обязательно слабой.[1] Нижний край зубной кости выпуклый, в то время как прямой Маджунгазавр.[1]

На нижней челюсти обнаружена окостеневший подъязычные кости, в том положении, в котором они были бы, если бы животное было живым. Эти тонкие кости, поддерживающие мускулатуру языка и несколько других мышц, редко встречаются у динозавров, потому что они часто хрящевой и не соединены с другими костями и поэтому легко теряются.[ОБЪЯВЛЕНИЕ][26][1] В Карнотаврсохранились три подъязычные кости: пара изогнутых стержневидных цератобранхиальных костей, которые сочленяются с одним трапециевидным элементом - базихиальной. Карнотавр - единственный известный нептичий теропод, от которого известен базихиал.[1] В задней части черепа, как и у других абелизаврид, были хорошо развитые, заполненные воздухом камеры, окружавшие черепную коробку. Присутствовали две отдельные камерные системы, паратимпанальная система, которая была подключена к среднее ухо полости, а также камеры, образовавшиеся из наростов воздушные мешочки шеи.[24]
Номер аутапоморфии (отличительные черты) можно найти в черепе, включая пару рогов и очень короткий и глубокий череп. На верхней челюсти были выемки над переднечелюстным отверстием, которые должны были быть вырыты анторбитальным воздушным синусом (воздушные проходы в рыле). Носослезный канал, по которому транспортируется глазная жидкость, выходит на медиальную (внутреннюю) поверхность слезной жидкости через канал неопределенной функции. Другие предложенные аутапоморфии включают глубокую и длинную, заполненную воздухом выемку в квадратный и удлиненное углубление на крыловидный из нёбо.[1]
Позвонки

Позвоночный столб состоял из десяти шейный (шея), двенадцать спинной, шесть слитных крестцовый[AE] и неизвестное количество каудальный (хвостовые) позвонки.[4] Шея была почти прямой, а не S-образной, как у других теропод, а также необычно широкой, особенно в направлении основания.[27] В верхней части позвоночного столба шеи имелся двойной ряд увеличенных, направленных вверх костных отростков, называемых эпипофизы, создавая гладкую впадину на вершине шейных позвонков. Эти отростки были самыми высокими точками позвоночника, возвышаясь над необычно низким остистые отростки.[4][26] Эпипофизы, вероятно, обеспечивали места прикрепления для заметно сильной мускулатуры шеи.[AF] Подобный двойной ряд присутствовал и в хвосте, образованном там сильно модифицированным хвостовые ребра, на виде спереди выступают вверх в форме буквы V, их внутренние стороны создают гладкую плоскую верхнюю поверхность передних хвостовых позвонков. Конец каждого хвостового ребра имел выступающее вперед крючковидное расширение, которое соединялось с каудальным ребром предыдущего позвонка.[26][28]
Передние конечности

Передние конечности были пропорционально короче, чем у любых других крупных хищных динозавров, включая Тиранозавр.[AG] Предплечье было всего в четверть от плеча. Не было карпалия в руке, так что пястные кости шарнирно соединяется непосредственно с предплечьем.[29] Рука показала четыре основные цифры,[4] хотя, по-видимому, только две средние из них заканчивались костями пальцев, в то время как четвертая состояла из одной похожей на шину пястной кости, которая, возможно, представляла собой внешнюю «шпору». Сами пальцы были сросшимися и неподвижными, и, возможно, без когтей.[30] Карнотавр отличается от всех других абелизаврид наличием пропорционально более коротких и крепких передних конечностей, а также наличием четвертой, похожей на шину пястной кости как самой длинной кости в руке.[29] Исследование 2009 года предполагает, что руки были рудиментарный у абелизаврид, потому что нервные волокна, ответственные за передачу стимулов, были сокращены до такой степени, как сегодня эму и киви, у которых также есть рудиментарные передние конечности.[31]
Кожа

Карнотавр был первым обнаруженным динозавром-теропод со значительным количеством ископаемое кожные впечатления.[7] Эти отпечатки, обнаруженные под правой стороной скелета, поступают с разных частей тела, включая нижнюю челюсть,[7] передняя часть шеи, плечевой пояс, а грудная клетка.[AH] Самый большой участок кожи соответствует передней части хвоста.[AI] Первоначально правая сторона черепа также была покрыта большими участками кожи - это не было обнаружено при подготовке черепа, и эти участки были случайно уничтожены.[7] Тем не менее текстура поверхности нескольких костей черепа позволяет сделать вывод об их вероятном покрытии. Бугристая поверхность с бороздками, ямками и небольшими отверстиями по бокам и спереди морды указывает на чешуйчатый покров, возможно, с плоскими чешуйками, как у современных крокодилов. Верх морды был изрезан множеством мелких отверстий и шипов - эту текстуру, вероятно, можно соотнести с ороговевшей подушечкой (роговым покровом). Такая накладка также встречалась в Маджунгазавр но отсутствовал в Абелизавр и Rugops. Ряд крупных чешуек, вероятно, окружал глаз, о чем свидетельствует бугристая поверхность с продольными бороздками на слезной и заглазничной костях.[1][AJ]
Кожа состояла из мозаики из многоугольных неперекрывающихся чешуек диаметром примерно 5 мм (0,20 дюйма). Эта мозаика была разделена тонкими параллельными бороздками.[AK] Чешуйки были одинаковыми для разных частей тела, за исключением головы, которая, по-видимому, демонстрировала различный, неправильный рисунок чешуек.[AL][15] Нет никаких доказательств наличия перьев.[7] Уникально для теропод было остеодермы (шишковидные кожные кости), идущие по бокам шеи, спины и хвоста неправильными рядами. Каждая неровность имела низкий гребень и имела диаметр от 4 до 5 см (от 1,6 до 2,0 дюймов). Они располагались на расстоянии от 8 до 10 см (от 3,1 до 3,9 дюйма) друг от друга и становились больше по направлению к верхушке животного. Неровности, вероятно, представляют собой скопления конденсированных щитки, похожий на те, что видны на мягкой оборке, идущей вдоль средней линии тела в гадрозаврид («утконосые») динозавры.[ЯВЛЯЮСЬ][7] Стивен Черкас (1997) предположил, что эти структуры могли защищать бока животного во время борьбы с представителями того же вида (сородичи ) и других теропод, утверждая, что подобные структуры можно найти на шее современных игуана где они обеспечивают ограниченную защиту в бою.[7]
Классификация


Карнотавр один из наиболее понятных роды из Abelisauridae, семейство больших теропод, ограниченное древними южными суперконтинент Гондвана. Абелизавриды были доминирующими хищниками в позднем меловом периоде Гондваны, заменив кархародонтозавриды и заняв экологическую нишу, заполненную тираннозавриды на северных континентах.[17] Несколько примечательных черт, которые развились в этом семействе, включая укорочение черепа и рук, а также особенности шейных и хвостовых позвонков, были более выражены в Карнотавр чем в любом другом абелизавриде.[AN][AO][28]
Хотя отношения внутри Abelisauridae обсуждаются, Карнотавр неизменно считается одним из самых полученный членов семьи кладистическим анализом.[AP] Его ближайший родственник мог быть Ауказавр[32][33][34][35] или же Маджунгазавр.[36][37][38] Обзор 2008 года, напротив, предположил, что Карнотавр не был тесно связан ни с одним из родов, и вместо этого предложил Илокелесия как его сестринский таксон.[AQ] Хуан Канале и его коллеги в 2009 году возвели новую кладу Брахиростры, в которую вошли Карнотавр но нет Маджунгазавр; С тех пор этой классификации последовал ряд исследований.[32][35][39]
Карнотавр является одноименным для двух подгрупп Abelisauridae: Carnotaurinae и Карнотаурини. Палеонтологи не всегда принимают эти группы. Carnotaurinae был определен как включающий всех производных абелизаврид, за исключением Абелизавр, который в большинстве исследований считается базальным членом.[40] Однако обзор 2008 года показал, что Абелизавр вместо этого был производным абелизавридом.[AR] Карнотаурини было предложено назвать клады образована Карнотавр и Ауказавр;[33] только те палеонтологи, которые считают Ауказавр как ближайший родственник Карнотавр используйте эту группу.[41]
Ниже представлена кладограмма, опубликованная Канале и его коллегами в 2009 году.[32]
| Carnotaurinae |
| ||||||||||||||||||||||||||||||
Палеобиология
Функция рогов

Карнотавр - единственное известное хищное двуногое животное с парой рогов на лобной кости.[42] Использование этих рогов не совсем понятно. Несколько интерпретаций вращались вокруг использования в борьбе с сородичами или в убийстве добычи, хотя также возможно использование в демонстрации для ухаживания или признания представителей одного и того же вида.[1]
Грег Пол (1988) предположил, что рога были ударом оружия и что небольшая орбита сводила к минимуму возможность поранить глаза во время боя.[9] Херардо Маццетта и его коллеги (1998) предположили, что Карнотавр использовал свои рога подобно баранам. Они подсчитали, что мускулатура шеи была достаточно сильной, чтобы поглотить силу двух человек, сталкивающихся с головами спереди со скоростью 5,7 м / с каждый.[22] Фернандо Новас (2009) интерпретировал несколько особенностей скелета как приспособления для нанесения ударов головой.[В КАЧЕСТВЕ] Он предположил, что короткость черепа могла ускорить движения головы за счет уменьшения момент инерции, а мускулистая шея допускала сильные удары головой. Он также отметил повышенную жесткость и прочность позвоночника, который, возможно, развился, чтобы выдерживать удары, проводимые головой и шеей.[В]
Другие исследования показывают, что соперники Карнотавр не наносили быстрых ударов головой, а медленно толкались друг о друга верхней стороной черепа.[42][43] В 2009 году Маццетта и его коллеги утверждали, что рога могли быть устройством для распределения сил сжатия без повреждения мозга. Это подтверждается уплощенными верхними сторонами рогов, сильно сросшимися костями верхней части черепа и неспособностью черепа выдерживать быстрые удары головой.[42] Рафаэль Делькурт в 2018 году предположил, что рога можно было использовать либо для медленных ударов головой, либо для толкания, как это видно на современном примере. Морская игуана, или в ударах по шее и бокам противника, как это видно в современном жирафа.[35] Последняя возможность была ранее предложена для связанных Маджунгазавр в документе конференции 2011 года.[44]
Херардо Маццетта и его коллеги (1998) предполагают, что рога также могли использоваться для ранения или убийства мелкой добычи. Хотя сердечники рогов тупые, они могли иметь форму, аналогичную современным бычий рога, если был ороговевший покрытие. Однако это будет единственный зарегистрированный пример использования рогов в качестве охотничьего оружия у животных.[22]
Функция челюсти и диета

Анализ строения челюсти Карнотавр Маццетта и его коллеги в 1998, 2004 и 2009 годах предполагают, что животное было способно к быстрым, но не сильным укусам.[22][23][42] Быстрые укусы более важны, чем сильные, когда поимка мелкой добычи, как показали исследования современных крокодилы.[42] Эти исследователи также отметили высокую степень гибкости (кинезис ) внутри черепа и особенно нижней челюсти, чем-то похожего на современные змеи. Эластичность челюсти позволила бы Карнотавр заглатывать мелкие предметы добычи целиком. Кроме того, передняя часть нижней челюсти была шарнирной и, следовательно, могла двигаться вверх и вниз. При нажатии вниз зубы выступали бы вперед, позволяя Карнотавр колоть мелкую добычу; когда зубы были загнуты вверх, теперь выступающие назад зубы не давали пойманной жертве сбежать.[22] Маццетта и его коллеги также обнаружили, что череп способен противостоять силам, возникающим при рывке крупных предметов добычи.[42] Карнотавр поэтому могли питаться в основном относительно мелкой добычей, но также могли охотиться на крупных динозавров.[42] В 2009 году Маццетта и его коллеги оценили силу укуса примерно в 3341 ньютон.[42]
Эта интерпретация была подвергнута сомнению Франсуа Терриеном и его коллегами (2005), которые обнаружили, что сила укуса Карнотавр был вдвое больше Американский аллигатор, который может иметь самый сильный укус из всех живых четвероногий. Эти исследователи также отметили аналогии с современными Комодские драконы: the предел прочности при изгибе нижней челюсти линейно уменьшается по направлению к кончику, что указывает на то, что челюсти не подходят для высокоточной ловли мелкой добычи, а предназначены для нанесения рубящих ран, чтобы ослабить большую добычу. Как следствие, согласно этому исследованию, Карнотавр должно быть, в основном охотились на крупных животных, возможно, из засады.[45] Серрони и его коллеги в 2020 году утверждали, что гибкость ограничивалась нижней челюстью, в то время как утолщенная крыша черепа и окостенение нескольких черепных суставов предполагают, что у черепа не было кинезиса или было очень мало.[1]
Роберт Баккер (1998) обнаружили, что Карнотавр питались в основном очень крупной добычей, особенно зауроподы. Как он заметил, несколько приспособлений черепа - короткая морда, относительно маленькие зубы и крепкая задняя часть черепа (затылок ) - независимо развились в Аллозавр. Эти особенности предполагают, что верхняя челюсть использовалась как зазубренная дубинка для нанесения ран; большие зауроподы были бы ослаблены повторными атаками.[46]
Передвижение
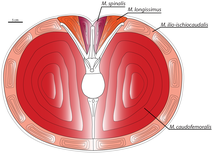

Маццетта и его коллеги (1998, 1999) предположили, что Карнотавр был быстрым бегуном, утверждая, что бедренная кость был адаптирован, чтобы выдерживать высокие изгибающие моменты во время бега. Способность ноги животного противостоять этим силам ограничивает его максимальную скорость. Беговые приспособления Карнотавр были бы лучше, чем у человека, хотя и не так хорошо, как у человека страус.[AU][47] Ученые подсчитали, что карнотавр имел максимальную скорость до 48–56 км (30–35 миль) в час.[48]
В динозаврах, наиболее важным локомоторной мышца находится в хвосте. Эта мышца, называемая каудофеморальный, прикрепляется к четвертый вертел, выступающий гребень на бедренной кости, который при сокращении тянет бедренную кость назад. Скотт Персонс и Фил Карри (2011) отмечают, что в хвостовых позвонках Карнотаврхвостовые ребра не выступали горизонтально («Т-образно»), а были наклонены к вертикальной оси позвонков, образуя «V». Это обеспечило бы дополнительное пространство для каудофеморальной мышцы, большей, чем у любого другого теропод - мышечная масса была рассчитана от 111 до 137 килограммов (от 245 до 302 фунтов) на ногу. Следовательно, Карнотавр мог быть одним из самых быстрых крупных теропод.[28] В то время как каудофеморальная мышца была увеличена, эпаксиальные мышцы расположенные выше хвостовых ребер были бы пропорционально меньше. Эти мышцы, называемые длинная мышца и позвоночник мышцы, отвечали за движение и устойчивость хвоста. Чтобы сохранить стабильность хвоста, несмотря на сокращение этих мышц, хвостовые ребра несут выступающие вперед отростки, переплетая позвонки друг с другом и с тазом, придавая хвосту жесткость. Как следствие, способность делать крутые повороты была бы уменьшена, потому что бедро и хвост нужно было поворачивать одновременно, в отличие от других теропод.[28]
Мозг и чувства
Черрони и Паулина-Карабахал в 2019 году использовали компьютерную томографию для изучения эндокраниальной полости, в которой находился мозг. Объем внутричерепной полости составлял 168,8 см3, хотя мозг мог заполнить лишь часть этого пространства. Авторы использовали две разные оценки размера мозга, предполагая, что размер мозга составляет 50% и 37% эндокраниальной полости соответственно. Это приводит к коэффициент энцефализации рептилий (показатель интеллекта) больше, чем у связанных Маджунгазавр но меньше, чем в тираннозавриды. В шишковидная железа, который производит гормоны, мог быть меньше, чем у других абелизаврид, на что указывает небольшое расширение твердой мозговой оболочки - пространство над передним мозгом, в котором, как полагают, располагалась шишковидная железа.[24]
В обонятельные луковицы, в которых размещалось обоняние, были большими, а зрительные доли, которые отвечали за прицел, были относительно небольшими. Это указывает на то, что обоняние могло быть лучше развито, чем зрение, тогда как у современных птиц наблюдается обратное. Передний конец обонятельных трактов и луковиц был изогнут вниз, что характерно только для Индозавр; у других абелизаврид эти структуры ориентированы горизонтально. Согласно гипотезе Серрони и Паулины-Карабахал, эта кривизна вниз вместе с большим размером луковиц может указывать на то, что Карнотавр полагались больше на обоняние, чем другие абелизавриды. В хлопья, доля мозга, которая, как считалось, связана со стабилизацией взгляда (координация между глазами и телом), была большой в Карнотавр и другие южноамериканские абелизавриды. Это могло указывать на то, что в этих формах часто использовались быстрые движения головы и тела. Слух мог быть плохо развит в Карнотавр и другие абелизавриды, на что указывает короткий лагена из внутреннее ухо. Диапазон слышимости был оценен ниже 3 кГц.[24]
Возраст и палеосреда

Первоначально скалы, в которых Карнотавр были отнесены к верхней части Формация Горро Фриджио, которому приблизительно 100 миллионов лет (Альбианский или же Сеноманский сцена).[5][СРЕДНИЙ] Позже выяснилось, что они относятся к гораздо более молодым Формация Ла Колония,[13] датируется от 83,6 до 72,1 миллиона лет назад (Поздний мел, Кампанский к Маастрихтский этапы).[1] Новас в книге 2009 года дал более узкий временной интервал от 72 до 69,9 миллиона лет назад (ниже Маастрихтский сцена).[AW] Карнотавр следовательно, был известен последний южноамериканский абелизаврид.[28] К позднему меловому периоду Южная Америка была уже изолирована как от Африки, так и от Северной Америки.[49]
Формация Ла-Колония обнажена на южном склоне холма. Северный Патагонский массив.[50] Большинство окаменелостей позвоночных, в том числе Карнотавр, происходят из средней части формации (называемой ассоциация средних фаций).[50] Эта часть, вероятно, представляет собой отложения среды эстуарии, приливные отмели или же прибрежные равнины.[50] Климат был бы сезонным с засушливыми и влажными периодами.[50] Наиболее часто собираемые позвоночные включают: цератодонтид двоякодышащая рыба, черепахи, крокодилы, плезиозавры, динозавры, ящерицы, змеи и млекопитающие.[51] Некоторые из найденных змей принадлежат семьям Boidae и Madtsoidae, такие как Аламитофис аргентинус.[52] Черепахи представлены минимум пятью таксоны, четыре из Chelidae (Pleurodira ) и один из Meiolaniidae (Криптодира ).[53] Среди морских рептилий есть плезиозавр. Sulcusuchus erraini семьи Поликотилиды.[53] Млекопитающие представлены Reigitherium bunodontum, который считался первой пластинкой южноамериканского докодонт,[50] и Argentodites Coloniensis, возможно, из Мультитуберкулята.[54] В 2011 году открытие нового энантиорнитин была объявлена птица из формации Ла Колония.[55]
Смотрите также
Примечания
- ^ п. 276 в Novas (2009)[2]
- ^ п. 2 в Бонапарте (1990)[4]
- ^ п. 2 в Бонапарте (1990)[4]
- ^ п. 2 в Бонапарте (1990)[4]
- ^ п. 191 в Каррано и Сэмпсон (2008)[8]
- ^ п. 191 в Каррано и Сэмпсон (2008)[8]
- ^ п. 3 в Бонапарте (1990)[4]
- ^ п. 38 в Бонапарте (1990)[4]
- ^ п. 162 в Juárez Valieri et al. (2010)[18]
- ^ п. 191 в Каррано и Сэмпсон (2008)[8]
- ^ п. 162 в Juárez Valieri et al. (2010)[18]
- ^ п. 163 в Хуаресе Вальери и др. (2010)[18]
- ^ п. 556 в Calvo et al. (2004)[20]
- ^ п. 191 в Каррано и Сэмпсон (2008)[8]
- ^ п. 30 в Бонапарте (1990)[4]
- ^ п. 187 в Mazzetta et al. (1998)[22]
- ^ п. 79 в Mazzetta et al. (2004)[23]
- ^ п. 276 в Novas (2009)[2]
- ^ п. 28–32 в Бонапарте (1990)[4]
- ^ п. 8 в Бонапарте (1990)[4]
- ^ п. 191 в Каррано и Сэмпсон (2008)[8]
- ^ п. 4–5 в Бонапарте (1990)[4]
- ^ п. 5 в Бонапарте (1990)[4]
- ^ п. 3 в Бонапарте (1990)[4]
- ^ п. 191 в Mazzetta et al. (1998)[22]
- ^ п. 255 в Novas (2009)[2]
- ^ п. 6 в Бонапарте (1990)[4]
- ^ п. 6 в Бонапарте (1990)[4]
- ^ п. 6 в Бонапарте (1990)[4]
- ^ п. 6 в Бонапарте (1990)[4]
- ^ п. 191 в Каррано и Сэмпсон (2008)[8]
- ^ стр. 257 в Novas (2009)[2]
- ^ п. 1276 в Ruiz et al. (2011)[29]
- ^ п. 32 в Бонапарте (1990)[4]
- ^ п. 32 в Бонапарте (1990)[4]
- ^ п. 3 в Бонапарте (1990)[4]
- ^ стр. 264–299 в Novas (2009)[2]
- ^ стр. 264–299 в Novas (2009)[2]
- ^ п. 32 в Бонапарте (1990)[4]
- ^ п. 276–277 в Novas (2009)[2]
- ^ стр. 256–261 в Novas (2009)[2]
- ^ стр. 188–189 и 202 в Carrano and Sampson (2008).[8]
- ^ п. 202 в Каррано и Сэмпсон (2008)[8]
- ^ п. 202 в Каррано и Сэмпсон (2008)[8]
- ^ стр. 259–261 в Novas (2009)[2]
- ^ стр. 260–261 в Novas (2009)[2]
- ^ п. 186 и 190 в Mazzetta et al. (1998)[22]
- ^ п. 3 в Бонапарте (1990)[4]
- ^ п. 276 в Novas (2009)[2]
Рекомендации
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты Cerroni, M. A .; Canale, J. I .; Новас, Ф. Э. (2020). "Череп Carnotaurus sastrei Бонапарт в 1985 году снова: идеи черепно-лицевых костей, неба и нижней челюсти ". Историческая биология: 1–42. Дои:10.1080/08912963.2020.1802445.
- ^ а б c d е ж грамм час я j k Новас, Фернандо Э. (2009). Эпоха динозавров в Южной Америке. Блумингтон: Издательство Индианского университета. ISBN 978-0-253-35289-7.
- ^ а б Сальгадо, Леонардо; Бонапарт, Хосе Ф. (1991). "Un nuevo sauropodo Dicraeosauridae, Amargasaurus cazaui ген. et sp. nov., de la Formacion La Amarga, Neocomiano de la Provincia del Neuquén, Аргентина ". Амегиниана (на испанском). 28 (3–4): 334.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс Бонапарт, Хосе Ф .; Novas, Fernando E .; Кориа, Родольфо А. (1990). "Carnotaurus sastrei Бонапарт, рогатый, легко сложенный карнозавр из среднего мела Патагонии » (PDF). Вклад в науку. 416: 1–41. Архивировано из оригинал (PDF) 21 июля 2010 г.
- ^ а б c d е Бонапарт, Хосе Ф. (1985). «Рогатый меловой карнозавр из Патагонии». Национальное географическое исследование. 1 (1): 149–151.
- ^ а б Паулина Карабахал, Ариана (2011). "Анатомия черепной коробки Carnotaurus sastrei (Theropoda: Abelisauridae) из верхнего мела Патагонии ". Журнал палеонтологии позвоночных. 31 (2): 378–386. Дои:10.1080/02724634.2011.550354. S2CID 129933997.
- ^ а б c d е ж грамм час я Czerkas, Stephen A .; Черкас, Сильвия Дж. (1997). «Покровы и восстановление жизни карнотавра». В Wolberg, D. I .; Пень, E .; Розенберг, Г. Д. (ред.). Динофест Интернэшнл. Академия естественных наук, Филадельфия. С. 155–158.
- ^ а б c d е ж грамм час я Каррано, Мэтью Т .; Сэмпсон, Скотт Д. (январь 2008 г.). «Филогения Ceratosauria (Dinosauria: Theropoda)». Журнал систематической палеонтологии. 6 (2): 183–236. Дои:10.1017 / S1477201907002246. S2CID 30068953.
- ^ а б c d Пол, Грегори С. (1988). Хищные динозавры мира. стр.284–285. ISBN 978-0-671-61946-6.
- ^ Йонг, Эд (18 октября 2011 г.). "Бутч хвост сделал Карнотавра чемпионом по спринтерам динозавров". Национальная география. Получено 31 июля, 2019.
- ^ Хедден, Хайме А. (19 сентября 2006 г.). "Re: Carnotaurus sastrei этимология". Список рассылки динозавров.
- ^ Бонапарт, Хосе Ф. (1991). «Семейства гондванских теропод Abelisauridae и Noasauridae». Историческая биология. 5: 1. Дои:10.1080/10292389109380385.
- ^ а б Бонапарт, Хосе Ф. (1996). «Меловые четвероногие Аргентины». Münchener Geowissenschaftliche Abhandlung. А (30): 89.
- ^ Glut, Дональд Ф. (1997). "Карнотавр". Динозавры, энциклопедия. Джефферсон, Северная Каролина: издательство McFarland & Company, Inc. стр.256–259. ISBN 978-0-375-82419-7.
- ^ а б Glut, Дональд Ф. (2003). "Карнотавр". Динозавры: Энциклопедия. 3-е дополнение. Джефферсон, Северная Каролина: McFarland & Company, Inc., стр.274–276. ISBN 978-0-7864-1166-5.
- ^ Glut, Дональд Ф. (2000). "Карнотавр". Динозавры: Энциклопедия. 1-е приложение. Джефферсон, Северная Каролина: McFarland & Company, Inc., стр. 165–167. ISBN 978-0-7864-0591-6.
- ^ а б Кандейро, Карлос Роберту душ Аньос; Мартинелли, Агустин Гильермо. «Abelisauroida и carchardontosauridae (тероподы, динозавры) в меловом периоде Южной Америки. Палеогеографические и геокронологические последствия». Уберландия. 17 (33): 5–19.
- ^ а б c Хуарес Вальери, Рубен Д .; Porfiri, Juan D .; Кальво, Хорхе О. (2010). "Новая информация о Экриксинатозавр novasi Calvo et al. 2004 г. - гигантский массивный абелизавроид из «среднего мела» Патагонии ». Палеонтология и динозавры в Латинской Америке: 161–169.
- ^ а б c Пол, Грегори С. (2010). Принстонский полевой путеводитель по динозаврам. Издательство Принстонского университета. ISBN 9780691137209.
- ^ Кальво, Хорхе О .; Рубилар-Роджерс, Дэвид; Морено, Карен (2004). «Новый Abelisauridae (Dinosauria: Theropoda) с северо-запада Патагонии» (PDF). Амегиниана. 41 (4): 555–563. Архивировано из оригинал (PDF) 9 марта 2012 г.
- ^ Grillo, O.N .; Делькур, Р. (2016). «Аллометрия и длина тела абелизавроидных теропод: Pycnonemosaurus nevesi новый король ". Меловые исследования. 69: 71–89. Дои:10.1016 / j.cretres.2016.09.001.
- ^ а б c d е ж грамм Mazzetta, Gerardo V.; Fariña, Richard A.; Vizcaíno, Sergio F. (1998). "On the palaeobiology of the South American horned theropod Carnotaurus sastrei Bonaparte" (PDF). Гайя. 15: 185–192.
- ^ а б Mazzetta, Gerardo V.; Кристиансен, Пер; Fariña, Richard A. (2004). "Giants and Bizarres: Body size of some southern South American Cretaceous dinosaurs" (PDF). Историческая биология. 16 (2): 71–83. CiteSeerX 10.1.1.694.1650. Дои:10.1080/08912960410001715132. S2CID 56028251.
- ^ а б c d Cerroni, Mauricio A.; Paulina-Carabajal, Ariana (2019). "Novel information on the endocranial morphology of the abelisaurid theropod Carnotaurus sastrei". Comptes Rendus Palevol. 18 (8): 985–995. Дои:10.1016/j.crpv.2019.09.005.
- ^ а б Sampson, Scott D.; Witmer, Lawrence M. (2007). "Craniofacial Anatomy of Majungasaurus crenatissimus (Theropoda: Abelisauridae) From the Late Cretaceous of Madagascar". Журнал палеонтологии позвоночных. 27 (sp8): 95–96. Дои:10.1671/0272-4634(2007)27[32:CAOMCT]2.0.CO;2.
- ^ а б c Hartman, Scott (2012). "Carnotaurus – delving into self-parody?". Получено 7 декабря, 2012.
- ^ Méndez, Ariel (2014). "The cervical vertebrae of the Late Cretaceous abelisaurid dinosaur Carnotaurus sastrei" (PDF). Acta Palaeontologica Polonica. 59 (1): 99–107. Дои:10.4202/app.2011.0129. S2CID 67792084. Архивировано из оригинал (PDF) 3 апреля 2013 г.. Получено 30 декабря, 2012.
- ^ а б c d е Persons, W.S.; Currie, P.J. (2011). Farke, Andrew Allen (ed.). "Dinosaur Speed Demon: The caudal musculature of Carnotaurus sastrei and implications for the evolution of South American abelisaurids". PLOS ONE. 6 (10): e25763. Bibcode:2011PLoSO...625763P. Дои:10.1371/journal.pone.0025763. ЧВК 3197156. PMID 22043292.
- ^ а б c d Ruiz, Javier; Torices, Angélica; Serrano, Humberto; López, Valle (2011). "The hand structure of Carnotaurus sastrei (Theropoda, Abelisauridae): implications for hand diversity and evolution in abelisaurids" (PDF). Палеонтология. 54 (6): 1271–1277. Дои:10.1111/j.1475-4983.2011.01091.x.
- ^ Agnolin, Federico L.; Chiarelli, Pablo (June 2010). "The position of the claws in Noasauridae (Dinosauria: Abelisauroidea) and its implications for abelisauroid manus evolution". Paläontologische Zeitschrift. 84 (2): 293–300. Дои:10.1007/s12542-009-0044-2. S2CID 84491924.
- ^ Senter, P. (2010). "Vestigial skeletal structures in dinosaurs". Журнал зоологии. 280 (4): 60–71. Дои:10.1111/j.1469-7998.2009.00640.x.
- ^ а б c Канале, Хуан I .; Scanferla, Carlos A.; Agnolin, Federico; Novas, Fernando E. (2009). "New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods". Naturwissenschaften. 96 (3): 409–14. Bibcode:2009NW.....96..409C. Дои:10.1007/s00114-008-0487-4. HDL:11336/52024. PMID 19057888. S2CID 23619863.
- ^ а б Coria, Rodolfo A.; Chiappe, Луис М .; Dingus, Lowell (2002). "A new close relative of Carnotaurus sastrei Bonaparte 1985 (Theropoda: Abelisauridae) from the Late Cretaceous of Patagonia". Журнал палеонтологии позвоночных. 22 (2): 460. Дои:10.1671/0272-4634(2002)022[0460:ANCROC]2.0.CO;2.
- ^ Ezcurra, Martín D .; Agnolin, Federico L.; Novas, Fernando E. (2010). "An abelisauroid dinosaur with a non-atrophied manus from the Late Cretaceous Pari Aike Formation of southern Patagonia" (PDF). Zootaxa. 2450: 14. Дои:10.11646/zootaxa.2450.1.1.
- ^ а б c Delcourt, Rafael (2018). "Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers". Научные отчеты. 8 (1): 9730. Bibcode:2018NatSR...8.9730D. Дои:10.1038/s41598-018-28154-x. ЧВК 6021374. PMID 29950661.
- ^ Серено, Пол К.; Wilson, Jeffrey A.; Conrad, Jack L. (July 7, 2004). "New dinosaurs link southern landmasses in the Mid-Cretaceous". Труды Королевского общества B: биологические науки. 271 (1546): 1325–1330. Дои:10.1098/rspb.2004.2692. ЧВК 1691741. PMID 15306329.
- ^ Tykoski, Ronald B.; Rowe, Timothy (2004). "Ceratosauria". In Weishampel, David B.; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (Второе изд.). Беркли: Калифорнийский университет Press. п.65. ISBN 978-0-520-24209-8.
- ^ Wilson, Jeffrey A.; Sereno, Paul C.; Srivastava, Suresh; Bhatt, Devendra K.; Хосла, Ашу; Sahni, Ashok (2003). "A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India". Вклады музея палеонтологии. 31 (1): 25. HDL:2027.42/48667.
- ^ Wang, S .; Stiegler, J .; Amiot, R .; Ван, X .; Du, G.-H .; Clark, J.M .; Сюй, X. (2017). "Extreme ontogenetic changes in a ceratosaurian theropod" (PDF). Текущая биология. 27 (1): 144–148. Дои:10.1016 / j.cub.2016.10.043. PMID 28017609. S2CID 441498.
- ^ Sereno, Paul (2005). "Carnotaurinae". Taxon Search. Архивировано из оригинал 16 мая 2012 г.. Получено 29 декабря, 2012.
- ^ Sereno, Paul (2005). "Carnotaurini". Taxon Search. Архивировано из оригинал 16 мая 2012 г.. Получено 29 декабря, 2012.
- ^ а б c d е ж грамм час Mazzetta, Gerardo V.; Cisilino, Adrián P.; Blanco, R. Ernesto; Calvo, Néstor (2009). "Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei". Журнал палеонтологии позвоночных. 29 (3): 822–830. Дои:10.1671/039.029.0313. S2CID 84565615.
- ^ Chure, Daniel J. (1998). "On the orbit of theropod dinosaurs". Гайя. 15: 233–240.
- ^ Snively, Eric; Cotton, John R.; Witmer, Lawrence; Ridgely, Ryan; Theodor, Jessica (2011). "Finite element comparison of cranial sinus function in the dinosaur Маджунгазавр and head-clubbing giraffes". Summer Bioengineering Conference. 54587. Американское общество инженеров-механиков. pp. 1075–1076.
- ^ Therrien, François; Хендерсон, Дональд; Ruff, Christopher (2005). "Bite Me – Biomechanical Models of Theropod Mandibles and Implications for Feeding Behavior". В Карпентер, Кеннет (ред.). The carnivorous dinosaurs. Издательство Индианского университета. стр.179 –198, 228. ISBN 978-0-253-34539-4.
- ^ Bakker, Robert T. (1998). "Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues" (PDF). Гайя. 15: 145–158.
- ^ Mazzetta, Gerardo V.; Farina, Richard A. (1999). "Estimacion de la capacidad atlética de Amargasaurus cazaui Salgado y Bonaparte, 1991, y Carnotaurus sastrei Bonaparte, 1985 (Saurischia, Sauropoda-Theropoda)". XIV Jornadas Argentinas de Paleontologia de Vertebrados, Ameghiniana (на испанском). 36 (1): 105–106.
- ^ "Predatory dinosaur was fearsomely fast". CBC Новости. October 21, 2011. Получено 22 апреля, 2017.
- ^ Le Loeuff, Jean (1997). "Biogeography". In Padian, Kevin; Currie, Philip J. (eds.). Encyclopedia of dinosaurs. Сан-Диего: Academic Press. стр.51 –56. ISBN 978-0-12-226810-6.
- ^ а б c d е Pascual, Rosendo; Goin, Francisco J .; González, Pablo; Ardolino, Alberto; Puerta, Pablo F. (2000). «Докодонт высокоразвитого происхождения из патагонского позднего мела: эволюционное значение для гондванских млекопитающих». Геодиверситас. 22 (3): 395–414.
- ^ Sterli, Juliana; De la Fuente, Marcelo S. (2011). "A new turtle from the La Colonia Formation (Campanian–Maastrichtian), Patagonia, Argentina, with remarks on the evolution of the vertebral column in turtles". Палеонтология. 54 (1): 65. Дои:10.1111/j.1475-4983.2010.01002.x.
- ^ Albino, Adriana M. (2000). "New record of snakes from the Cretaceous of Patagonia (Argentina)". Геодиверситас. 22 (2): 247–253.
- ^ а б Гаспарини, Зульма; De la Fuente, Marcelo (2000). "Tortugas y Plesiosaurios de la Formación La Colonia (Cretácico Superior) de Patagonia, Argentina". Revista Española de Paleontología (на испанском). 15 (1): 23.
- ^ Kielan−Jaworowska, Zofia; Ortiz−Jaureguizar, Edgardo; Vieytes, Carolina; Pascual, Rosendo; Goin, Francisco J. (2007). "First ?cimolodontan multi−tuberculate mammal from South America" (PDF). Acta Palaeontologica Polonica. 52 (2): 257–262.
- ^ Lawver, Daniel R .; Debee, Aj M.; Кларк, Джулия А .; Rougier, Guillermo W. (January 1, 2011). "A New Enantiornithine Bird from the Upper Cretaceous La Colonia Formation of Patagonia, Argentina". Летопись музея Карнеги. 80 (1): 35–42. Дои:10.2992/007.080.0104. S2CID 85015365.
внешняя ссылка
- Укус Карнотавр at Universidad Nacional de Mar del Plata. (на испанском)
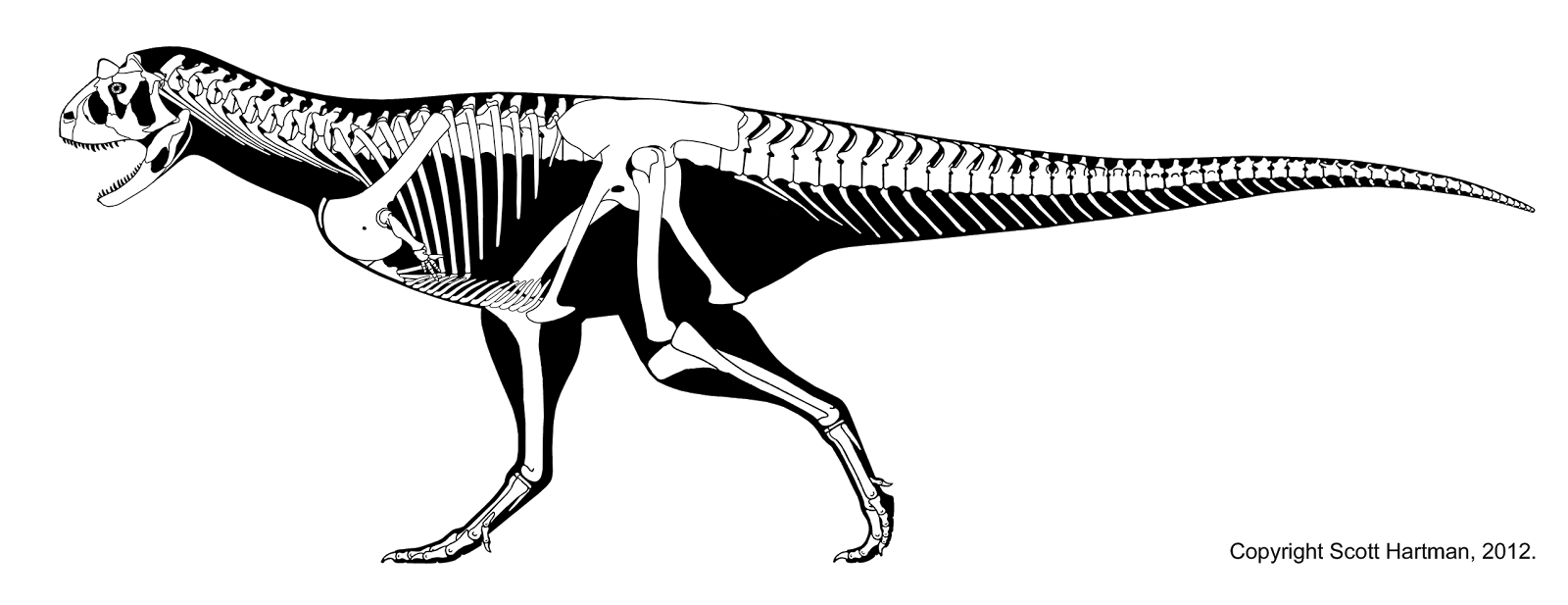
- Skeletal reconstruction by Scott Hartman
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||














{kind=link}
| Авторитетный контроль |
|---|