Homo naledi - Homo naledi
| Homo naledi | |
|---|---|
 | |
| 737 известных элементов Х. наледи | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Haplorhini |
| Инфразаказ: | Симиобразные |
| Семья: | Гоминиды |
| Подсемейство: | Гоминины |
| Племя: | Гоминини |
| Род: | Гомо |
| Разновидность: | †Х. наледи |
| Биномиальное имя | |
| †Homo naledi Бергер и другие., 2015 | |
 | |
| Расположение Пещера восходящей звезды в Колыбель человечества, Южная Африка | |
Homo naledi это разновидность архаичный человек обнаружен в Пещера восходящей звезды, Колыбель человечества, Южная Африка Средний плейстоцен 335 000–236 000 лет назад. Первоначальная находка включает 1550 экземпляров, представляющих 737 различных элементов и по крайней мере 15 разных людей. Несмотря на это исключительно большое количество экземпляров, их отнесение к другим Гомо остается неясным.
Наряду с сходством с современными Гомо, они имеют несколько общих черт с Австралопитек и рано Гомо а также (мозаичная анатомия), особенно небольшой объем черепа 465–610 см3 (28,4–37,2 куб. Дюймов), по сравнению с 1270–1330 см3 (78–81 куб. Дюймов) у современного человека. По оценкам, их средний рост составляет 143,6 см (4 фута 9 дюймов), а вес - 39,7 кг (88 фунтов), что дает небольшой коэффициент энцефализации из 4.5. Тем не менее, Х. наледи анатомия мозга, похоже, была похожа на современную Гомо, что может указывать на равную когнитивную сложность. Столь долгое существование людей с маленьким мозгом среди современников с большим мозгом пересматривает предыдущую концепцию, что больший мозг обязательно приведет к эволюционному преимуществу, а их мозаичная анатомия значительно расширяет известный диапазон вариаций для рода.
Х. наледи анатомия показывает, что, хотя они были способны путешествовать на большие расстояния с походкой и походкой, подобными человеческим, они были более древесный чем другие Гомо, лучше приспособлен к лазанию и сдерживающее поведение на деревьях, чем бег на выносливость. Анатомия зубов предполагает употребление твердой пищи, покрытой частицами, такими как пыль или грязь. Хотя они не были связаны с каменными инструментами или какими-либо признаками материальной культуры, они, похоже, были достаточно ловкими, чтобы производить инструменты и обращаться с ними, и, вероятно, производили Рано или же Середина Каменный век отрасли. Кроме того, было предположено, что спорно эти люди были даны погребальные обряды, а также были проведены в и помещают в камеру.
Таксономия
Открытие

В октябре 2013 г. спелеологи Рик Хантер и Стив Такер под руководством южноафриканского палеоантрополога Ли Роджерс Бергер, обнаружил камеру Диналеди в Пещера восходящей звезды в Колыбель человечества, Южная Африка, содержит несколько окаменелостей гомининов.[2] Камера расположена примерно в 80 м (260 футов) от входа, а главный проход имеет длину около 10 м (33 фута) и 25–50 см (9,8–19,7 дюйма) в самом узком месте и находится в нижней части 12 м (39 футов) перепад высот.[3] Всего более 1550 костей, принадлежащих как минимум пятнадцати особям (9 незрелых и 6 взрослых[4]) были восстановлены из глина -богатые отложения. Бергер и его коллеги опубликовали результаты в 2015 году.[5]
Окаменелости представляют 737 анатомических элементов, включая части черепа, челюсти, ребер, зубов, конечностей и костей внутреннего уха, от старых, взрослых, молодых и младенческих людей. Есть также некоторые сочлененный или почти сочлененные элементы, включая череп с челюстной костью и почти целые руки и ноги.[5][3] По количеству представителей обоих полов в разных возрастных группах это самое богатое скопление связанных ископаемых гомининов, обнаруженное в Африке. Помимо Sima de los Huesos коллекция и позже Неандерталец и современных человеческих образцов, место раскопок наиболее полно отображает элементы скелета на протяжении всей жизни и от множества людей в летописи окаменелостей гомининов.[5]

В образец голотипа, DH1, включает частичную вилку свода черепа (верхняя часть черепа), частичная верхняя челюсть, и почти полностью челюсть. В паратипы, DH2-5, все включают частичный свод черепа. Бергер и его коллеги назвали вид Homo naledi, название вида означает "звезда" в Сото язык, потому что останки прибыли из пещеры Восходящей звезды.[5]
Останки по крайней мере еще трех человек - двух взрослых и ребенка - были обнаружены в камере Леседи пещеры. Джон Хоукс и коллеги в 2017 году.[6]
Классификация
В 2017 году останки Диналеди были датированы 335–236 000 лет назад в Средний плейстоцен, с помощью электронный спиновой резонанс (СОЭ) и уран-торий (U-Th) датировка по 3 зубам, а U-Th и палеомагнитный датировка отложений, в которых они были отложены.[1] Ранее считалось, что окаменелости датируются 1-2 миллиона лет назад.[5][7][8][2] потому что гоминины с таким же мелким мозгом ранее не были известны в Африке с недавнего времени.[9] (менее умные Homo floresiensis Индонезии жили на изолированном острове и, по-видимому, вымерли вскоре после прибытия современных людей.[10]) Способность гоминина с таким маленьким мозгом выживать так долго среди людей с большим мозгом. Гомо значительно пересматривает предыдущие концепции эволюция человека и представление о том, что больший мозг обязательно приведет к эволюционному преимуществу.[9] Их мозаичная анатомия также значительно расширяет диапазон вариаций для рода.[11]
Х. наледи предполагается, что он очень рано отошел от современных Гомо. Неясно, разветвились ли они примерно во время H. habilis, Х. рудольфенсис, и А. седиба; площадь сестринский таксон к H. erectus и современник с большим мозгом Гомо; или являются сестринским таксоном потомкам H. предшественник (современные люди и неандертальцы). Это означало бы, что они отошли от современных Гомо самое позднее, до 900 000 лет назад, и, возможно, еще до Плиоцен. Также возможно, что их предки были определены после скрещивания между Гомо и поздние австралопитеки.[9] Глядя на череп, Х. наледи имеет ближайший родство к H. erectus.[11]
Неясно, если эти Х. наледи были изолированной популяцией в Колыбели Человечества, или если они жили по всей Африке. Если последнее, то несколько окаменелостей грацильных гомининов по всей Африке, которые традиционно классифицируются как поздние H. erectus потенциально может представлять Х. наледи образцы.[12]

Анатомия
Череп

Два самца Х. наледи черепа из камеры Диналеди имели черепные объемы около 560 см3 (34 у.е. дюйма) и два женских черепа 465 см3 (28,4 куб. Дюймов). Мужчина Х. наледи череп из камеры Леседи имел объем черепа 610 см3 (37 куб. Дюймов). Экземпляры Dinaledi больше похожи на черепную емкость австралопитеков; для сравнения, H. erectus в среднем около 900 см3 (55 куб. Дюймов),[6] и современные люди 1270 и 1130 см3 (78 и 69 куб. Дюймов) для мужчин и женщин соответственно.[13] Однако образец Леседи находится в пределах H. habilis и Он. georgicus. В коэффициент энцефализации из Х. наледи был оценен в 4,5, что соответствует пигмею H. floresiensis, но заметно меньше всех остальных Гомо (современный Гомо были все выше 6).[14] Тем не менее, форма черепа больше похожа на Гомо, при более тонкой форме наличие временный и затылочный доли головного мозга и уменьшенные посторбитальное сужение (череп за глазницами не сужается).[5][15] В лобная доля морфология более или менее одинакова у всех Гомо мозги, несмотря на размер, который отличается от Австралопитек, и был вовлечен в производство инструментов, развитие языка и социальность. Неясно, если Х. наледи унаследовал небольшой размер мозга от последнего общего Гомо предок, или, если он возник вторично позже.[15]
Подобно современным людям, но в отличие от ископаемых гомининов (включая южноафриканских австралопитеков, H. erectus, и неандертальцы), постоянный 2-й коренной зуб прорезывал сравнительно поздно в жизни, появляясь вместе с премолярами, а не раньше, что указывает на более медленное созревание, необычно сопоставимое с современными людьми.[16] Скорость образования передних зубов также больше всего похожа на современного человека.[17] Общий размер и форма моляров больше всего напоминают три неустановленных. Гомо образцы из местных Сварткранс и Восточной Африки Кооби Фора Пещеры и по размеру (но не по форме) похожи на плейстоценовые. Х. сапиенс. Шейки коренных зубов пропорционально аналогичны шейкам коренных зубов. A. afarensis и Парантроп.[18] В отличие от современных людей и современных Гомо, Х. наледи не имеет нескольких дополнительных стоматологических функций, и часто встречаются лица, представляющие основные куспиды, а именно метакон (средняя линия со стороны языка) и лицемер (справа со стороны губ) на 2-м и 3-м молярах и Y-образный гипоконулид (гребень со стороны губ к щеке) на всех 3 молярах. Тем не менее, Х. наледи также имеет много стоматологического сходства с современными Гомо.[19]
В наковальня (а кость среднего уха ) больше похож на шимпанзе, горилл и Парантроп чем Гомо.[20] Нравиться H. habilis и H. erectus, Х. наледи имеет хорошо развитую надбровную дугу с трещиной, тянущейся поперек самого гребня, и H. erectus ярко выраженный затылочная бугорка. Х. наледи имеет некоторые черты лица с Х. рудольфенсис.[19]
Строить

В Х. наледи По оценкам, образцы в среднем имели рост около 143,6 см (4 фута 9 дюймов) и весили 39,7 кг (88 фунтов). Эта масса тела занимает промежуточное положение между тем, что обычно наблюдается в Австралопитек и Гомо разновидность. Как и другие Гомо, мужской и женский Х. наледи были примерно одного размера, самцы в среднем примерно на 20% крупнее самок.[14] Ювенильная особь DH7 скелетно соответствует скорости роста, аналогичной более быстрым обезьяноподобным траекториям MH1 (А. седиба) и Мальчик туркана (Х. эргастер ). Однако, поскольку развитие зубов очень похоже на развитие зубов у современного человека, более низкая скорость созревания полностью не исключена. При более высокой скорости роста DH7 умер бы в 8–11 лет, но при более медленном росте DH7 умер бы в 11–15 лет.[21]
Касательно позвоночник, только 10 и 11 грудные позвонки (в области груди) сохранились предположительно от одного человека, которые пропорционально похожи на таковых современных Гомо, хотя и являются самыми маленькими из всех гомининов. Два поперечных отростка позвонка, которые выступают по диагонали, больше всего похожи на таковые у неандертальцев. В нервные каналы внутри пропорционально большие, похожие на современных людей, неандертальцев и Он. georgicus. 11-е ребро прямое, как у A. afarensis, а 12-е ребро в сечении прочное, как у неандертальцев. Как и у неандертальцев, 12-е ребро, похоже, поддерживало сильную межреберные мышцы выше, и сильный квадратная мышца поясницы ниже. Однако, в отличие от неандертальцев, была слабая привязанность к диафрагма. В целом это Х. наледи образец, кажется, был маленьким по сравнению с другими Гомо, хотя неясно, является ли этот единственный экземпляр репрезентативным для данного вида.[22]
Плечи больше похожи на плечи австралопитеков, с лопатка расположены выше на спине и дальше от средней линии, короткие ключицы, и небольшой перекрут плечевой кости или его нет.[5] Возвышенные плечевые и ключичные кости указывают на узкую грудную клетку.[22] В таз и ноги имеют черты, напоминающие Австралопитек, в том числе сжатые передне (спереди назад) бедренные шейки, сжатие медиолатерально (слева направо) большеберцовые кости, и несколько круговой малоберцовая шейка;[23][24] которые указывают на широкий живот. Эта комбинация помешала бы эффективному бег на выносливость в Х. наледи, В отличие от H. erectus и потомки. Вместо, Х. наледи кажется, было больше древесный.[22]
Конечности


В пястная кость большой палец был хорошо развит, он использовался для удержания и манипулирования большими предметами, и имел сильные гребни для поддержки его opponens pollicis muscle используется в прецизионном захвате, и его тенарные мышцы. Это больше похоже на другие Гомо чем Австралопитек. Х. наледи похоже, был сильный длинный сгибатель большого пальца как у современных людей, с человеческими ладонями и подушечками пальцев, которые важны для сильного захвата между большим и пальцами. Однако в отличие от Гомо, то Х. наледи Пястный сустав большого пальца сравнительно мал по сравнению с длиной большого пальца, а большой палец фаланговый сустав уплощен. Дистальная кость фаланги большого пальца крепкая и пропорционально более похожа на кости H. habilis и P. robustus.[25]
Пястные кости других пальцев имеют общие с современными людьми и неандертальцами приспособления для манипулирования объектами и манипулирования ими, а лучезапястный сустав в целом сопоставим с таковым у современных людей и неандертальцев. Напротив, проксимальные фаланги изогнуты и почти идентичны таковым у A. afarensis и H. habilis, что трактуется как приспособление к лазанию и сдерживающее поведение. Такая кривизна более выражена у взрослых, чем у молодых, что позволяет предположить, что взрослые лазали так же или больше, чем молодые, и такое поведение было обычным. Пальцы также пропорционально длиннее, чем у любого другого ископаемого гоминина (кроме древесного Ardipithecus ramidus и современный образец человека из Пещера Кафзех, Израиль), что также согласуется со скалолазным поведением.[25]
Х. наледи был двуногий и встал прямо.[5] Как и другие Гомо, у них были сильные вставка для ягодичная мышца мышцы, четко очерченные Linea Aspera (гребень, спускающийся по задней поверхности бедра), толстый надколенники, длинные голени и изящные малоберцовые кости. Это указывает на то, что они были способны путешествовать на большие расстояния.[24] В Х. наледи нога была похожа на ногу современного человека и других Гомо, с приспособлениями для двуногие и походка, подобная человеку. Тем не менее пяточная кость имеет низкую ориентацию, сравнимую с ориентацией человекообразных человекообразных обезьян, и лодыжка имеет низкий наклон, что, возможно, указывает на то, что ступня была бы немного жестче во время фазы опоры при ходьбе до того, как ступня оттолкнется от земли.[26]
Патология
Правая нижняя челюсть взрослого человека U.W. 101-1142 имеет костное поражение, указывающее на доброкачественная опухоль. Человек мог бы испытать отек и локальный дискомфорт, но расположение опухоли рядом с медиальная крыловидная мышца могли затруднить функцию мышцы и изменить высоту правой стороны челюсти, а также вызвать дискомфорт справа височно-нижнечелюстной сустав (соединение челюсти с черепом).[27]
Стоматологические дефекты в Х. наледи особи в течение 1,6–2,8 и 4,3–7,6 мес развития, скорее всего, были вызваны сезонными стрессорами. Это могло произойти из-за экстремальных летних и зимних температур, вызывающих нехватку продуктов питания. Кроме того, минимальная зимняя температура в этом районе составляет в среднем около 3 ° C (37 ° F) и может опускаться ниже нуля; сохранение тепла для младенца с маленьким телом Х. наледи было бы трудно, а зимы, вероятно, увеличили восприимчивость к респираторным заболеваниям. Экологические стрессоры соответствуют современным сезоны гриппа в Южной Африке достигает пика зимой, а педиатрические понос госпитализация чаще всего бывает летом в разгар сезона дождей.[28]
Местные гоминины, вероятно, были жертвами крупных хищников, таких как львы, леопарды и гиены. Тем не менее, кажется, наблюдается явная нехватка останков крупных хищников с северного конца Колыбели человечества, где находится пещера Восходящей звезды, возможно потому, что хищники предпочитали Река Блаубанк к югу, который, возможно, предлагал лучшие охотничьи угодья с большим количеством крупных объектов добычи. С другой стороны, поскольку на юге известно гораздо больше стоянок, чем на севере, пространственные структуры хищников могут не быть хорошо представлены в летописи окаменелостей.[29]
Культура
Еда
Сколы на зубах и их ношение указывают на привычное употребление в пищу мелких твердых предметов, таких как грязь и пыль, а чашеобразный износ задних зубов может быть результатом песчаной пищи. Такие частицы могли образоваться из немытых корней и клубней. С другой стороны, засушливость могла привести к попаданию твердых частиц на продукты питания, покрывая их пыль. Возможно, они обычно ели более крупные твердые продукты, такие как семена и орехи, но перед употреблением они были переработаны на более мелкие кусочки.[30][31] В любом случае, Х. наледи занял, казалось бы, уникальный ниша от предыдущих южноафриканских гомининов, в том числе Австралопитек и Парантроп. Тем не менее, зубы всех трех видов указывают на то, что им нужно было приложить большое усилие сдвига, чтобы прожевать, возможно, растительные или мышечные волокна. Зубы других Гомо не может создавать такие высокие силы, возможно, из-за использования некоторых методов обработки пищевых продуктов, таких как приготовление пищи.[30]
Технологии
Хотя Х. наледи останки не связаны с какими-либо каменными орудиями, вероятно, они произведены Ранний каменный век (Ашельский и, возможно, более ранний Oldawan ) или же Средний каменный век отрасли потому что они имеют те же приспособления к руке, что и другие человеческие виды, которые участвуют в производстве инструментов.[9][14] Х. наледи - единственный идентифицированный вид человека, существовавший в раннем среднем каменном веке Highveld регион, Южная Африка, что, возможно, указывает на то, что этот вид создал и сохранил эту традицию, по крайней мере, в этот период времени. Такие отрасли и методы обработки камня, вероятно, развивались независимо несколько раз среди разных Гомо виды и популяции, или были перенесены на большие расстояния изобретателями или учениками и обучены.[9]
Похороны

В 2015 году археолог Пол Диркс, Бергер и его коллеги пришли к выводу, что тела должны были быть намеренно перенесены и помещены в камеру людьми, потому что они выглядели неповрежденными, когда были впервые помещены в камеру (нет доказательств травмы из-за того, что они были помещены в камеру). попадание в камеру ни хищников, ни исключительной сохранности), камера недоступна для крупных хищников, камера кажется изолированной системой и никогда не была затоплена (то есть, естественные силы не участвовали в игре), нет никаких скрытых шахта, через которую могли случайно попасть люди, и нет никаких свидетельств какой-либо катастрофы, в результате которой погибли все люди внутри камеры. Они сказали, что также возможно, что тела сбрасывались по желобу и падали медленно из-за неровностей и узости пути вниз или из-за мягкой грязевой подушки, на которую они приземлялись. В обоих сценариях гробовщикам потребовалось бы искусственное освещение для навигации по пещере; и это место неоднократно использовалось для захоронений, поскольку не все тела были захоронены одновременно.[3]
В 2016 году палеоантрополог Аврора Вал возражала, что такое сохранение могло быть связано с мумификация нежели тщательное захоронение, и отсутствие длинная кость Головы напоминает хищничество, и она считает, что игнорирование природных сил, таких как наводнение, для захоронения тел неоправданно. Кроме того, есть доказательства повреждений, нанесенных жуками, личинками жуков и улитками (которые способствуют разложению); но камера не обеспечивает идеальных условий для улиток и не содержит раковин улиток, что указывало бы на то, что разложение действительно началось до осаждения в камере.[32]
Тем не менее, в 2017 году Диркс, Бергер и его коллеги подтвердили, что нет никаких доказательств того, что вода течет в пещеру, и что более вероятно, что эти Х. наледи были похоронены в камере. Они также сказали, что, возможно, их похоронили современные люди. Гомо, например, предки современных людей, а не другие Х. наледи, но, тем не менее, культурное поведение погребальных обрядов не является невозможным для Х. наледи, и захоронение в камере могло быть сделано для удаления разлагающихся тел из поселения, предотвращения падальщиков или из-за социальных связей и горя.[9]
В 2018 году антрополог Чарльз Эгеланн и его коллеги поддержали мнение Вэла и заявили, что недостаточно доказательств, чтобы сделать вывод о том, что человеческий вид разработал концепцию загробная жизнь так рано. Они также сказали, что сохранение особей Dinaledi аналогично сохранению трупов павианов, которые накапливаются в пещерах (либо естественной смертью обитающих в пещерах павианов, либо леопарда, таскающего туши).[33]
Галерея
- Скелетные элементы

Эндокраниум цифровая реконструкция

Зажимы LES1 (слева) и DH1 (справа)

Верхняя челюсть LES1 (слева) и DH1 (справа)
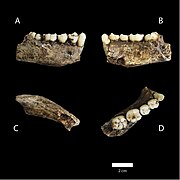
Кость нижней челюсти

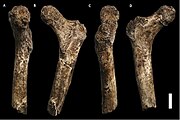
А ключица
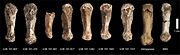
Пястные кости из разных образцов

10-е грудной позвонок

11-е грудной позвонок

А и Б) подвздошная кость, C и D) подростковый крестец, E и F) седалищная кость

Кости лодыжки из разных образцов

1) правая ступня взрослого человека, 2) ювенильный левый, 3 и 4) левый взрослый, 5) ювенильный правый














Смотрите также
- Африканская археология
- Австралопитек седиба - Гоминин возрастом два миллиона лет из "Колыбели человечества"
- Денисовский - Азиатский архаичный человек
- Homo luzonensis - Архаичный человек из Лусона, Филиппины
- Homo floresiensis - Архаичный человек из Флореса, Индонезия
- Неандерталец - Евразийский вид или подвид архаичного человека
- Люди из пещеры благородного оленя - Архаические люди из 12500 г. до н.э. на юго-западе Китая
- Хронология эволюции человека
Рекомендации
- ^ а б Диркс, П. Х. Г. М .; Робертс, Э. М .; Hilbert-Wolf, H .; Kramers, J.D .; Hawks, J .; и другие. (2017). "Возраст Homo naledi и сопутствующие отложения в пещере Восходящей звезды, Южная Африка ". eLife. 6: e24231. Дои:10.7554 / eLife.24231. ЧВК 5423772. PMID 28483040.
- ^ а б Хоукс, Дж. (2016). "Последние новости Homo naledi". Американский ученый. 104 (4): 198. Дои:10.1511/2016.121.198. Архивировано из оригинал 16 мая 2017 г.
- ^ а б c Диркс, П. Х. Г. М .; Бергер, Л.; Робертс, Э. М .; и другие. (2015). "Геологический и тафономический контекст для нового вида гомининов Homo naledi из камеры Диналеди, Южная Африка ". eLife. 4: e09561. Дои:10.7554 / eLife.09561. ISSN 2050-084X. ЧВК 4559842. PMID 26354289.
- ^ Bolter, D. R .; Хоукс, Дж.; Богин, Б .; Кэмерон, Н. (2018). «Палеодемография людей в камере Диналеди, использующих останки зубов». Южноафриканский научный журнал. Претория. 114 (1/2). Дои:10.17159 / sajs.2018 / 20170066.
- ^ а б c d е ж грамм час Бергер, Л.; и другие. (2015). "Homo naledi, новый вид рода Гомо из камеры Диналеди, Южная Африка ". eLife. 4. Дои:10.7554 / eLife.09560. ЧВК 4559886. PMID 26354291.
- ^ а б Хоукс, Дж. Д.; Elliott, M .; Schmid, P .; Черчилль, С. Э .; de Ruiter, D. J .; Робертс, Э. М. (2017). "Новые ископаемые останки Homo naledi из Палаты Леседи, Южная Африка ". eLife. 6: e24232. Дои:10.7554 / eLife.24232. ЧВК 5423776. PMID 28483039.
- ^ Дембо, М .; Радовчич, Д .; Garvin, H.M .; Laird, M. F .; Schroeder, L .; Scott, J. E .; Brophy, J .; Ackermann, R.R .; Мусиба, Ч. М. (2016). "Эволюционные отношения и возраст Homo naledi: Оценка с использованием устаревших байесовских филогенетических методов ». Журнал эволюции человека. 97: 17–26. Дои:10.1016 / j.jhevol.2016.04.008. HDL:2164/8796. PMID 27457542.
- ^ Теккерей, Дж. Ф. (2015). "Оценка возраста и родства Homo naledi". Южноафриканский научный журнал. 111 (11/12). Дои:10.17159 / sajs.2015 / a0124. ISSN 1996-7489.
- ^ а б c d е ж Бергер, Л.; Хоукс, Дж.; Диркс, П. Х. Г. М .; Elliott, M .; Робертс, Э. М. (2017). "Homo naledi и эволюция плейстоценовых гомининов в субэкваториальной Африке ». eLife. 6. Дои:10.7554 / eLife.24234. ЧВК 5423770. PMID 28483041.
- ^ Сутикна, Т .; Tocheri, M. W .; Морвуд, М. Дж .; Saptomo, E.W .; Ятмико; Awe, R.D .; Wasisto, S .; Westaway, K. E .; Обер, М .; Li, B .; Zhao, J .; Этажная, М .; Alloway, B.V .; Морли, М. У .; Meijer, H. J. M .; van den Bergh, G.D .; Grün, R .; Dosseto, A .; Brumm, A .; Jungers, W. L .; Робертс, Р. Г. (2016). "Пересмотренная стратиграфия и хронология для Homo floresiensis в Лянг Буа в Индонезии ". Природа. 532 (7599): 366–369. Bibcode:2016Натура.532..366S. Дои:10.1038 / природа17179. PMID 27027286. S2CID 4469009.
- ^ а б Schroeder, L .; Scott, J. E .; Garvin, H.M .; Laird, M. F .; и другие. (2017). "Разнообразие черепов в Гомо происхождение и относительное положение Homo naledi". Журнал эволюции человека. 104: 124–135. Дои:10.1016 / j.jhevol.2016.09.014. PMID 27836166.
- ^ Стрингер, К. (2015). "Множество загадок Homo naledi". eLife. 4: e10627. Дои:10.7554 / eLife.10627. ISSN 2050-084X. ЧВК 4559885. PMID 26354290.
- ^ Allen, J. S .; Damasio, H .; Грабовски, Т. J. (2002). «Нормальные нейроанатомические изменения в человеческом мозге: объемное исследование МРТ». Американский журнал физической антропологии. 118 (4): 341–358. Дои:10.1002 / ajpa.10092. PMID 12124914. S2CID 21705705.
- ^ а б c Garvin, H.M .; Elliot, M.C .; Делезене, Л. К. (2017). "Размер тела, размер мозга и половой диморфизм у Homo naledi из палаты Диналеди ". Журнал эволюции человека. 111: 119–138. Дои:10.1016 / j.jhevol.2017.06.010. PMID 28874266.
- ^ а б Hollowaya, R.L .; Hurstb, S.D .; Garvin, H.M .; Schoenemann, P.T .; Vanti, W. B .; Бергер, Л.; Хоукс, Дж. (2018). "Морфология эндокаста Homo naledi из камеры Диналеди, Южная Африка ". Труды Национальной академии наук. 115 (22): 5738–5743. Дои:10.1073 / pnas.1720842115. ЧВК 5984505. PMID 29760068.
- ^ Кофран, Чжунтао; Скиннер, М. М .; Уокер, К.С. (2016). «Стоматологическое развитие и история жизни в Homo naledi". Американский журнал физической антропологии. 159: 3–346. Дои:10.1098 / rsbl.2017.0339. ЧВК 5582112. PMID 26914367.
- ^ Gautelli-Steinberg, D .; О'Хара, М. С .; Le Cabec, A .; и другие. (2018). «Паттерны латерального роста эмали в Homo naledi согласно оценке по распределению и количеству перикимат " (PDF). Журнал эволюции человека. 121: 40–54. Дои:10.1016 / j.jhevol.2018.03.007. PMID 29709292.
- ^ Kupczik, K .; Делезене, Л. К .; Скиннер, М. М. (2019). «Морфология корня моляра нижней челюсти и полости пульпы в Homo naledi и другие гоминины плио-плейстоцена » (PDF). Журнал эволюции человека. 130: 83–95. Дои:10.1016 / j.jhevol.2019.03.007. PMID 31010546.
- ^ а б Irish, J. D .; Bailey, S.E .; Guatelli-Steinberg, D .; Делезене, Л. К .; Бергер, Л. (2018). "Древние зубы, фенетическая близость и африканские гоминины: еще один взгляд на то, где Homo naledi вписывается в " (PDF). Журнал эволюции человека. 122: 108–123. Дои:10.1016 / j.jhevol.2018.05.007. PMID 29887210.
- ^ Elliott, M.C .; Quam, R .; Nalla, S .; de Ruiter, D. J .; Хоукс, Дж. Д.; Бергер, Л. (2018). "Описание и анализ трех Homo naledi входит из камеры Диналеди, пещера Восходящей звезды (ЮАР) ». Журнал эволюции человека. 122: 146–155. Дои:10.1016 / j.jhevol.2018.06.008. PMID 30001870.
- ^ Bolter, D. R .; Elliot, M.C .; Хок, Дж. Д.; Бергер, Л. (2020). "Незрелые останки и первый частичный скелет молоди Homo naledi, гоминин позднего среднего плейстоцена из Южной Африки ». PLOS ONE. 15 (4): e0230440. Дои:10.1371 / journal.pone.0230440. ЧВК 7112188. PMID 32236122.
- ^ а б c Уильямс, С. А .; Гарсиа-Мартинес, Д .; и другие. (2017). "Позвонки и ребра Homo naledi". Журнал эволюции человека. 104: 136–154. Дои:10.1016 / j.jhevol.2016.11.003. PMID 28094004.
- ^ VanSickle, C .; Cofran, Z .; Гарсиа-Мартинес, Д .; и другие. (2018). "Homo naledi останки таза из камеры Диналеди, Южная Африка ". Журнал эволюции человека. 125: 122–136. Дои:10.1016 / j.jhevol.2017.10.001. PMID 29169681.
- ^ а б Марчи, Д .; Уокер, С. С .; Wei, P .; и другие. (2017). "Бедро и нога Homo naledi". Журнал эволюции человека. 104: 174–204. Дои:10.1016 / j.jhevol.2016.09.005. PMID 27855981.
- ^ а б Kivell, Tracy L .; Дин, Эндрю С .; Точери, Мэтью В .; Орр, Кейли М .; Шмид, Питер; Ястребы, Джон; Berger, Lee R .; Черчилль, Стивен Э. (2015). "Рука Homo naledi". Nature Communications. 6: 8431. Bibcode:2015НатКо ... 6,84 31 тыс.. Дои:10.1038 / ncomms9431. ЧВК 4597335. PMID 26441219.
- ^ Harcourt-Smith, W.E.H .; Throckmorton, Z .; Congdon, K. A .; Zipfel, B .; Deane, A. S .; Drapeau, M. S. M .; Черчилль, С. Э .; Berger, L.R .; Де Сильва, Дж. М. (2015). "Подножие Homo naledi". Nature Communications. 6: 8432. Bibcode:2015 НатКо ... 6,84 32 ч. Дои:10.1038 / ncomms9432. ЧВК 4600720. PMID 26439101.
- ^ Odes, E.J .; Делезене, Л. К .; и другие. (2018). «Случай доброкачественной остеогенной опухоли в г. Homo naledi: Доказательства периферической остеомы в U.W. 101-1142 нижняя челюсть ». Международный журнал палеопатологии. 21: 47–55. Дои:10.1016 / j.ijpp.2017.05.003. PMID 29778414.
- ^ Скиннер, М. Ф. (2019). «Стресс развития у южноафриканских гомининов: Сравнение рецидивирующих гипоплазий эмали у Австралопитек африканский и Homo naledi". Южноафриканский научный журнал. 115 (5–6). Дои:10.17159 / sajs.2019 / 5872.
- ^ Рейнольдс, С. С. (2010). «Где были дикие животные: пространственное и временное распределение хищников в колыбели человечества (Гаутенг, Южная Африка) в связи с скоплением скоплений млекопитающих и гомининов». Журнал тафономии. 8 (2–3): 233–257.
- ^ а б Berthaume, M.A .; Делезене, Л. К .; Купчик, К. (2018). «Топография зубов и диета Homo naledi" (PDF). Журнал эволюции человека. 118: 14–26. Дои:10.1016 / j.jhevol.2018.02.006. PMID 29606200.
- ^ Towle, I .; Irish, J. D .; де Гроот, И. (2017). "Поведенческие выводы из высокого уровня зубного скола в Homo naledi" (PDF). Американский журнал физической антропологии. 164 (1): 184–192. Дои:10.1002 / ajpa.23250. ISSN 1096-8644. PMID 28542710.
- ^ Вал, А. (2016). «Преднамеренное уничтожение тела гомининами в камере Диналеди, Колыбель человечества, Южная Африка?». Журнал эволюции человека. 96: 145–148. Дои:10.1016 / j.jhevol.2016.02.004. PMID 27039664.
- ^ Egeland, C.P .; Домингес-Родриго, М .; Пикеринг, Т. Р .; и другие. (2018). "Обилие скелетных частей гомининов и заявления о преднамеренном избавлении от трупов в среднем плейстоцене". Труды Национальной академии наук. 115 (18): 4601–4606. Дои:10.1073 / pnas.1718678115. ЧВК 5939076. PMID 29610322.
дальнейшее чтение
- Бергер, Л.; Хоукс, Дж. Д. (2017). Почти человек: удивительная история о Homo naledi и открытии, изменившем нашу человеческую историю. Вашингтон, округ Колумбия: Национальное географическое общество. ISBN 978-1-4262-1811-8.
внешняя ссылка
- Реконструкции Х. наледи автор: palaeoartist Джон Гурч
- Уиллер, Шэрон. "Депеши одной из восходящих звезд пещеры". Тьма внизу.
- «Выдающиеся окаменелости гоминидов». Talk Origins.
- «Изучение летописи окаменелостей гоминидов». Фонд Брэдшоу.
- "блог участников Rising Star Expedition". Национальная география. Архивировано из оригинал 9 сентября 2015 г.
- "Трехмерное сканирование Homo naledi окаменелости ". MorphoSource.
- «Хронология человека (интерактивная)». Национальный музей естественной истории. Смитсоновский институт.
| Таксономия (Гоминины ) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Предки |
| ||||||||||||||||||||||||||
| Модели |
| ||||||||||||||||||||||||||
| Сроки | |||||||||||||||||||||||||||
| Другие |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Места |  | ||||
|---|---|---|---|---|---|
| Окаменелости | |||||
| Антропология |
| ||||