Тейкопланин - Teicoplanin
 | |
| Клинические данные | |
|---|---|
| Торговые наименования | Таргоцид |
| AHFS /Drugs.com | Международные названия лекарств |
| Беременность категория |
|
| Маршруты администрация | Внутривенно, внутримышечно |
| Код УВД | |
| Легальное положение | |
| Легальное положение | |
| Фармакокинетический данные | |
| Биодоступность | 90% (учитывая Я ) |
| Связывание с белками | От 90% до 95% |
| Метаболизм | Ноль |
| Устранение период полураспада | От 70 до 100 часов |
| Экскреция | Почечный (97% без изменений) |
| Идентификаторы | |
| |
| Количество CAS | |
| PubChem CID | |
| DrugBank | |
| ChemSpider | |
| UNII | |
| КЕГГ | |
| ЧЭМБЛ | |
| NIAID ChemDB | |
| Панель управления CompTox (EPA) | |
| Химические и физические данные | |
| Формула | Переменная |
| Молярная масса | От 1564,3 до 1907,7 г / моль |
| |
| | |
Тейкопланин является антибиотик используется в профилактика и лечение серьезных инфекций, вызванных Грамположительный бактерии, включая метициллин-устойчивый Золотистый стафилококк и Enterococcus faecalis. Это полусинтетический гликопептидный антибиотик со спектром активности, аналогичным ванкомицин. Его механизм действия заключается в подавлении синтеза клеточной стенки бактерий.[1]
Тейкопланин продается Санофи-Авентис под торговой маркой Таргоцид. Другие торговые наименования включают Тикоцин продается компанией Cipla (Индия).
Доказано, что пероральный тейкопланин эффективен при лечении: псевдомембранозный колит и Clostridium difficile -ассоциированная диарея, сравнимая по эффективности с ванкомицином.[2]
Считается, что его сила обусловлена длиной углеводородной цепи.[3]
Данные о восприимчивости
Тейкопланин нацелен на синтез пептидогликана, что делает его эффективным противомикробным средством против грамположительных бактерий, включая Стафилококки и Clostridium виды Ниже представлены данные о чувствительности к МПК для нескольких значимых с медицинской точки зрения патогенов:
- Clostridium difficile: 0,06 мкг / мл - 0,5 мкг / мл
- Золотистый стафилококк: ≤0,06 мкг / мл - ≥128 мкг / мл
- Эпидермальный стафилококк: ≤0,06 мкг / мл - 32 мкг / мл
Химия
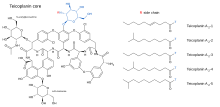
Тейкопланин (TARGOCID, продаваемый Sanofi Aventis Ltd) на самом деле представляет собой смесь нескольких соединений, пяти основных (названных тейкопланин А2-1 через А2-5) и четыре второстепенных (названы тейкопланин RS-1 через рS-4).[5]Все тейкопланины имеют одно и то же гликопептид ядро, называемое тейкопланин А3-1 - а сплавленное кольцо структура, к которой два углеводы (манноза и N-ацетилглюкозамин ) прилагаются. Основные и второстепенные компоненты также содержат третий углевод. часть — β-D-глюкозамин - и отличаются только длиной и строением боковая цепь прикреплен к нему.
Структуры ядра тейкопланина и боковых цепей, которые характеризуют пять основных соединений тейкопланина, показаны ниже.

Биосинтез
Тейкопланин относится к комплексу родственных природных продуктов, выделенных из ферментационного бульона штамма Actinoplanes teichomyceticus,[6] состоящий из группы из пяти структур. Эти структуры имеют общий агликон, или ядро, состоящее из семи аминокислот, связанных пептидными и эфирными связями с образованием четырехкольцевой системы. Эти пять структур различаются идентичностью жирной ацильной боковой цепи, присоединенной к сахару. Происхождение этих семи аминокислот в биосинтезе тейкопланина было изучено 1Рука 13C. Ядерный магнитный резонанс.[7] Исследования указывают на аминокислоты 4-л.с., 3-Cl-Tyr и 3-хлор-β-гидрокситирозин являются производными тирозина, а аминокислота 3,5-дигидроксифенилглицин (3,5-Dpg) происходит из ацетата. Тейкопланин содержит 6 непротеиногенных аминокислот и три сахарных фрагмента, N-ацил-β-D-глюкозамин, N-ацетил-β-D-глюкозамин и D-манноза.
Генный кластер
Изучение генетического кластера, кодирующего биосинтез тейкопланина, выявило 49 предполагаемых открытых рамок считывания (ORF), участвующих в биосинтезе, экспорте, устойчивости и регуляции соединения. Тридцать пять из этих открытых рамок считывания сходны с открытыми в других кластерах гликопептидных генов. Функция каждого из этих генов описана Ли с сотрудниками.[8] Краткое описание структуры и назначения гена показано ниже.
Макет гена. Гены пронумерованы. Буквы L и R обозначают направление транскрипции. Наличие символа * означает, что ген обнаружен после NRP, которые представлены буквами A, B, C и D. На основании рисунка из: Li, T-L .; Хуанг, Ф .; Haydock, S. F .; Мироненко, Т .; Leadlay, P. F .; Спенсер, Дж. Б. Химия и биология. 2004, 11, с. 109.
[11-левый] [10-левый] [9-левый] [8-левый] [7-левый] [6-левый] [5-левый] [4-левый] [3-левый] [2-левый] [1-R] [AR] [BR] [CR] [DR] [1 * -R] [2 * -R] [3 * -R] [4 * -R] [5 * -R] [6 * -R] [7 * -R] [8 * -R] [9 * -R] [10 * -R] [11 * -R] [12 * -R] [13 * -R] [14 * -R ] [15 * -R] [16 * -R] [17 * -R] [18 * -R] [19 * -R] [20 * -R] [21 * -R] [22 * -R] [ 23 * -R] [24 * -R] [25 * -L] [26 * -L] [27 * -R] [28 * -R] [29 * -R] [30 * -R] [31 * -R] [32 * -L] [33 * -L] [34 * -R]
| Фермент, продуцируемый последовательностью гена | Регуляторные белки | Другие ферменты | Устойчивые ферменты | Ферменты биосинтеза Β-гидрокситирозина и 4-гидроксифенилглицина | Гликозилтрансферазы | Пептид синтетазы | P450 оксигеназы | Галогеназа | Ферменты биосинтеза 3,5-дигидроксифенилглицина |
| Гены | 11, 10, 3, 2, 15*, 16*, 31* | 9, 8, 1*, 2*, 4*, 11*, 13*, 21*, 26*, 27*, 30*, 32*, 33*, 34* | 7, 6, 5 | 4, 12*, 14*, 22*, 23*, 24*, 25*, 28*, 29* | 1, 3*, 10* | А, Б, В, D | 5*, 6*, 7*, 9* | 8* | 17*, 18*, 19*, 20*, 23* |
Синтез гептапептидного остова
Гептапептидный каркас тейкопланина собирается с помощью нерибосомных пептидных синтетаз (NRPS) TeiA, TeiB, TeiC и TeiD. Вместе они составляют семь модулей, каждый из которых содержит несколько доменов, каждый из которых отвечает за включение одной аминокислоты. Модули 1, 4 и 5 активируют L-4-Hpg как аминоацил-AMP, модули 2 и 6 активируют L-Tyr, а модули 3 и 7 активируют L-3,5-Dpg. Активированные аминокислоты ковалентно связаны с NRPS в виде тиоэфиров с помощью кофактора фосфопантетеина, который присоединен к домену белка-носителя пептидила (PCP). Связанные с ферментом аминокислоты затем соединяются амидными связями под действием домена конденсации (C).
Гептапетид тейкопланина содержит 4 D-аминокислоты, образованные эпимеризацией активированных L-аминокислот. Каждый из модулей 2, 4 и 5 содержит домен эпимеризации (E), который катализирует это изменение. Модуль 1 не содержит домена E, и предполагается, что эпимеризация будет катализироваться доменом C.[9] В целом, шесть из семи общих аминокислот основной цепи тейкопланина состоят из небелковых или модифицированных аминокислот. Одиннадцать ферментов согласованно индуцируются для образования этих шести необходимых остатков.[10] Тейкопланин содержит два хлорированных положения: 2 (3-Cl-Tyr) и 6 (3-Cl-β-Hty). Галогеназа Tei8 * действует, катализируя галогенирование обоих остатков тирозина. Хлорирование происходит на уровне аминоацил-PCP во время биосинтеза, до фенольного окислительного связывания, при этом тирозин или β-гидрокситирозин могут быть субстратами хлорирования.[11] Также происходит гидроксилирование остатка тирозина модуля 6. в транс во время сборки гептапептидного остова.
Модификация после образования гептапептидного остова
Как только гептапептидный остов сформирован, линейный промежуточный продукт, связанный с ферментом, подвергается циклизации.[10] Исследования разрушения генов показывают, что оксигеназы цитохрома P450 являются ферментами, выполняющими реакции связывания. X-домен в конечном модуле NRPS необходим для рекрутирования ферментов оксигеназы.[12] OxyB образует первое кольцо путем связывания остатков 4 и 6, а OxyE затем соединяет остатки 1 и 3. OxyA соединяет остатки 2 и 4 с последующим образованием связи C-C между остатками 5 и 7 посредством OxyC.[13] Региоселективность и атропоизомер Было высказано предположение, что селективность этих вероятных реакций одноэлектронного связывания обусловлена требованиями к укладке и ориентации частично перекрестно-связанных субстратов в активном центре фермента.[10] Реакции сочетания показаны ниже.

Было показано, что специфическое гликозилирование происходит после образования гептпептидного агликона.[14] Для гликозилирования агликона тейкопланина необходимы три отдельных гликозилтрансферазы. Tei10 * катализирует добавление GlcNAc к остатку 4 с последующим деацетилированием с помощью Tei2 *. Затем к ацильной цепи (образованной действием Tei30 * и Tei13 *) добавляется Tei11 *. Затем Tei1 добавляет второй GlcNAc к β-гидроксильной группе остатка 6 с последующим маннозилированием остатка 7, катализируемым Tei3 *.[15]
Рекомендации
- ^ Reynolds PE (ноябрь 1989 г.). «Структура, биохимия и механизм действия гликопептидных антибиотиков». Европейский журнал клинической микробиологии и инфекционных заболеваний. 8 (11): 943–50. Дои:10.1007 / BF01967563. PMID 2532132. S2CID 21551939.
- ^ de Lalla F, Nicolin R, Rinaldi E, Scarpellini P, Rigoli R, Manfrin V, Tramarin A (октябрь 1992 г.). «Проспективное исследование перорального тейкопланина по сравнению с пероральным ванкомицином для терапии псевдомембранозного колита и диареи, связанной с Clostridium difficile». Противомикробные препараты и химиотерапия. 36 (10): 2192–6. Дои:10.1128 / AAC.36.10.2192. ЧВК 245474. PMID 1444298.
- ^ Гилпин М., Милнер П. (1997). «Сопротивление изменениям. За последние 40 лет гликопептидные антибиотики сыграли решающую роль в лечении бактериальных инфекций. Но как долго это может продолжаться?». Королевское химическое общество. Архивировано из оригинал 21 декабря 2002 г.. Получено 2006-10-15. - включает изображение структуры Тейкопланина.
- ^ Данные о чувствительности к тейкопланину и минимальной ингибирующей концентрации (МИК)
- ^ Бернареджи А., Борги А., Боргонови М., Кавенаги Л., Феррари П, Веке К. и др. (Август 1992 г.). «Метаболизм тейкопланина у человека». Противомикробные препараты и химиотерапия. 36 (8): 1744–9. Дои:10.1128 / AAC.36.8.1744. ЧВК 192040. PMID 1416858.
- ^ Юнг Х.М., Джея М., Ким С.И., Мун Х.Д., Кумар Сингх Р., Чжан Ю.В., Ли Дж.К. (сентябрь 2009 г.). «Биосинтез, биотехнологическое производство и применение тейкопланина: состояние и перспективы». Прикладная микробиология и биотехнология. 84 (3): 417–28. Дои:10.1007 / s00253-009-2107-4. PMID 19609520. S2CID 45038487.
- ^ Heydorn A, Petersen BO, Duus JO, Bergmann S, Suhr-Jessen T., Nielsen J (март 2000 г.). «Биосинтетические исследования гликопептида тейкопланина с помощью (1) H и (13) C ЯМР». Журнал биологической химии. 275 (9): 6201–6. Дои:10.1074 / jbc.275.9.6201. PMID 10692413.
- ^ Ли Т.Л., Хуанг Ф., Хейдок С.Ф., Мироненко Т., Лидли П.Ф., Спенсер Дж.Б. (январь 2004 г.). «Кластер биосинтетических генов гликопептидного антибиотика тейкопланина: характеристика двух гликозилтрансфераз и ключевой ацилтрансферазы». Химия и биология. 11 (1): 107–19. Дои:10.1016 / j.chembiol.2004.01.001. PMID 15113000.
- ^ Kaniusaite M, Tailhades J, Kittilä T, Fage CD, Goode RJ, Schittenhelm RB, Cryle MJ (май 2020 г.). «Понимание ранних стадий образования пептидов во время биосинтеза тейкопланина и родственных гликопептидных антибиотиков». Журнал FEBS: febs.15350. Дои:10.1111 / фев.15350. PMID 32359003.
- ^ а б c Кане Д., Леймкулер С., Лу В., Уолш С. (февраль 2005 г.). «Гликопептидные и липогликопептидные антибиотики». Химические обзоры. 105 (2): 425–48. Дои:10.1021 / cr030103a. PMID 15700951.
- ^ Киттиля Т., Киттель С., Тейлхэйдс Дж., Бутц Д., Шоппет М., Бюттнер А. и др. (Сентябрь 2017 г.). «Галогенирование гликопептидных антибиотиков происходит на аминокислотном уровне при синтезе нерибосомных пептидов». Химическая наука. 8 (9): 5992–6004. Дои:10.1039 / C7SC00460E. ЧВК 5620994. PMID 28989629.
- ^ Haslinger K, Peschke M, Brieke C, Maximowitsch E, Cryle MJ (май 2015 г.). «X-домен пептидных синтетаз привлекает оксигеназы, важные для биосинтеза гликопептидов». Природа. 521 (7550): 105–9. Bibcode:2015Натура.521..105H. Дои:10.1038 / природа14141. PMID 25686610. S2CID 4466657.
- ^ Пешке М., Брике С., Крайл М.Дж. (октябрь 2016 г.). «Формирование кольца F-O-G в биосинтезе гликопептидных антибиотиков катализируется OxyE». Научные отчеты. 6 (1): 35584. Bibcode:2016НатСР ... 635584П. Дои:10.1038 / srep35584. ЧВК 5067714. PMID 27752135.
- ^ Каплан Дж, Корти Б.Д., Аксельсен П.Х., Лолл П.Дж. (май 2001 г.). «Роль сахарных остатков в молекулярном распознавании ванкомицином». Журнал медицинской химии. 44 (11): 1837–40. Дои:10.1021 / jm0005306. PMID 11356118.
- ^ Ющук О., Осташ Б., Фам Т.Х., Лужецкий А., Федоренко В., Труман А.В., Горбаль Л. (август 2016 г.). "Характеристика пост-сборочных процессов пошива в биосинтезе тейкопланина". ACS Химическая биология. 11 (8): 2254–64. Дои:10.1021 / acschembio.6b00018. PMID 27285718.