Hsp70 - Hsp70
| Белок Hsp70 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
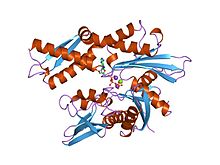 Структура фрагмента АТФазы 70K родственного белка теплового шока.[1] | |||||||||
| Идентификаторы | |||||||||
| Символ | HSP70 | ||||||||
| Pfam | PF00012 | ||||||||
| Pfam клан | CL0108 | ||||||||
| ИнтерПро | IPR013126 | ||||||||
| PROSITE | PDOC00269 | ||||||||
| SCOP2 | 3hsc / Объем / СУПФАМ | ||||||||
| |||||||||
В 70 килодальтон белков теплового шока (Hsp70s или DnaK) представляют собой семейство консервативных повсеместно экспрессируемых белки теплового шока. Белки с похожей структурой существуют практически во всех живых организмах. Hsp70 являются важной частью клеточного механизма сворачивания белков и помогают защитить клетки от стресса.[2][3]
Открытие
Члены семейства Hsp70 очень сильно активируются тепловым стрессом и токсичный химические вещества, особенно тяжелые металлы, такие как мышьяк, кадмий, медь, ртуть и т. д. Hsp70 был впервые обнаружен Ф. М. Ритосса в 1960-х годах, когда сотрудник лаборатории случайно повысил температуру инкубации дрозофилы (плодовых мух). Изучая хромосомы, Ритосса обнаружил «паттерн вздутия», который указывал на повышенную транскрипцию гена неизвестного белка.[4][5] Позже это было описано как «реакция теплового шока», а белки были названы «белками теплового шока» (Hsps).
Структура

Белки Hsp70 выполняют три основных функциональных домены:
- N-концевой АТФаза домен - связывает АТФ (Аденозинтрифосфат ) и гидролизует его до АДФ (Аденозиндифосфат ). NBD состоит из двух долей с глубокой щелью между ними, на дне которой связывается нуклеотид (АТФ и АДФ). Обмен АТФ и АДФ приводит к конформационным изменениям в двух других доменах.
- Домен связывания субстрата - состоит из субдомена β-листа 15 кДа и спирального субдомена 10 кДа. Поддомен β-листов состоит из скрученных β-листов с выступающими вверх петлями, как типичный β баррель, которые охватывают пептидный остов субстрата. SBD содержит бороздку со сродством к нейтральным, гидрофобным аминокислота остатки. Канавка достаточно длинная, чтобы взаимодействовать с пептидами длиной до семи остатков.
- C-терминал домен - богат в альфа-спиральная структура действует как «крышка» для домена связывания субстрата. Спиральный субдомен состоит из пяти спиралей, причем две спирали упакованы с двух сторон субдомена β-листа, стабилизируя внутреннюю структуру. Кроме того, одна из спиралей образует солевой мостик и несколько водородных связей с внешними петлями, тем самым закрывая карман для связывания субстрата как крышку. Три спирали в этом домене образуют еще одно гидрофобное ядро, которое может быть стабилизацией «крышки». Когда белок Hsp70 связан с АТФ, крышка открыта, и пептиды связываются и высвобождаются относительно быстро. Когда белки Hsp70 связаны с АДФ, крышка закрывается, и пептиды прочно связываются с доменом связывания субстрата.[7]
Функция и регулирование
Система Hsp70 взаимодействует с протяженными пептидными сегментами белков, а также с частично свернутыми белками, вызывая агрегацию белков в ключевых путях для дерегуляции активности.[8][9]Когда Hsp70 не взаимодействует с пептидом-субстратом, он обычно находится в связанном с АТФ состоянии. Сам по себе Hsp70 характеризуется очень слабой активностью АТФазы, так что спонтанный гидролиз не происходит в течение многих минут. Поскольку вновь синтезированные белки выходят из рибосомы, субстрат-связывающий домен Hsp70 распознает последовательности гидрофобных аминокислотных остатков и взаимодействует с ними. Это спонтанное взаимодействие обратимо, и в связанном с АТФ состоянии Hsp70 может относительно свободно связываться и высвобождать пептиды. Однако присутствие пептида в связывающем домене стимулирует АТФазную активность Hsp70, увеличивая его обычно медленную скорость гидролиза АТФ. Когда АТФ гидролизуется до АДФ, связывающий карман Hsp70 закрывается, прочно связывая теперь захваченную пептидную цепь. Еще больше ускоряют гидролиз АТФ так называемые коапероны J-домена: Hsp40 у эукариот и DnaJ у прокариот. Эти коапероны резко увеличивают АТФазную активность Hsp70 в присутствии взаимодействующих пептидов.

Путем связывания с частично синтезированными пептидными последовательностями (неполными белками) Hsp70 предотвращает их агрегацию и их нефункциональность. После того, как весь белок синтезирован, фактор обмена нуклеотидов (прокариотический GrpE, эукариотический BAG1 и HspBP1 среди тех, которые были идентифицированы) стимулирует высвобождение АДФ и связывание свежего АТФ, открывая карман связывания. Затем белок может сворачиваться самостоятельно или передаваться другим шаперонам для дальнейшей обработки.[10] ПРЫГАТЬ (в ЧАСsp70 / Hsp90 Оорганизация пrotein) может связываться как с Hsp70, так и с Hsp90 одновременно, и опосредует перенос пептидов от Hsp70 к Hsp90.[11]
Hsp70 также способствует трансмембранному транспорту белков, стабилизируя их в частично свернутом состоянии. Также известно, что он фосфорилируется[12] который регулирует несколько его функций.[13][14][15]
Белки Hsp70 могут защищать клетки от термического или окислительного стресса. Эти стрессы обычно действуют, повреждая белки, вызывая частичное разворачивание и возможную агрегацию. Временно связываясь с гидрофобными остатками, подверженными стрессу, Hsp70 предотвращает агрегацию этих частично денатурированных белков и препятствует их повторной укладке. Низкий уровень АТФ характерен для теплового шока, а устойчивое связывание рассматривается как подавление агрегации, в то время как восстановление после теплового шока включает связывание субстрата и цикл нуклеотидов. У термофильных анаэробов (Thermotoga maritima) Hsp70 демонстрирует окислительно-восстановительное связывание с модельными пептидами, что указывает на второй способ регуляции связывания, основанный на окислительном стрессе.
Hsp70, по-видимому, может участвовать в удалении поврежденных или дефектных белков. Взаимодействие с ЧИП (Cарбоксильный конец ЧАСsp70 явзаимодействие пrotein) –an E3 убиквитинлигаза –Позволяет Hsp70 передавать белки в клетки убиквитинирование и протеолиз пути.[16]
Наконец, помимо улучшения общей целостности белка, Hsp70 непосредственно ингибирует апоптоз.[17] Отличительной чертой апоптоза является высвобождение цитохрома c, который затем рекрутирует Apaf-1 и dATP / ATP в комплекс апоптосом. Затем этот комплекс расщепляет прокаспазу-9, активируя каспазу-9 и в конечном итоге индуцируя апоптоз через активацию каспазы-3. Hsp70 ингибирует этот процесс, блокируя рекрутирование прокаспазы-9 в апоптосомный комплекс Apaf-1 / dATP / цитохром c. Он не связывается напрямую с сайтом связывания прокаспазы-9, но, вероятно, вызывает конформационные изменения, которые делают связывание прокаспазы-9 менее благоприятным. Показано, что Hsp70 взаимодействует с белком-сенсором стресса эндоплазматического ретикулума IRE1alpha, тем самым защищая клетки от апоптоза, вызванного стрессом ER. Это взаимодействие продлевает сплайсинг мРНК XBP-1, тем самым вызывая активацию транскрипции мишеней сплайсированного XBP-1, таких как EDEM1, ERdj4 и P58IPK, спасая клетки от апоптоза.[18] Другие исследования предполагают, что Hsp70 может играть антиапоптотическую роль на других стадиях, но не участвует в апоптозе, опосредованном Fas-лигандом (хотя Hsp 27 участвует). Следовательно, Hsp70 не только сохраняет важные компоненты клетки (белки), но и напрямую спасает клетку в целом. Принимая во внимание, что белки реакции на стресс (такие как Hsp70) эволюционировали до апоптотического аппарата, прямая роль Hsp70 в ингибировании апоптоза дает интересную эволюционную картину того, как более поздний (апоптотический) аппарат приспособился к предыдущему аппарату (Hsps), таким образом выравнивая улучшенную целостность клеточных белков. с повышенными шансами на выживание этой конкретной клетки.
Рак
Hsp70 сверхэкспрессируется при злокачественной меланоме[19] и недоэкспрессируется при почечно-клеточном раке.[20][21]В клеточной линии рака молочной железы (MCF7) было обнаружено, что не только Hsp90 взаимодействует с рецептором эстрогена альфа (ERα), но также Hsp70-1 и Hsc70 также взаимодействуют с ERα.[22]
Выражение в кожной ткани
И HSP70, и HSP47 было показано, что они выражаются в дерма и эпидермис следующий лазерное облучение, а пространственные и временные изменения в паттернах экспрессии HSP определяют зону термического повреждения, вызванную лазером, и процесс заживления в тканях. HSP70 может биохимически определять зону термического повреждения, в которой клетки нацелены на разрушение, а HSP47 может иллюстрировать процесс восстановления после термического повреждения.[23]
Члены семьи
Прокариоты экспрессируют три белка Hsp70: DnaK, HscA (Hsc66), и HscC (Hsc62).[24]
Эукариотические организмы экспрессируют несколько немного разных белков Hsp70. Все они имеют общую доменную структуру, но каждый имеет уникальный паттерн экспрессии или субклеточной локализации. Это, среди прочего:
- Hsc70 (Hsp73 / HSPA8) является конститутивно экспрессируемым белком-шапероном. Обычно он составляет от одного до трех процентов от общего клеточного белка.
- Hsp70 (закодирован тремя очень тесно связанными паралогами: HSPA1A, HSPA1B, и HSPA1L ) является стресс-индуцированным белком. Высокие уровни могут вырабатываться клетками в ответ на гипертермию, окислительный стресс и изменения в организме. pH.
- Связывающий белок иммуноглобулина (BiP или Grp78) представляет собой белок, локализованный в эндоплазматический ретикулум. Он участвует в сворачивании белков и может активироваться в ответ на стресс или голодание.
- mtHsp70 или Grp75 это митохондриальный Hsp70.
Ниже приводится список генов Hsp70 человека и соответствующих им белков:[2]
| ген | белок | синонимы | субклеточное расположение |
|---|---|---|---|
| HSPA1A | Hsp70 | HSP70-1, Hsp72 | Nuc /Цито |
| HSPA1B | Hsp70 | HSP70-2 | Nuc / Cyto |
| HSPA1L | Hsp70 | ? | |
| HSPA2 | Hsp70-2 | ? | |
| HSPA5 | Hsp70-5 | BiP / Grp78 | ER |
| HSPA6 | Hsp70-6 | ? | |
| HSPA7 | Hsp70-7 | ? | |
| HSPA8 | Hsp70-8 | Hsc70 | Nuc / Cyto |
| HSPA9 | Hsp70-9 | Grp75 / mtHsp70 | Мито |
| HSPA12A | Hsp70-12a | ? | |
| HSPA14 | Hsp70-14 | ? |
Hsp110
Суперсемейство Hsp70 также включает семейство Hsp110/Grp170 (Sse) белки, которые представляют собой более крупные белки, связанные с Hsp70.[25] Семейство белков Hsp110 выполняет различные функции: дрожжевой Sse1p имеет небольшую активность АТФазы, но сам по себе является шапероном, а также фактором обмена нуклеотидов для Hsp70, тогда как близкородственный Sse2p имеет небольшую разворачивающую активность.[10]
Ниже приводится список названных в настоящее время генов HSP110 человека. HSPH2-4 - предлагаемые имена, и текущее имя связано:[25]
| ген | синонимы | субклеточное расположение |
|---|---|---|
| HSPH1 | HSP105 | Цито |
| HSPH2 | HSPA4; АПГ-2; HSP110 | Цито |
| HSPH3 | HSPA4L; АПГ-1 | Nuc |
| HSPH4 | HYOU1 / Grp170; ORP150; HSP12A | ER |
Смотрите также
Рекомендации
- ^ Флаэрти К.М., ДеЛука-Флаэрти С., Маккей Д.Б. (август 1990 г.). «Трехмерная структура фрагмента АТФазы родственного белка теплового шока 70K». Природа. 346 (6285): 623–8. Bibcode:1990Натура.346..623F. Дои:10.1038 / 346623a0. PMID 2143562.
- ^ а б Тавария М., Габриэле Т., Кола I, Андерсон Р.Л. (апрель 1996 г.). "Автостопом по человеческому семейству Hsp70". Клеточный стресс и шапероны. 1 (1): 23–8. Дои:10.1379 / 1466-1268 (1996) 001 <0023: AHSGTT> 2.3.CO; 2. ЧВК 313013. PMID 9222585.
- ^ Морано К.А. (октябрь 2007 г.). «Новые приемы для старой собаки: развивающийся мир Hsp70». Летопись Нью-Йоркской академии наук. 1113 (1): 1–14. Bibcode:2007НЯСА1113 .... 1М. Дои:10.1196 / летопись.1391.018. PMID 17513460.
- ^ Ритосса Ф (1962). «Новый паттерн пухлости, вызванный температурным шоком и DNP у дрозофилы». Клеточные и молекулярные науки о жизни. 18 (12): 571–573. Дои:10.1007 / BF02172188.
- ^ Ритосса Ф (июнь 1996 г.). «Открытие реакции на тепловой шок». Клеточный стресс и шапероны. 1 (2): 97–8. Дои:10.1379 / 1466-1268 (1996) 001 <0097: DOTHSR> 2.3.CO; 2. ЧВК 248460. PMID 9222594.
- ^ Vostakolaei MA, Hatami-Baroogh L, Babaei G, Molavi O, Kordi S, Abdolalizadeh J (ноябрь 2020 г.). «Hsp70 при раке: двойной агент в битве между выживанием и смертью». Журнал клеточной физиологии: jcp.30132. Дои:10.1002 / jcp.30132. PMID 33169384.
- ^ MP Mayer (август 2010 г.). «Гимнастика молекулярных шаперонов». Молекулярная клетка. 39 (3): 321–31. Дои:10.1016 / j.molcel.2010.07.012. PMID 20705236.
- ^ Машаги А., Безрукавников С., Минде Д.П., Вентинк А.С., Китик Р., Захманн-Бранд Б., Майер М.П., Крамер Г., Букау Б., Танс С.Дж. (ноябрь 2016 г.). «Альтернативные способы привязки клиента обеспечивают функциональную пластичность Hsp70». Природа. 539 (7629): 448–451. Bibcode:2016Натура.539..448М. Дои:10.1038 / природа20137. PMID 27783598.
- ^ а б Vostakolaei MA, Hatami-Baroogh L, Babaei G, Molavi O, Kordi S, Abdolalizadeh J (ноябрь 2020 г.). «Hsp70 при раке: двойной агент в битве между выживанием и смертью». Журнал клеточной физиологии. Дои:10.1002 / jcp.30132. PMID 33169384.
- ^ а б Брахер А., Вергезе Дж. (2015). "Белки домена GrpE, Hsp110 / Grp170, HspBP1 / Sil1 и BAG: факторы обмена нуклеотидов для молекулярных шаперонов Hsp70". Субклеточная биохимия. Субклеточная биохимия. 78: 1–33. Дои:10.1007/978-3-319-11731-7_1. ISBN 978-3-319-11730-0. PMID 25487014.
- ^ Wegele H, Müller L, Buchner J (2004). Hsp70 и Hsp90 - команда реле для сворачивания белков. Rev. Physiol. Biochem. Pharmacol. Обзоры физиологии, биохимии и фармакологии. 151. С. 1–44. Дои:10.1007 / s10254-003-0021-1. ISBN 978-3-540-22096-1. PMID 14740253.
- ^ Цворо А., Дунджерски Дж., Трайкович Д., Матич Г. (1999-04-01). «Уровень и фосфорилирование Hsp70 в цитозоле печени крысы после адреналэктомии и гипертермии». Cell Biology International. 23 (4): 313–20. Дои:10.1006 / cbir.1998.0247. PMID 10600240.
- ^ Гао Т., Ньютон AC (август 2002 г.). «Мотив поворота представляет собой переключатель фосфорилирования, который регулирует связывание Hsp70 с протеинкиназой C». Журнал биологической химии. 277 (35): 31585–92. Дои:10.1074 / jbc.M204335200. PMID 12080070.
- ^ Труман А.В., Кристьянсдоттир К., Вольфгехер Д., Хасин Н., Польер С., Чжан Х., Перретт С., Продромоу С., Джонс Г.В., Крон С.Дж. (декабрь 2012 г.). «CDK-зависимое фосфорилирование Hsp70 контролирует обилие циклина G1 и прогрессию клеточного цикла». Клетка. 151 (6): 1308–18. Дои:10.1016 / j.cell.2012.10.051. ЧВК 3778871. PMID 23217712.
- ^ Мюллер П., Руккова Е., Халада П., Коутс П.Дж., Хрстка Р., Лейн Д.П., Войтесек Б. (июнь 2013 г.). «С-концевое фосфорилирование Hsp70 и Hsp90 регулирует попеременное связывание с ко-шаперонами CHIP и HOP для определения баланса складывания / деградации клеточного белка». Онкоген. 32 (25): 3101–10. Дои:10.1038 / onc.2012.314. PMID 22824801.
- ^ Людерс Дж., Требование Дж., Хёфельд Дж. (Февраль 2000 г.). «Убиквитин-родственный BAG-1 обеспечивает связь между молекулярными шаперонами Hsc70 / Hsp70 и протеасомой». Журнал биологической химии. 275 (7): 4613–7. Дои:10.1074 / jbc.275.7.4613. PMID 10671488.
- ^ Beere HM, Wolf BB, Cain K, Mosser DD, Mahboubi A, Kuwana T., Tailor P, Morimoto RI, Cohen GM, Green DR (август 2000 г.). «Белок теплового шока 70 ингибирует апоптоз, предотвращая рекрутирование прокаспазы-9 в апоптосому Apaf-1». Природа клеточной биологии. 2 (8): 469–75. Дои:10.1038/35019501. PMID 10934466.
- ^ Гупта С., Дипти А., Диган С., Лиссабона Ф., Хетц С., Самали А. (июль 2010 г.). Келли JW (ред.). «HSP72 защищает клетки от апоптоза, вызванного стрессом ER, посредством усиления передачи сигналов IRE1alpha-XBP1 посредством физического взаимодействия». PLoS Биология. 8 (7): e1000410. Дои:10.1371 / journal.pbio.1000410. ЧВК 2897763. PMID 20625543.
- ^ Ricaniadis N, Kataki A, Agnantis N, Androulakis G, Karakousis CP (февраль 2001 г.). «Долгосрочное прогностическое значение экспрессии HSP-70, c-myc и HLA-DR у пациентов со злокачественной меланомой». Европейский журнал хирургической онкологии. 27 (1): 88–93. Дои:10.1053 / ejso.1999.1018. PMID 11237497.
- ^ Ramp U, Mahotka C, Heikaus S, Shibata T, Grimm MO, Willers R, Gabbert HE (октябрь 2007 г.). «Экспрессия белка теплового шока 70 в почечно-клеточной карциноме и ее связь с прогрессированием опухоли и прогнозом». Гистология и гистопатология. 22 (10): 1099–107. Дои:10.14670 / HH-22.1099. PMID 17616937.
- ^ Шерман М, Multhoff G (октябрь 2007 г.). «Белки теплового шока при раке». Летопись Нью-Йоркской академии наук. 1113 (1): 192–201. Bibcode:2007НЯСА1113..192С. Дои:10.1196 / летопись.1391.030. PMID 17978282.
- ^ Дхамад AE, Zhou Z, Zhou J, Du Y (2016). «Систематическая протеомная идентификация белков теплового шока (Hsp), которые взаимодействуют с альфа-рецептором эстрогена (ERα), и биохимическая характеристика взаимодействия ERα-Hsp70». PLOS One. 11 (8): e0160312. Дои:10.1371 / journal.pone.0160312. PMID 27483141.
- ^ Саджади А.Ю., Митра К., Грейс М. (октябрь 2013 г.). «Экспрессия белков теплового шока 70 и 47 в тканях после короткоимпульсного лазерного излучения: оценка теплового повреждения и заживления» (PDF). Медицинская инженерия и физика. 35 (10): 1406–14. Дои:10.1016 / j.medengphy.2013.03.011. PMID 23587755.
- ^ Ёсимунэ К., Ёсимура Т., Накаяма Т., Нишино Т., Эсаки Н. (май 2002 г.). «Hsc62, Hsc56 и GrpE, третья система шаперонов Hsp70 Escherichia coli». Сообщения о биохимических и биофизических исследованиях. 293 (5): 1389–95. Дои:10.1016 / S0006-291X (02) 00403-5. PMID 12054669.
- ^ а б Кампинга Х. Х., Хагеман Дж., Вос М. Дж., Кубота Х., Тангуай Р. М., Бруфорд Э. А., Читам МЭ, Чен Б., Хайтауэр ЛЭ (январь 2009 г.). «Рекомендации по номенклатуре белков теплового шока человека». Клеточный стресс и шапероны. 14 (1): 105–11. Дои:10.1007 / s12192-008-0068-7. ЧВК 2673902. PMID 18663603.
внешняя ссылка
- HSP70 + тепловой шок + белки в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)