Клеточная мембрана - Википедия - Cell membrane
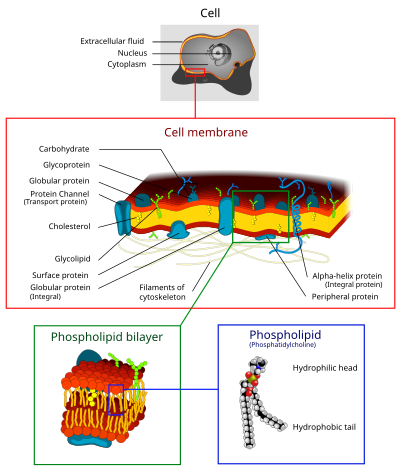

В клеточная мембрана (также известный как плазматическая мембрана (ВЕЧЕРА) или же цитоплазматическая мембрана, и исторически называемый плазмалемма) это биологическая мембрана что разделяет интерьер из всех клетки от внешняя среда (внеклеточное пространство), которое защищает клетку от окружающей среды.[1][2] Клеточная мембрана состоит из липидный бислой, включая холестерины (липидный компонент), которые находятся между фосфолипиды сохранить свои текучесть при различных температурах. Мембрана также содержит мембранные белки, включая интегральные белки которые проходят через мембрану, служащую мембранные транспортеры, и периферические белки которые свободно прикрепляются к внешней (периферической) стороне клеточной мембраны, действуя как ферменты формирование клетки.[3] Клеточная мембрана контролирует движение веществ внутри и вне клеток и органелл. Таким образом, это избирательно проницаемый к ионы и Органические молекулы.[4] Кроме того, клеточные мембраны участвуют во множестве клеточных процессов, таких как клеточная адгезия, ионная проводимость и клеточная сигнализация и служат поверхностью прикрепления для нескольких внеклеточных структур, включая клеточная стенка углеводный слой называется гликокаликс, и внутриклеточная сеть белковых волокон, называемая цитоскелет. В области синтетической биологии клеточные мембраны могут быть искусственно собранный.[5][6][7]
История
Пока Роберт Гук Открытие ячеек в 1665 г. привело к предложению Клеточная теория, Гук ввел в заблуждение теория клеточной мембраны что все клетки содержат твердую клеточную стенку, поскольку в то время можно было наблюдать только клетки растений.[8] Микроскопы сосредоточились на клеточной стенке более 150 лет, пока не были достигнуты успехи в микроскопии. В начале 19 века клетки были признаны отдельными объектами, не связанными и связанными отдельными клеточными стенками после того, как было обнаружено, что клетки растений могут быть разделены. Эта теория распространилась на клетки животных, чтобы предложить универсальный механизм защиты и развития клеток. Ко второй половине XIX века микроскопия все еще была недостаточно развита, чтобы различать клеточные мембраны и клеточные стенки. Однако некоторые микроскописты правильно определили в то время, что, хотя они невидимы, можно сделать вывод, что клеточные мембраны существуют в клетках животных из-за внутриклеточного движения компонентов внутри, а не снаружи, и что мембраны не были эквивалентом клеточной стенки для растительной клетки. Также предполагалось, что клеточные мембраны не являются жизненно важными компонентами для всех клеток. Многие опровергли существование клеточной мембраны еще в конце 19 века. В 1890 году в обновленной теории клеток было заявлено, что клеточные мембраны существуют, но являются лишь вторичными структурами. Только в более поздних исследованиях осмоса и проницаемости клеточные мембраны получили большее признание.[8] В 1895 г. Эрнест Овертон предположил, что клеточные мембраны состоят из липидов.[9]
Гипотеза липидного бислоя, предложенная в 1925 г. Гортер и Грендель,[10] создали предположение к описанию двухслойной структуры клеточной мембраны на основе кристаллографических исследований и наблюдений за мыльными пузырями. Пытаясь принять или отвергнуть эту гипотезу, исследователи измерили толщину мембраны.[8] В 1925 году Фрике определил, что толщина мембран эритроцитов и дрожжевых клеток составляет от 3,3 до 4 нм, что соответствует толщине липидного монослоя. Выбор диэлектрической проницаемости, использованной в этих исследованиях, был поставлен под сомнение, но будущие испытания не могли опровергнуть результаты первоначального эксперимента. Независимо от того, лептоскоп был изобретен для измерения очень тонких мембран путем сравнения интенсивности света, отраженного от образца, с интенсивностью эталона мембраны известной толщины. Инструмент мог определять толщину, которая зависела от измерений pH и присутствия мембранных белков в диапазоне от 8,6 до 23,2 нм, при этом более низкие измерения подтверждали гипотезу липидного бислоя. Позже, в 1930-х годах, модель мембранной структуры, по общему признанию, стала пауцимолекулярная модель из Davson и Даниэлли (1935). Эта модель была основана на исследованиях поверхностного натяжения между маслами и иглокожие яйца. Поскольку значения поверхностного натяжения оказались намного ниже, чем можно было бы ожидать для границы раздела нефть-вода, было сделано предположение, что какое-то вещество отвечает за снижение межфазного натяжения на поверхности клеток. Было высказано предположение, что липидный бислой находится между двумя тонкими слоями белка. Пауцимолекулярная модель сразу же стала популярной, и в течение следующих 30 лет она доминировала в исследованиях клеточных мембран, пока не стала конкурировать с жидкой мозаичной моделью. Певица и Николсон (1972).[11][8]
Несмотря на многочисленные модели клеточной мембраны, предложенные до жидкая мозаичная модель, он остается первичным архетипом клеточной мембраны еще долгое время после своего появления в 1970-х годах.[8] Хотя модель жидкой мозаики был модернизирован для детализации современных открытий, основы остались неизменными: мембрана представляет собой липидный бислой, состоящий из гидрофильных внешних головок и гидрофобной внутренней части, где белки могут взаимодействовать с гидрофильными головками посредством полярных взаимодействий, но белки, которые покрывают бислой полностью или частично, имеют гидрофобные аминокислоты, которые взаимодействуют с неполярной липидной внутренней частью. В модель жидкой мозаики он не только обеспечил точное представление о механике мембран, но и расширил возможности изучения гидрофобных сил, которые позже превратились в существенное описательное ограничение для описания биологических макромолекулы.[8]
На протяжении многих столетий цитируемые ученые не соглашались со значением структуры, которую они рассматривали как клеточную мембрану. В течение почти двух столетий мембраны видели, но в большинстве случаев игнорировали это как важную структуру с клеточной функцией. Только в 20 веке значение клеточной мембраны было признано. Наконец, два ученых Гортер и Грендель (1925) сделали открытие, что мембрана «основана на липидах». Исходя из этого, они продвинули идею о том, что эта структура должна быть в формации, имитирующей слои. После дальнейшего изучения было обнаружено, что путем сравнения суммы поверхностей клеток и поверхностей липидов было оценено соотношение 2: 1; таким образом, обеспечивая первую основу известной сегодня двухслойной структуры. Это открытие инициировало множество новых исследований, которые возникли во всем мире в различных областях научных исследований, подтвердив, что структура и функции клеточной мембраны широко распространены.[8]
Эта структура по-разному упоминается разными авторами как эктопласт (де Фрис, 1885),[12] Плазмахаут (плазменная кожа, Пфеффер, 1877, 1891),[13] Hautschicht (скин-слой, Пфеффер, 1886; используется в другом значении Hofmeister, 1867), плазматическая мембрана (Pfeffer, 1900),[14] плазматическая мембрана, цитоплазматическая мембрана, клеточная оболочка и клеточная мембрана.[15][16] Некоторые авторы, которые не верили в существование функциональной проницаемой границы на поверхности клетки, предпочитали использовать термин плазмалемма (придуманный Мастом, 1924) для обозначения внешней области клетки.[17][18][19]
Сочинение
Клеточные мембраны содержат множество биологические молекулы, особенно липиды и белки. Состав не задан, но постоянно меняется в зависимости от текучести и изменений окружающей среды, даже колеблется на разных стадиях развития клеток. В частности, количество холестерина в мембране первичных нейронных клеток человека изменяется, и это изменение в составе влияет на текучесть на всех стадиях развития.[20]
Материал включается в мембрану или удаляется из нее с помощью различных механизмов:
- Слияние внутриклеточных пузырьки с мембраной (экзоцитоз ) не только выводит содержимое везикулы, но также включает компоненты мембраны везикулы в клеточную мембрану. Мембрана может образовывать пузыри вокруг внеклеточного материала, который отщепляется и превращается в пузырьки (эндоцитоз ).
- Если мембрана является непрерывной с трубчатой структурой из мембранного материала, то материал из трубки может втягиваться в мембрану непрерывно.
- Хотя концентрация компонентов мембраны в водной фазе низкая (стабильные компоненты мембраны имеют низкую растворимость в воде), существует обмен молекулами между липидной и водной фазами.
Липиды

Клеточная мембрана состоит из трех классов: амфипатический липиды: фосфолипиды, гликолипиды, и стеролы. Количество каждого из них зависит от типа клетки, но в большинстве случаев фосфолипиды являются наиболее распространенными, часто составляя более 50% всех липидов в плазматических мембранах.[21][22] Гликолипиды составляют лишь небольшое количество около 2%, а стерины составляют остальное. В РБК Согласно исследованиям, 30% плазматической мембраны составляет липид. Однако для большинства эукариотических клеток состав плазматических мембран примерно наполовину состоит из липидов и наполовину из белков.
Жировые цепи в фосфолипиды и гликолипиды обычно содержат четное число атомов углерода, обычно от 16 до 20. Наиболее распространены жирные кислоты с 16 и 18 атомами углерода. Жирные кислоты могут быть насыщенными или ненасыщенными, с конфигурацией двойных связей почти всегда «цис». Длина и степень ненасыщенности цепей жирных кислот оказывают сильное влияние на текучесть мембран, поскольку ненасыщенные липиды создают изгиб, препятствуя плотной упаковке жирных кислот, тем самым уменьшая температура плавления (увеличение текучести) мембраны.[21][22] Способность некоторых организмов регулировать текучесть их клеточных мембран изменением липидного состава вызывается гомеовязкая адаптация.
Вся мембрана скрепляется посредством нековалентный взаимодействие гидрофобных хвостов, однако структура довольно текучая и не закреплена жестко на месте. Под физиологические условия молекулы фосфолипидов в клеточной мембране находятся в жидкокристаллическое состояние. Это означает, что молекулы липидов могут свободно диффундировать и проявлять быструю латеральную диффузию вдоль слоя, в котором они присутствуют.[21] Однако обмен молекулами фосфолипидов между внутриклеточными и внеклеточными листочками бислоя - очень медленный процесс. Липидные рафты и кавеолы являются примерами холестерин -обогащенные микродомены в клеточной мембране.[22] Кроме того, фракция липида, непосредственно контактирующая с интегральными мембранными белками, которая прочно связана с поверхностью белка, называется кольцевая липидная оболочка; он ведет себя как часть белкового комплекса.
В клетках животных холестерин обычно в различной степени диспергирован по клеточным мембранам, в нерегулярных пространствах между гидрофобными хвостами мембранных липидов, где он оказывает эффект жесткости и укрепления на мембрану.[4] Кроме того, количество холестерина в биологических мембранах варьируется между организмами, типами клеток и даже в отдельных клетках. Холестерин, основной компонент плазматических мембран животных, регулирует текучесть всей мембраны, а это означает, что холестерин контролирует количество движения различных компонентов клеточной мембраны в зависимости от его концентрации.[4] При высоких температурах холестерин подавляет движение цепей фосфолипидных жирных кислот, вызывая снижение проницаемости для малых молекул и снижение текучести мембран. Обратное верно для роли холестерина в низких температурах. Производство холестерина и, следовательно, его концентрация регулируется (увеличивается) в ответ на низкую температуру. При низких температурах холестерин мешает взаимодействию цепей жирных кислот. Действуя как антифриз, холестерин поддерживает текучесть мембраны. Холестерин больше у животных в холодную погоду, чем у животных в теплую погоду. В растениях, в которых отсутствует холестерин, родственные соединения, называемые стеролами, выполняют ту же функцию, что и холестерин.[4]
Фосфолипиды, образующие липидные пузырьки
Липидные везикулы или липосомы представляют собой примерно сферические карманы, заключенные в липидный бислой.[23] Эти структуры используются в лабораториях для изучения воздействия химических веществ на клетки, доставляя эти химические вещества непосредственно в клетку, а также для более глубокого понимания проницаемости клеточных мембран. Липидные везикулы и липосомы образуются путем сначала суспендирования липида в водном растворе, а затем перемешивания смеси через обработка ультразвуком, в результате чего образуется пузырек. Измеряя скорость излияние от внутренней части везикулы до окружающего раствора, позволяет исследователю лучше понять проницаемость мембраны. Везикулы могут быть образованы с молекулами и ионами внутри везикулы путем образования везикулы с желаемой молекулой или ионом, присутствующими в растворе. Белки также могут быть встроены в мембрану путем солюбилизации желаемых белков в присутствии детергентов и присоединения их к фосфолипидам, в которых образуется липосома. Они предоставляют исследователям инструмент для изучения различных функций мембранных белков.
Углеводы
Плазменные мембраны также содержат углеводы, преимущественно гликопротеины, но с некоторыми гликолипидами (цереброзиды и ганглиозиды ). Углеводы играют важную роль клеточное распознавание у эукариот; они расположены на поверхности клетки, где они распознают клетки-хозяева и обмениваются информацией, вирусы, которые связываются с клетками с помощью этих рецепторов, вызывают инфекцию [24] По большей части нет гликозилирование происходит на мембранах внутри клетки; чаще всего гликозилирование происходит на внеклеточной поверхности плазматической мембраны. В гликокаликс это важная особенность всех клеток, особенно эпителий с микроворсинками. Последние данные предполагают, что гликокаликс участвует в адгезии клеток, возвращение лимфоцитов,[24] и много других. В предпоследний сахар это галактоза а конечный сахар сиаловая кислота, поскольку сахарный остов изменен в аппарат Гольджи. Сиаловая кислота несет отрицательный заряд, создавая внешний барьер для заряженных частиц.
Белки
| Тип | Описание | Примеры |
| Интегральные белки или трансмембранные белки | Растяните мембрану и получите гидрофильную цитозольный домен, который взаимодействует с внутренними молекулами, гидрофобным охватывающим мембрану доменом, который закрепляет его внутри клеточной мембраны, и гидрофильным внеклеточным доменом, который взаимодействует с внешними молекулами. Гидрофобный домен состоит из одного, нескольких или комбинации α-спирали и β лист белок мотивы. | Ионные каналы, протонные насосы, Рецептор, связанный с G-белком |
| Заякоренные липиды белков | Ковалентно связан с одной или несколькими молекулами липидов; гидрофобно вставляется в клеточную мембрану и закрепляет белок. Сам белок не контактирует с мембраной. | G белки |
| Периферические белки | Присоединены к интегральным мембранным белкам или связаны с периферическими участками липидного бислоя. Эти белки, как правило, только временно взаимодействуют с биологическими мембранами, и после реакции молекула диссоциирует, чтобы продолжить свою работу в цитоплазме. | Некоторые ферменты, некоторые гормоны |
Клеточная мембрана имеет большое количество белков, обычно около 50% от объема мембраны.[25] Эти белки важны для клетки, потому что они отвечают за различные биологические активности. Примерно треть гены в дрожжи код специально для них, а у многоклеточных организмов это число еще больше.[23] Мембранные белки состоят из трех основных типов: интегральные белки, периферические белки и липидно-заякоренные белки.[4]
Как показано в соседней таблице, интегральные белки представляют собой амфипатические трансмембранные белки. Примеры интегральных белков включают ионные каналы, протонные насосы и рецепторы, связанные с g-белком. Ионные каналы позволяют неорганическим ионам, таким как натрий, калий, кальций или хлор, диффундировать вниз по своему электрохимическому градиенту через липидный бислой через гидрофильные поры через мембрану. Электрическое поведение клеток (т.е. нервных клеток) контролируется ионными каналами.[4] Протонные насосы - это протеиновые насосы, встроенные в липидный бислой, которые позволяют протонам перемещаться через мембрану, переходя от одной боковой цепи аминокислоты к другой. В таких процессах, как перенос электронов и генерирование АТФ, используются протонные насосы.[4] Рецептор, связанный с G-белком, представляет собой одиночную полипептидную цепь, которая семь раз пересекает липидный бислой, отвечая на сигнальные молекулы (то есть гормоны и нейротрансмиттеры). Рецепторы, сопряженные с G-белком, используются в таких процессах, как передача сигналов от клетки к клетке, регулирование продукции цАМФ и регулирование ионных каналов.[4]
Клеточная мембрана, подвергающаяся воздействию внешней среды, является важным местом межклеточной коммуникации. Таким образом, большое количество белковых рецепторов и идентификационных белков, таких как антигены, присутствуют на поверхности мембраны. Функции мембранных белков могут также включать межклеточный контакт, распознавание поверхности, контакт с цитоскелетом, передачу сигналов, ферментативную активность или транспортировку веществ через мембрану.
Большинство мембранных белков необходимо каким-то образом встроить в мембрану.[26] Чтобы это произошло, N-концевая «сигнальная последовательность» аминокислот направляет белки к эндоплазматический ретикулум, который вставляет белки в липидный бислой. После вставки белки затем транспортируются к месту назначения в везикулах, где везикула сливается с целевой мембраной.
Функция


Клеточная мембрана окружает цитоплазма живых клеток, физически разделяющих внутриклеточный компоненты из внеклеточный среда. Клеточная мембрана также играет роль в закреплении цитоскелет чтобы придать форму ячейке, и при прикреплении к внеклеточный матрикс и другие ячейки, чтобы удерживать их вместе, чтобы сформировать ткани. Грибы, бактерии, наиболее археи, и растения также есть клеточная стенка, который обеспечивает механическую поддержку клетки и препятствует прохождению более крупные молекулы.
Клеточная мембрана избирательно проницаемый и может регулировать, что входит и выходит из клетки, тем самым облегчая транспорт материалов, необходимых для выживания. Движение веществ через мембрану может быть либо "пассивный ", происходящие без ввода клеточной энергии, или"активный ", требуя, чтобы клетка тратила энергию на ее транспортировку. Мембрана также поддерживает потенциал клетки. Таким образом, клеточная мембрана работает как избирательный фильтр, который позволяет только определенным вещам попадать внутрь или выходить из клетки. Клетка задействует ряд транспортных механизмов, в которых задействованы биологические мембраны:
1. Пассивный осмос и распространение: Некоторые вещества (небольшие молекулы, ионы), такие как диоксид углерода (CO2) и кислород (O2), могут перемещаться через плазматическую мембрану посредством диффузии, которая представляет собой пассивный транспортный процесс. Поскольку мембрана действует как барьер для определенных молекул и ионов, они могут встречаться в различных концентрациях на двух сторонах мембраны. Диффузия происходит, когда небольшие молекулы и ионы свободно перемещаются от высокой концентрации к низкой, чтобы уравновесить мембрану. Это считается пассивным транспортным процессом, потому что он не требует энергии и приводится в движение градиентом концентрации, создаваемым каждой стороной мембраны.[27] Такой градиент концентрации через полупроницаемую мембрану создает осмотический поток для воды. Осмос в биологических системах включает в себя растворитель, движущийся через полупроницаемую мембрану аналогично пассивной диффузии, поскольку растворитель все еще движется с градиентом концентрации и не требует энергии. Хотя вода является наиболее распространенным растворителем в ячейке, это могут быть и другие жидкости, а также сверхкритические жидкости и газы.[28]
2. Трансмембранные белковые каналы и транспортеры: Трансмембранные белки проходят через липидный бислой мембран; они действуют по обе стороны мембраны, перемещая через нее молекулы.[29] Питательные вещества, такие как сахар или аминокислоты, должны поступать в клетку, а некоторые продукты метаболизма должны покидать клетку. Такие молекулы могут пассивно диффундировать через белковые каналы, такие как аквапорины в облегченное распространение или перекачиваются через мембрану трансмембранные транспортеры. Белки протеиновых каналов, также называемые проникает, обычно довольно специфичны, они распознают и переносят только ограниченный набор химических веществ, часто ограниченный одним веществом. Другим примером трансмембранного белка является рецептор клеточной поверхности, который позволяет сигнальным молекулам клетки связываться между клетками.[29]
3. Эндоцитоз: Эндоцитоз - это процесс, при котором клетки поглощают молекулы, поглощая их. Плазматическая мембрана создает небольшую деформацию внутрь, называемую впячиванием, при которой транспортируемое вещество захватывается. Это инвагинация вызывается белками снаружи на клеточной мембране, действующими как рецепторы и объединяющимися в углубления, которые в конечном итоге способствуют накоплению большего количества белков и липидов на цитозольной стороне мембраны.[30] Затем деформация отрывается от мембраны внутри клетки, создавая везикулу, содержащую захваченное вещество. Эндоцитоз - это путь интернализации твердых частиц ("поедание клеток" или фагоцитоз ), маленькие молекулы и ионы ("питье клетки" или пиноцитоз ) и макромолекулы. Эндоцитоз требует энергии и, таким образом, является формой активного транспорта.
4. Экзоцитоз: Так же, как материал может быть доставлен в клетку путем инвагинации и образования везикулы, мембрана везикулы может сливаться с плазматической мембраной, вытесняя ее содержимое в окружающую среду. Это процесс экзоцитоза. Экзоцитоз происходит в различных клетках для удаления непереваренных остатков веществ, внесенных в результате эндоцитоза, для выделения таких веществ, как гормоны и ферменты, и для полного переноса вещества через клеточный барьер. В процессе экзоцитоза непереваренная пищевая вакуоль, содержащая отходы, или секреторный пузырек отпочковываются из аппарат Гольджи, сначала перемещается цитоскелетом изнутри клетки на поверхность. Мембрана везикул контактирует с плазматической мембраной. Молекулы липидов двух бислоев перестраиваются, и две мембраны, таким образом, сливаются. В слитой мембране образуется проход, и везикулы выводят свое содержимое за пределы клетки.
Прокариоты
Прокариоты делятся на две разные группы, Археи и Бактерии, при этом бактерии делятся на грамположительный и грамотрицательный. Грамотрицательные бактерии имеют как плазматическую мембрану, так и внешняя мембрана разделены по периплазма однако другие прокариоты есть только плазматическая мембрана. Эти две мембраны различаются по многим аспектам. Наружная мембрана грамотрицательных бактерий отличается от других прокариот тем, что фосфолипиды формируя внешнюю часть бислоя, и липопротеины и фосфолипиды, образующие внутреннюю часть.[31] Внешняя мембрана обычно имеет пористую структуру из-за присутствия в ней мембранных белков, таких как грамотрицательные порины, которые являются порообразующими белками. Внутренняя плазматическая мембрана также обычно симметрична, тогда как внешняя мембрана асимметрична из-за белков, таких как вышеупомянутые. Кроме того, что касается прокариотических мембран, есть несколько факторов, которые могут повлиять на текучесть. Одним из основных факторов, которые могут повлиять на текучесть, является состав жирных кислот. Например, когда бактерии Золотистый стафилококк был выращен в 37◦C в течение 24 часов мембрана показывала более жидкое состояние вместо гелеобразного состояния. Это подтверждает идею о том, что при более высоких температурах мембрана более текучая, чем при более низких температурах. Когда мембрана становится более жидкой и нуждается в большей стабилизации, она образует более длинные цепи жирных кислот или цепи насыщенных жирных кислот, чтобы помочь стабилизировать мембрану.[32] Бактерии также окружены клеточная стенка состоит из пептидогликан (аминокислоты и сахара). Некоторые эукариотические клетки также имеют клеточные стенки, но ни одна из них не состоит из пептидогликана. Наружная мембрана грамотрицательных бактерий богата липополисахариды, которые представляют собой комбинированные поли- или олигосахаридные и углеводные липидные области, которые стимулируют естественный иммунитет клетки.[33] Наружная мембрана может пузырь в периплазматические выпячивания в условиях стресса или при необходимости вирулентности при встрече с клеткой-мишенью, и, таким образом, такие пузырьки могут работать как органеллы вирулентности.[34] Бактериальные клетки представляют собой многочисленные примеры разнообразных способов адаптации мембран прокариотических клеток к структурам, которые соответствуют нише организма. Например, белки на поверхности некоторых бактериальных клеток помогают в их скользящем движении.[35] Многие грамотрицательные бактерии имеют клеточные мембраны, которые содержат управляемые АТФ системы экспорта белков.[35]
Структуры
Модель жидкой мозаики
Согласно модель жидкой мозаики из С. Дж. Сингер и Г. Л. Николсон (1972), которые заменили более ранние модель Дэвсона и Даниелли, биологические мембраны можно рассматривать как двумерная жидкость в котором молекулы липидов и белков диффундируют более или менее легко.[36] Хотя липидные бислои, образующие основу мембран, действительно сами по себе образуют двумерные жидкости, плазматическая мембрана также содержит большое количество белков, которые обеспечивают большую структуру. Примерами таких структур являются белковые комплексы, пикеты и заграждения, образованные актином на основе цитоскелет, и потенциально липидные рафты.
Липидный бислой

Липидные бислои форма в процессе самосборка. Клеточная мембрана состоит в основном из тонкого слоя амфипатический фосфолипиды которые спонтанно располагаются так, что гидрофобные «хвостовые» области изолированы от окружающей воды, в то время как гидрофильные «головные» области взаимодействуют с внутриклеточными (цитозольными) и внеклеточными поверхностями образующегося бислоя. Это образует непрерывный сферический липидный бислой. Гидрофобные взаимодействия (также известные как гидрофобный эффект ) являются основными движущими силами в формировании липидных бислоев. Увеличение взаимодействий между гидрофобными молекулами (вызывающее кластеризацию гидрофобных областей) позволяет молекулам воды более свободно связываться друг с другом, увеличивая энтропию системы. Это сложное взаимодействие может включать нековалентные взаимодействия, такие как ван дер Ваальс, электростатические и водородные связи.
Липидные бислои обычно непроницаемы для ионов и полярных молекул. Расположение гидрофильных головок и гидрофобных хвостов липидного бислоя предотвращает диффузию полярных растворенных веществ (например, аминокислот, нуклеиновых кислот, углеводов, белков и ионов) через мембрану, но обычно обеспечивает пассивную диффузию гидрофобных молекул. Это дает клетке возможность контролировать движение этих веществ через трансмембранный белок комплексы, такие как поры, каналы и ворота.Флиппасы и скрамбласы концентрировать фосфатидилсерин, несущий отрицательный заряд на внутренней мембране. Вместе с NANA, это создает дополнительный барьер для заряженных части движется через мембрану.
Мембраны выполняют различные функции в эукариотический и прокариотический клетки. Одна из важных ролей - регулирование движения материалов в клетки и из них. Двухслойная структура фосфолипидов (модель жидкой мозаики) со специфическими мембранными белками объясняет избирательную проницаемость мембраны, а также пассивные и активные механизмы транспорта. Кроме того, мембраны прокариот, митохондрий и хлоропластов эукариот способствуют синтезу АТФ посредством хемиосмоса.
Полярность мембраны
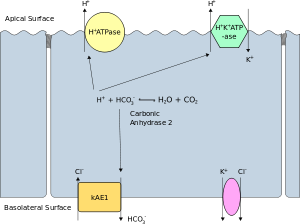
Апикальная мембрана поляризованной клетки - это поверхность плазматической мембраны, которая обращена внутрь к просвет. Это особенно заметно в эпителиальный и эндотелиальные клетки, но также описывает другие поляризованные ячейки, такие как нейроны. В базолатеральная мембрана Поляризованной клетки является поверхность плазматической мембраны, которая образует ее базальную и боковую поверхности. Он обращен наружу, к интерстиций, и подальше от просвета. Базолатеральная мембрана - это сложная фраза, относящаяся к терминам «базальная (базовая) мембрана» и «боковая (боковая) мембрана», которые, особенно в эпителиальных клетках, идентичны по составу и активности. Белки (например, ионные каналы и насосы ) могут свободно перемещаться от базальной к боковой поверхности клетки или наоборот в соответствии с модель жидкой мозаики. Узкие стыки присоединяются к эпителиальным клеткам около их апикальной поверхности, чтобы предотвратить миграцию белков с базолатеральной мембраны на апикальную мембрану. Таким образом, базальная и боковая поверхности остаются примерно одинаковыми.[требуется разъяснение ] друг к другу, но отличные от апикальной поверхности.
Мембранные конструкции

Клеточная мембрана может образовывать различные типы «супрамембранных» структур, такие как Caveola, постсинаптическая плотность, подосома, инвадоподиум, очаговая адгезия, и разные типы клеточные соединения. Эти структуры обычно отвечают за клеточная адгезия, коммуникация, эндоцитоз и экзоцитоз. Их можно визуализировать электронная микроскопия или же флуоресцентная микроскопия. Они состоят из определенных белков, таких как интегрины и кадгерины.
Цитоскелет
В цитоскелет находится под клеточной мембраной в цитоплазме и обеспечивает каркас для мембранных белков, к которым они прикрепляются, а также образуют органеллы которые выходят из клетки. В самом деле, цитоскелетные элементы широко и тесно взаимодействуют с клеточной мембраной.[37] Заякоренные белки ограничивают их определенной клеточной поверхностью - например, апикальной поверхностью эпителиальных клеток, выстилающих позвоночное животное кишка - и ограничивает, насколько далеко они могут диффундировать внутри бислоя. Цитоскелет способен образовывать органеллы, похожие на придатки, такие как реснички, которые микротрубочка -основанные расширения, покрытые клеточной мембраной, и филоподии, которые актин на основе расширений. Эти расширения заключены в мембрану и выступают из поверхности клетки, чтобы воспринимать внешнюю среду и / или вступать в контакт с субстратом или другими клетками. Апикальные поверхности эпителиальных клеток плотные с актиновыми пальцеобразными выступами, известными как микроворсинки, которые увеличивают площадь поверхности клеток и тем самым увеличивают скорость поглощения питательных веществ. Локальное разъединение цитоскелета и клеточной мембраны приводит к образованию пузырь.
Внутриклеточные мембраны
Содержимое клетки внутри клеточной мембраны состоит из многочисленных мембраносвязанных органелл, которые вносят вклад в общую функцию клетки. Происхождение, структура и функция каждой органеллы приводят к большим вариациям в составе клеток из-за индивидуальной уникальности, связанной с каждой органеллой.
- Считается, что митохондрии и хлоропласты произошли от бактерий, известных как эндосимбиотическая теория. Эта теория возникла из идеи, что Паракокк и РодопсоводомонадаТипы бактерий имеют схожие функции с митохондриями и сине-зелеными водорослями, или цианобактерии, имеют схожие функции с хлоропластами. В эндосимбиотическая теория предполагает, что в ходе эволюции эукариотическая клетка поглотила эти 2 типа бактерий, что привело к образованию митохондрий и хлоропластов внутри эукариотических клеток. Это поглощение привело к двум системам мембран этих органелл, в которых внешняя мембрана произошла от плазматической мембраны хозяина, а внутренняя мембрана была плазматической мембраной эндосимбионта. Учет того, что митохондрии и хлоропласты содержат свою собственную ДНК, является дополнительным подтверждением того, что обе эти органеллы произошли от поглощенных бактерий, которые процветали внутри эукариотической клетки.[38]
- В эукариотических клетках ядерная мембрана отделяет содержимое ядра от цитоплазмы клетки.[39] Ядерная мембрана образована внутренней и внешней мембранами, обеспечивая строгую регуляцию поступления материалов в ядро и из него. Материалы перемещаются между цитозолем и ядром через ядерные поры в ядерной мембране. Если ядро клетки более активно в транскрипция, его мембрана будет иметь больше пор. Белковый состав ядра может сильно отличаться от цитозоля, так как многие белки не могут проникать через поры посредством диффузии. Внутри ядерной мембраны внутренняя и внешняя мембраны различаются по белковому составу, и только внешняя мембрана является непрерывной с эндоплазматический ретикулум (ER) мембрана. Как и ER, внешняя мембрана также содержит рибосомы, ответственные за производство и транспортировку белков в пространство между двумя мембранами. Ядерная мембрана разбирается на ранних стадиях митоза и собирается на более поздних стадиях митоза.[40]
- ER, который является частью эндомембранной системы, составляет очень большую часть общего содержимого мембран клетки. ER представляет собой замкнутую сеть канальцев и мешочков, и его основные функции включают синтез белка и метаболизм липидов. Есть 2 типа ER: гладкая и грубая. Грубый ER имеет прикрепленные к нему рибосомы, используемые для синтеза белка, в то время как гладкий ER используется больше для обработки токсинов и регуляции кальция в клетке.[41]
- В аппарат Гольджи имеет две соединенные между собой круглые цистерны Гольджи. Отделения аппарата образуют множественные трубчато-ретикулярные сети, отвечающие за организацию, соединение штабелей и транспортировку грузов, которые отображают непрерывные, похожие на виноград, струнные везикулы размером от 50 до 60 нм. Аппарат состоит из трех основных отделов, плоской дискообразной цистерны с трубчато-ретикулярной сеткой и пузырьков.[42]
Вариации
Клеточная мембрана имеет различный липидный и белковый состав в разных типы ячеек и поэтому могут иметь определенные имена для определенных типов ячеек.
- Сарколемма в миоциты: «Сарколемма» - это название клеточной мембраны миоцитов (также известных как мышечные клетки).[43] Хотя сарколемма похожа на другие клеточные мембраны, у нее есть другие функции, которые ее отличают. Например, сарколемма передает синаптические сигналы, помогает генерировать потенциалы действия и очень участвует в сокращениях мышц.[44] В отличие от других клеточных мембран, сарколемма состоит из небольших каналов, называемых «Т-канальцами», которые проходят через все мышечные клетки. Также было обнаружено, что средняя сарколемма имеет толщину 10 нм в отличие от 4 нм толщины обычной клеточной мембраны.[45][43]
- Оолемма - это клеточная мембрана в ооциты: Оолемма ооцитов (незрелых яйцеклеток) не соответствует липидному бислою, поскольку в них отсутствует бислой и они не состоят из липидов.[46] Скорее, структура имеет внутренний слой, оболочку для оплодотворения, а внешний слой состоит из желточного слоя, который состоит из гликопротеинов; однако каналы и белки все еще присутствуют для выполнения своих функций в мембране.
- Аксолемма: Специализированная плазматическая мембрана на аксоны нервных клеток, отвечающих за генерацию потенциала действия. Он состоит из гранулированного, плотно упакованного липидного бислоя, который тесно взаимодействует с компонентами цитоскелета спектрином и актином. Эти компоненты цитоскелета способны связываться с трансмембранными белками аксолеммы и взаимодействовать с ними.[47][48]
Проницаемость
В проницаемость мембраны - это скорость пассивного распространение молекул через мембрану. Эти молекулы известны как проницаемый молекулы. Проницаемость зависит в основном от электрический заряд и полярность молекулы и в меньшей степени молярная масса молекулы. Из-за гидрофобной природы клеточной мембраны небольшие электрически нейтральные молекулы проходят через мембрану легче, чем заряженные большие. Неспособность заряженных молекул проходить через клеточную мембрану приводит к раздел pH веществ во всем отсеки для жидкости тела.
Смотрите также
- Кольцевая липидная оболочка
- Искусственная ячейка
- Структура бактериальной клетки
- Синдром Бангстада
- Клеточная кора
- Повреждение клеток, в том числе повреждение клеточной мембраны
- Клеточная теория
- Цитонема
- Эластичность клеточных мембран
- Грамположительные бактерии
- Мембранные модели
- Мембранные нанотрубочки
- История теории клеточных мембран
- Липидный плот
- Трогоцитоз
Примечания и ссылки
- ^ Страницы биологии Кимбалла В архиве 2009-01-25 на Wayback Machine, Клеточные мембраны
- ^ Синглтон П (1999). Бактерии в биологии, биотехнологии и медицине (5-е изд.). Нью-Йорк: Вили. ISBN 978-0-471-98880-9.
- ^ Том Херрманн1; Сандип Шарма 2. (2 марта 2019 г.). «Физиология, мембрана». StatPearls. 1 Медицинский факультет SIU 2 Баптистский региональный медицинский центр. PMID 30855799.CS1 maint: использует параметр авторов (связь) CS1 maint: location (связь)
- ^ а б c d е ж грамм час Альбертс Б., Джонсон А., Льюис Дж. И др. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Наука о гирляндах. ISBN 978-0-8153-3218-3. В архиве из оригинала от 20.12.2017.
- ^ Будин I, Деварадж Н.К. (январь 2012 г.). «Мембранная сборка, управляемая реакцией биомиметического связывания». Журнал Американского химического общества. 134 (2): 751–3. Дои:10.1021 / ja2076873. ЧВК 3262119. PMID 22239722.
- ^ Персонал (25 января 2012 г.). «Химики синтезируют искусственную клеточную мембрану». ScienceDaily. В архиве из оригинала 29 января 2012 г.. Получено 18 февраля, 2012.
- ^ Персонал (26 января 2012 г.). «Химики создают искусственную клеточную мембрану». kurzweilai.net. В архиве из оригинала 26 февраля 2012 г.. Получено 18 февраля, 2012.
- ^ а б c d е ж грамм Ломбард J (декабрь 2014 г.). «Давным-давно клеточные мембраны: 175 лет исследований границ клеток». Биология Директ. 9: 32. Дои:10.1186 / s13062-014-0032-7. ЧВК 4304622. PMID 25522740.
- ^ Лерэ, С. Хронологическая история липидного центра. Киберлипид Центр. Последнее обновление 11 ноября 2017 г. связь В архиве 2017-10-13 на Wayback Machine.
- ^ Гортер Э, Грендель Ф (март 1925 г.). «О бимолекулярных слоях липоидов на хромоцитах крови». Журнал экспериментальной медицины. 41 (4): 439–43. Дои:10.1084 / jem.41.4.439. ЧВК 2130960. PMID 19868999.
- ^ Сингер С. Дж. И Николсон Г. Л. "Жидкая мозаичная модель структуры клеточных мембран". Наука. (1972) 175. 720-731.
- ^ де Фриз Х (1885). "Plasmolytische Studien über die Wand der Vakuolen". Jahrb. Wiss. Бот. 16: 465–598.
- ^ Pfeffer, W. 1877. Osmotische Untersuchungen: Studien zur Zell Mechanik. Энгельманн, Лейпциг.
- ^ Пфеффер, В., 1900–1906. Физиология растений, [1] В архиве 2018-06-02 в Wayback Machine. Перевод А. Й. Юарта со 2-го немецкого изд. из Pflanzenphysiologie, 1897-1904, [2] В архиве 2018-06-01 в Wayback Machine. Кларендон Пресс, Оксфорд.
- ^ Шарп, Л. У. (1921). Введение в цитологию. Нью-Йорк: Макгроу Хилл, стр. 42.
- ^ Kleinzeller, A. 1999. Концепция клеточной мембраны Чарльза Эрнеста Овертона. В: Проницаемость мембраны: 100 лет со времен Эрнеста Овертона (под ред. Димера Д.В., Кляйнцеллера А., Фамброу Д.М.), стр. 1–18, Academic Press, San Diego, [3].
- ^ Мачта СО (1924 г.). "Структура и движение в Амеба протей". Анат. Rec. 29 (2): 88. Дои:10.1002 / ар.1090290205.
- ^ Плау JQ (1931). «Мембраны в растительной клетке. I. Морфологические мембраны на протоплазматических поверхностях». Протоплазма. 12: 196–220. Дои:10.1007 / BF01618716.
- ^ Уэйн Р. (2009). Биология клетки растений: от астрономии к зоологии. Амстердам: Elsevier / Academic Press. п. 17. ISBN 9780080921273.
- ^ Ноутси П., Граттон Э, Чайеб С. (30.06.2016). «Оценка колебаний текучести мембран во время клеточного развития выявляет время и специфичность типа клеток». PLOS ONE. 11 (6): e0158313. Bibcode:2016PLoSO..1158313N. Дои:10.1371 / journal.pone.0158313. ЧВК 4928918. PMID 27362860.
- ^ а б c Лодиш Х., Берк А., Зипурский Л.С. и др. (2000). «Биомембраны: структурная организация и основные функции». Молекулярная клеточная биология (4-е изд.). Нью-Йорк: Книги Scientific American. ISBN 978-0-7167-3136-8.
- ^ а б c Купер GM (2000). «Строение плазменной мембраны». Клетка: молекулярный подход (2-е изд.). В архиве из оригинала от 19.09.2017.
- ^ а б Лодиш Х, Берк А, Зипурски С.Л., Мацудаира П., Балтимор Д., Дарнелл Дж. (2000). «Биомембраны: структурная организация и основные функции». Молекулярная клеточная биология (4-е изд.). В архиве из оригинала 2018-06-05.
- ^ а б Брэндли Б.К., Шнаар Р.Л. (июль 1986 г.). «Углеводы клеточной поверхности в распознавании и ответе клеток». Журнал биологии лейкоцитов. 40 (1): 97–111. Дои:10.1002 / jlb.40.1.97. PMID 3011937.
- ^ Джесси Грей; Шана Грошлер; Тони Ли; Зара Гонсалес (2002). «Мембранная структура» (SWF). Дэвидсон-колледж. В архиве из оригинала 2007-01-08. Получено 2007-01-11.
- ^ Лодиш Х, Берк А, Зипурски С.Л., Мацудаира П., Балтимор Д., Дарнелл Дж. (2000). «Посттрансляционные модификации и контроль качества в грубой ER». Молекулярная клеточная биология (4-е изд.).
- ^ Купер, Джеффри М. (2000). «Транспорт малых молекул». Клетка: молекулярный подход (2-е изд.). В архиве из оригинала 2018-06-05.
- ^ Крамер Э.М., Майерс Д.Р. (апрель 2013 г.). «Осмос не вызывается разбавлением водой». Тенденции в растениеводстве. 18 (4): 195–7. Дои:10.1016 / j.tplants.2012.12.001. PMID 23298880.
- ^ а б Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Мембранные белки». Молекулярная биология клетки (4-е изд.). В архиве из оригинала 2018-06-05.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Транспорт в клетку из плазматической мембраны: эндоцитоз». Молекулярная биология клетки (4-е изд.). Наука о гирляндах. В архиве из оригинала 2018-06-05.
- ^ Солтон MR, Ким К. (1996). Барон С. (ред.). Медицинская микробиология (4-е изд.). Галвестон (Техас): Медицинский филиал Техасского университета в Галвестоне. ISBN 978-0963117212. PMID 21413343.
- ^ Мишра Н.Н., Лю Г.Ю., Йеман М.Р., Наст СС, Проктор Р.А., Маккиннелл Дж., Байер А.С. (февраль 2011 г.). «Связанное с каротиноидами изменение текучести клеточной мембраны влияет на чувствительность Staphylococcus aureus к пептидам защиты хозяина». Противомикробные препараты и химиотерапия. 55 (2): 526–31. Дои:10.1128 / AAC.00680-10. ЧВК 3028772. PMID 21115796.
- ^ Александр C, Rietschel ET (2001). «Бактериальные липополисахариды и врожденный иммунитет». Журнал исследований эндотоксинов. 7 (3): 167–202. Дои:10.1177/09680519010070030101. PMID 11581570.
- ^ Яшрой RC (1999). «Структурная модель органелл вирулентности грамотрицательных организмов со ссылкой на патогенность сальмонелл в подвздошной кишке курицы». Индийский журнал птицеводства. 34 (2): 213–219. В архиве из оригинала от 07.11.2014.
- ^ а б Сайер MH (2013). «Микрокомпартменты и белковые машины прокариот». Журнал молекулярной микробиологии и биотехнологии. 23 (4–5): 243–69. Дои:10.1159/000351625. ЧВК 3832201. PMID 23920489.
- ^ Певица SJ, Николсон GL (февраль 1972 г.). «Жидкая мозаичная модель структуры клеточных мембран». Наука. 175 (4023): 720–31. Bibcode:1972Научный ... 175..720С. Дои:10.1126 / science.175.4023.720. PMID 4333397.
- ^ Доэрти Г.Дж., МакМахон ХТ (2008). «Посредничество, модуляция и последствия взаимодействий мембрана-цитоскелет». Ежегодный обзор биофизики. 37: 65–95. Дои:10.1146 / annurev.biophys.37.032807.125912. PMID 18573073. S2CID 17352662.
- ^ Ватли Дж. М., Джон П., Ватли Ф. Р. (апрель 1979 г.). «От внеклеточного к внутриклеточному: создание митохондрий и хлоропластов». Труды Лондонского королевского общества. Серия B, Биологические науки. 204 (1155): 165–87. Bibcode:1979RSPSB.204..165W. Дои:10.1098 / rspb.1979.0020. PMID 36620.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Структура и функции ДНК». Молекулярная биология клетки (4-е изд.). Наука о гирляндах.
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). «Транспорт молекул между ядром и цитозолем». Молекулярная биология клетки (4-е изд.). Наука о гирляндах.
- ^ Купер GM (2000). «Эндоплазматический ретикулум». Клетка: молекулярный подход (2-е изд.). В архиве из оригинала от 03.10.2017.
- ^ Сюй Х, Су В, Цай М, Цзян Дж, Цзэн Х, Ван Х (2013-04-16). «Асимметричная структура мембран аппарата Гольджи, выявленная с помощью атомно-силового микроскопа in situ». PLOS ONE. 8 (4): e61596. Bibcode:2013ПРИЛОЖЕНИЕ ... 861596X. Дои:10.1371 / journal.pone.0061596. ЧВК 3628984. PMID 23613878.
- ^ а б Рид Р., Вьюстон Т.В., Тодд П.М. (июль 1966 г.). «Строение и функция сарколеммы скелетных мышц». Природа. 211 (5048): 534–6. Bibcode:1966Натура.211..534R. Дои:10.1038 / 211534b0. PMID 5967498.
- ^ Кэмпбелл К.П., Стулл Д.Т. (апрель 2003 г.). "Взаимодействие базальной мембраны скелетных мышц-сарколеммы-цитоскелета" серия мини-обзоров ". Журнал биологической химии. 278 (15): 12599–600. Дои:10.1074 / jbc.r300005200. PMID 12556456.
- ^ Митра К., Убарретксена-Беландия I, Тагучи Т., Уоррен Г., Энгельман Д.М. (март 2004 г.). «Модуляция толщины бислоя мембран экзоцитарного пути мембранными белками, а не холестерином». Труды Национальной академии наук Соединенных Штатов Америки. 101 (12): 4083–8. Bibcode:2004ПНАС..101.4083М. Дои:10.1073 / pnas.0307332101. ЧВК 384699. PMID 15016920.
- ^ Wessel GM, Wong JL (октябрь 2009 г.). «Изменения клеточной поверхности яйца при оплодотворении». Молекулярное воспроизводство и развитие. 76 (10): 942–53. Дои:10.1002 / мрд.21090. ЧВК 2842880. PMID 19658159.
- ^ Рейн С.С. (1999). «Характеристики нейрона». Основы нейрохимии: молекулярные, клеточные и медицинские аспекты (6-е изд.).
- ^ Фитцпатрик МО, Максвелл В.Л., Грэм Д.И. (март 1998 г.). «Роль аксолеммы в инициировании травматического повреждения аксонов». Журнал неврологии, нейрохирургии и психиатрии. 64 (3): 285–7. Дои:10.1136 / jnnp.64.3.285. ЧВК 2169978. PMID 9527135.
внешняя ссылка
- Липиды, мембраны и обмен везикул - виртуальная библиотека биохимии и клеточной биологии
- Протокол выделения белка из клеточной мембраны
- Мембранный гомеостаз, регулирование натяжения, механочувствительный мембранный обмен и мембранный трафик
- 3D-структуры белков, связанных с плазматической мембраной эукариотических клеток
- Липидный состав и белки мембран некоторых эукариот
- [4]