Метапопуляция - Metapopulation
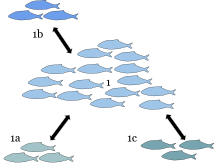
А метапопуляция состоит из группы пространственно разделенных популяций одного и того же разновидность которые взаимодействуют на каком-то уровне. Термин «метапопуляция» был придуман Ричард Левинс в 1969 г. для описания модели динамика населения насекомых-вредителей на сельскохозяйственных полях, но эта идея наиболее широко применялась к видам, естественным или искусственным фрагментированные среды обитания. По словам Левинса, она состоит из «популяции популяций».[1]
Обычно считается, что метапопуляция состоит из нескольких отдельных популяций вместе с территориями подходящей среды обитания, которые в настоящее время не заняты. В классической теории метапопуляции каждая популяция циклически меняется в относительной независимости от других популяций и в конечном итоге вымирает в результате демографических изменений. стохастичность (колебания численности населения из-за случайных демографический События); чем меньше популяция, тем больше шансов инбридинговой депрессии и склонности к исчезновению.
Хотя отдельные популяции имеют ограниченную продолжительность жизни, метапопуляция в целом часто остается стабильной, потому что иммигранты из одной популяции (которая может, например, испытывать демографический бум), вероятно, повторно колонизируют среду обитания, которая была оставлена открытой в результате исчезновения. другой популяции. Они также могут эмигрировать в небольшую популяцию и спасти эту популяцию от вымирания (так называемая спасательный эффект). Такой спасательный эффект может произойти, потому что сокращение численности населения оставляет нишевые возможности открытыми для «спасателей».
Развитие теории метапопуляции в сочетании с развитием динамика источник-сток, подчеркнула важность связи между, казалось бы, изолированными группами населения. Хотя ни одна отдельная популяция не может гарантировать долгосрочное выживание данного вида, совокупный эффект многих популяций может сделать это.
Теория метапопуляции была впервые разработана для наземных экосистем, а затем была применена к морской сфере.[2] В науке о рыболовстве термин «подгруппа» эквивалентен термину «местное население» в науке о метапопуляции. Большинство морских примеров представлено относительно оседлыми видами, населяющими отдельные участки среды обитания, как с местным набором, так и с привлечением других местных популяций в более крупной метапопуляции. Kritzer & Sale выступили против строгого применения критериев определения метапопуляции, согласно которым риски вымирания для местного населения должны быть существенными.[2]:32
Финский биолог Илкка Хански из Университет Хельсинки внес важный вклад в теорию метапопуляции.
Хищничество и колебания
Первые эксперименты с хищничество и пространственная неоднородность проводились Г. Ф. Гаузе в 1930-е годы на основе Уравнение Лотки – Вольтерра, который был сформулирован в середине 1920-х годов, но дальнейшее применение не проводилось.[3] Уравнение Лотки-Вольтерры предполагает, что отношения между хищниками и их добычей приведут к популяции колебания с течением времени в зависимости от начальной плотности хищника и жертвы. Ранние эксперименты Гаузе по доказательству предсказанных колебаний этой теории не увенчались успехом, потому что иммиграция не повлияла на взаимодействия хищник-жертва. Однако с введением иммиграции популяционные циклы точно отображали колебания, предсказанные уравнением Лотки-Вольтерра, с пиками численности жертв, немного сдвинутыми влево от пиков плотности хищников. Эксперименты Хаффакера расширили эксперименты Гаузе, изучив, как факторы миграции и пространственная неоднородность приводят к колебаниям хищник-жертва.
Эксперименты Хаффакера по взаимодействию хищник-жертва (1958)
Чтобы изучить хищники и колебания популяции, Хаффакер использовал виды клещей, один из которых был хищником, а другой - добычей.[4] Он поставил управляемый эксперимент, используя апельсины, которыми питалась добыча, как пространственно структурированную среду обитания, в которой хищник и жертва будут взаимодействовать.[5] Поначалу Хаффакер испытывал трудности, аналогичные трудностям Гаузе, в создании стабильного взаимодействия хищник-жертва. Использование только апельсинов привело к тому, что виды-жертвы быстро вымерли, а затем и хищники. Однако он обнаружил, что, изменяя пространственную структуру среды обитания, он может управлять динамикой популяции и обеспечивать повышение общей выживаемости для обоих видов. Он делал это, изменяя расстояние между добычей и апельсинами (их пищей), устанавливая барьеры для передвижения хищников и создавая коридоры, по которым жертва может разойтись.[3] Эти изменения привели к увеличению числа участков среды обитания и, в свою очередь, предоставили жертве больше мест для поиска временной защиты. Когда жертва вымирала локально на одном участке среды обитания, она могла восстановиться, мигрируя на новые участки, прежде чем подвергнуться нападению хищников. Эта пространственная структура мест обитания пятен позволяла сосуществовать между хищниками и жертвами и способствовала стабильной модели колебаний популяции.[6] Хотя термин «метапопуляция» еще не появился, факторы окружающей среды пространственная неоднородность а неоднородность среды обитания позже будет описывать условия метапопуляции, связанные с тем, как группы пространственно разделенных популяций видов взаимодействуют друг с другом. Эксперимент Хаффакера важен, потому что он показал, как метапопуляции могут напрямую влиять на взаимодействия хищник-жертва и, в свою очередь, влиять на динамику популяции.[7]
Модель Левинса
Первоначальная модель Левинса применялась к метапопуляции, распределенной по множеству участков подходящей среды обитания со значительно меньшим взаимодействием между участками, чем внутри участка. Динамика популяции внутри участка была упрощена до такой степени, что учитывались только наличие и отсутствие. Каждый патч в его модели либо заселен, либо нет.
Позволять N быть долей участков, занятых в данный момент. В течение времени dt, каждый занятый участок может стать незанятым с вероятностью исчезновения EDT. Дополнительно 1 -N патчей не заняты. Предполагая постоянную скорость c из пропагула поколение от каждого из N занятые участки, в течение времени dt, каждый незанятый участок может быть занят с вероятностью колонизации cNdt . Соответственно, скорость смены занятых участков, дН / дт, является

Это уравнение математически эквивалентно логистическая модель, с грузоподъемностью K данный

и темп роста р

Следовательно, при равновесии некоторая часть среды обитания вида всегда будет незанятой.
Стохастичность и метапопуляции
Хаффакера[4] Исследования пространственной структуры и межвидовых взаимодействий являются примером ранних экспериментов в динамике метапопуляции. Поскольку эксперименты Хаффакера[4] и Левинс,[1] созданы модели, объединяющие стохастические факторы. Эти модели показали, что сочетание изменчивости окружающей среды (стохастичности) и относительно небольшой скорости миграции приводит к неопределенной или непредсказуемой устойчивости. Однако эксперимент Хаффакера почти гарантировал бесконечную настойчивость из-за контролируемой иммиграционной переменной.
Стохастические модели занятости участков (SPOM)
Одним из основных недостатков модели Левинса является то, что она детерминированный, тогда как фундаментальные процессы метапопуляции стохастический. Метапопуляции особенно полезны при обсуждении видов в нарушенных среда обитания, а жизнеспособность их популяций, т.е. насколько вероятно, что они станут вымерший в заданный промежуток времени. Модель Левинса не может решить эту проблему. Простой способ расширить модель Левинса, включив в нее пространственные и стохастические соображения, - это использовать контактный процесс. Простые модификации этой модели также могут включать динамика патча. При данном порог перколяции, фрагментация среды обитания В этих конфигурациях имеют место эффекты, предсказывающие более резкие пороги вымирания.[8]
За биология сохранения Для целей модели метапопуляции должны включать (а) конечный характер метапопуляций (сколько участков подходит для среды обитания) и (б) вероятностный характер вымирания и колонизации. Также обратите внимание, что для применения этих моделей исчезновение и заселение пятен должно быть асинхронным.
Пятна микробычи (MHP) и бактериальные метапопуляции
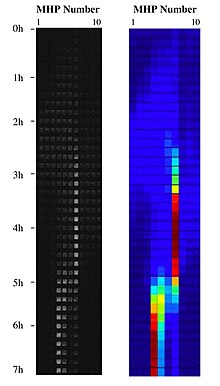
Объединив нанотехнологии с ландшафтная экология, ландшафт среды обитания можно создать на кристалле с помощью нанотехнологий, построив коллекцию наноразмерных сред обитания бактерий и соединив их коридорами в различных топологических схемах и наноразмерными каналами, обеспечивающими их локальную экосистемная услуга обновления среды обитания. Эти ландшафты MHP могут использоваться как физические реализации адаптивный ландшафт:[9] путем создания пространственной мозаики из кусочков возможностей, распределенных в пространстве и времени. Неровная природа этих жидких ландшафтов позволяет изучать адаптацию бактериальных клеток в системе метапопуляции, работающей на кристалле внутри синтетической экосистемы. Биология метапопуляции и эволюционная экология этих бактериальных систем в этих синтетических экосистемах можно решить с помощью экспериментальных биофизика.
Эволюция истории жизни
Модели метапопуляции использовались для объяснения эволюции жизненного цикла, такие как экологическая стабильность из амфибия метаморфоза в маленьком весенние пруды. Появились альтернативные экологические стратегии. Например, некоторые саламандры отказываются от метаморфоза и становятся половозрелыми как водные животные. неотены. Сезонная продолжительность водно-болотных угодий и миграционный ареал видов определяет, какие пруды связаны между собой и образуют ли они метапопуляцию. Продолжительность этапов жизненного цикла земноводных по отношению к продолжительности весенней лужи до ее высыхания регулирует экологическое развитие метапопуляций, соединяющих водные участки с участками суши.[10]
Смотрите также
- Конкуренция (биология)
- Биология сохранения
- Ландшафтная экология
- Уравнения Лотки – Вольтерра
- Колебание
- Анализ жизнеспособности популяции
- Хищничество
- Пространственная неоднородность
Рекомендации
- ^ а б Левинс, Р. (1969), "Некоторые демографические и генетические последствия неоднородности окружающей среды для биологического контроля", Бюллетень энтомологического общества Америки, 15 (3): 237–240, Дои:10.1093 / besa / 15.3.237
- ^ а б Критцер, Дж. П. и Сэйл, П. Ф. (редакторы) (2006) Морские метапопуляции, Academic Press, Нью-Йорк.
- ^ а б Реал, Лесли А. и Браун, Джеймс Х. 1991. Основы экологии: классические статьи с комментариями. Издательство Чикагского университета, Чикаго.
- ^ а б c Хаффакер, C.B. (1958), "Экспериментальные исследования хищничества: факторы дисперсии и колебания хищник-жертва", Хильгардия, 27 (343): 343–383, Дои:10.3733 / hilg.v27n14p343
- ^ Legendre, P .; Фортин, М.Дж. (1989), "Пространственный образец и экологический анализ", Экология растений, 80 (2): 107, CiteSeerX 10.1.1.330.8940, Дои:10.1007 / BF00048036
- ^ Карейва П. (1987), "Фрагментация среды обитания и стабильность взаимодействий хищник-жертва", Природа, 326 (6111): 388–390, Bibcode:1987Натура.326..388K, Дои:10.1038 / 326388a0
- ^ Янссен, А. и др. 1997. Динамика метапопуляции устойчивой системы хищник – жертва.
- ^ Кеймер Дж. Э; П.А. Марке; Х. Х. Веласко-Эрнандес; Левин С.А. (ноябрь 2000 г.). «Пороги вымирания и устойчивость метапопуляции в динамических ландшафтах». Американский натуралист. 156 (5): 478–4945. Дои:10.1086/303407. HDL:10533/172124. PMID 29587508.
- ^ Keymer J.E .; П. Галайда; К. Малдун Р. и Р. Остин (ноябрь 2006 г.). «Бактериальные метапопуляции в нанотехнологических ландшафтах». PNAS. 103 (46): 17290–295. Bibcode:2006ПНАС..10317290К. Дои:10.1073 / pnas.0607971103. ЧВК 1635019. PMID 17090676.
- ^ Петранка, Дж. У. (2007), «Эволюция сложных жизненных циклов земноводных: преодоление разрыва между динамикой метапопуляции и эволюцией жизненного цикла», Эволюционная экология, 21 (6): 751–764, Дои:10.1007 / s10682-006-9149-1.
- Bascompte J .; Соле Р. В. (1996), "Пороги фрагментации и исчезновения среды обитания в пространственно явных моделях", Журнал экологии животных, 65 (4): 465–473, Дои:10.2307/5781, JSTOR 5781.
- Ганский, И. Экология метапопуляции Издательство Оксфордского университета. 1999 г. ISBN 0-19-854065-5
- Фариг, Л. 2003. Влияние фрагментации среды обитания на биоразнообразие. Ежегодный обзор экологии, эволюции и систематики. 34: 1, с. 487.
- Левин С.А. (1974), "Дисперсия и взаимодействие популяций", Американский натуралист, 108 (960): 207, Дои:10.1086/282900.