Экологическая преемственность - Ecological succession

Экологическая преемственность это процесс изменения в разновидность структура экологическое сообщество через некоторое время. Временной масштаб может составлять десятилетия (например, после пожара) или даже миллионы лет после массовое вымирание.[1]
Сообщество начинается с относительно небольшого числа пионеры растений и животных и развивается путем увеличения сложности, пока не станет стабильным или самовоспроизводящийся как климакс сообщество. «Двигатель» преемственности, причина изменения экосистемы - это воздействие устоявшихся организмов на их собственную среду обитания. Следствием жизни является иногда незаметное, а иногда и явное изменение собственного окружения.[2]
Это явление или процесс, с помощью которого экологическое сообщество претерпевает более или менее упорядоченные и предсказуемые изменения после беспокойство или первоначальное заселение новой среды обитания. Сукцессия может быть инициирована либо образованием новой незанятой среды обитания, например, из поток лавы или серьезный оползень, или какой-либо формой беспокойство сообщества, например, из Огонь, суровый метание ветра, или же протоколирование. Сукцессия, которая начинается в новых средах обитания, не подверженная влиянию ранее существовавших сообществ, называется первичное правопреемство, тогда как преемственность, которая следует за разрушением ранее существовавшего сообщества, называется вторичная последовательность.
Преемственность была одной из первых теорий, выдвинутых в экология. Экологическая преемственность была впервые задокументирована в дюнах Индианы на северо-западе Индианы и остается в основе многих экологическая наука.[3]
История
Предшественники идеи экологической преемственности восходят к началу XIX века. Французский натуралист Адольф Дюро де ла Маль был первым, кто употребил слово преемственность о развитии растительности после рубок леса. В 1859 г. Генри Дэвид Торо написал обращение под названием «Наследие лесных деревьев»[4] в котором он описал последовательность в дубово-сосновом лесу. «Наблюдателям давно известно, что белки зарывают орехи в землю, но я не знаю, чтобы кто-то таким образом объяснил регулярную смену лесов».[5] Австрийский ботаник Антон Кернер опубликовали исследование о смене растений в Дунай бассейн реки в 1863 году.[6]
Х. К. Коулз

Генри Чендлер Коулз, на Чикагский университет, разработал более формальную концепцию наследования. Вдохновленный исследованиями Датский дюны Евгений Уорминг, Коулз изучал растительность застройка на песчаных дюнах на берегу озеро Мичиган (в Indiana Dunes ). Он признал, что растительность на дюнах разного возраста можно интерпретировать как разные стадии общей тенденции развития растительности на дюнах (подход к изучению изменения растительности, позже названный замещением пространства-времени, или хронопоследовательность исследования). Он впервые опубликовал эту работу в качестве статьи в Ботанический вестник в 1899 г. («Экологические отношения растительности песчаных дюн озера Мичиган»).[7] В этой классической публикации и последующих статьях он сформулировал идею первичной преемственности и понятие sere - повторяющаяся последовательность изменений сообщества, специфичных для конкретных условий окружающей среды.[3][8]
Глисон и Клементс
Однако примерно с 1900 по 1960 годы в понимании преемственности преобладали теории Фредерик Клементс, современник Коулза, который считал, что сериалы очень предсказуемы и детерминированы и сходятся на климатически обусловленных конюшнях. климакс сообщество независимо от стартовых условий. Клементс явно сравнил сукцессионное развитие экологических сообществ с онтогенетическим развитием отдельных организмов, и его модель часто называют псевдоорганической теорией экологии сообществ. Клементс и его последователи разработали сложную таксономию сообществ и последовательных путей.
Генри Глисон предложили контрастирующую основу еще в 1920-х годах. Модель Глисона была более сложной и гораздо менее детерминированной, чем модель Клемента. Он наиболее существенно отличается от точки зрения Клемента, предполагая гораздо большую роль случайные факторы и в отрицании существования согласованных, строго ограниченных типов сообществ. Глисон утверждал, что распределение видов индивидуально реагирует на факторы окружающей среды, и сообщества лучше всего рассматривать как артефакты сопоставления распределений видов. Идеи Глисона, впервые опубликованные в 1926 году, в значительной степени игнорировались до конца 1950-х годов.
Две цитаты иллюстрируют противоположные взгляды Клементса и Глисона. Клементс писал в 1916 году:
Изучение развития растительности обязательно основывается на предположении, что единица или климактерическая формация является органической единицей. Как организм, образование возникает, растет, созревает и умирает. Более того, каждое климактерическое образование способно воспроизводить себя, с существенной точностью повторяя этапы своего развития.
— Фредерик Клементс[9]
в то время как Глисон в своей статье 1926 года сказал:
Ассоциация - это не организм, даже не растительная единица, а просто совпадение.
— Генри Глисон[10]
На самом деле идеи Глисона больше соответствовали первоначальному мышлению Коулза о преемственности. О различении Климентса между первичное правопреемство и вторичная последовательность, Cowles писал (1911):
Эта классификация не представляет фундаментальной ценности, поскольку она разделяет такие тесно связанные явления, как эрозия и отложение, и объединяет такие непохожие вещи, как деятельность человека и оседание земли.
— Генри Коулз[11]
Современная эра
Более тщательное тестирование сукцессионных моделей и теории сообщества на основе данных обычно начиналось с работы Роберт Уиттакер и Джон Кертис в 1950-1960-х гг. С тех пор теория преемственности стала менее монолитной и более сложной. Дж. Коннелл и Р. Слейер предпринял попытку кодификации сукцессионных процессов по механизму. Среди британских и североамериканских экологов понятие конюшни климакс растительность был в значительной степени заброшен, и сукцессионные процессы стали рассматриваться как менее детерминированные, с важной ролью исторической случайность и для альтернативных путей в реальном развитии сообществ. Споры продолжаются относительно общей предсказуемости сукцессионной динамики и относительной важности равновесных и неравновесных процессов. Бывший профессор Гарварда Ф.А. Баззаз ввел понятие шкала в дискуссию, поскольку он считал, что в локальном масштабе или в масштабе небольшой территории процессы являются случайными и неоднородными, но, принимая во внимание более крупные региональные районы, нельзя отрицать определенные тенденции.[12]
Факторы
На траекторию сукцессионных изменений могут влиять условия участка, характер событий, инициирующих сукцессию (возмущения), взаимодействия присутствующих видов и многое другое. стохастический такие факторы, как наличие колонистов или семян или Погода условия на момент нарушения. Некоторые из этих факторов способствуют предсказуемости динамики сукцессии; другие добавляют больше вероятностный элементы. Два важных фактора возмущения сегодня: человеческие действия и изменение климата.[13]
В целом в сообществах в ранней преемственности будут преобладать быстрорастущие, хорошо развитыерассредоточенный разновидность (оппортунист, беглец, или же r-выбранный истории жизни). По мере продолжения сукцессии эти виды будут замещаться более конкурентоспособными (k-выбранный ) разновидность.
Тенденции в экосистема были предложены следующие друг за другом объекты собственности, но лишь немногие из них являются общими. Например, видовое разнообразие почти обязательно увеличивается во время ранней сукцессии по мере появления новых видов, но может снижаться в более поздней сукцессии, поскольку конкуренция устраняет условно-патогенные виды и приводит к преобладание местных конкурентов. Чистая первичная производительность, биомасса, и трофический Все свойства отображают изменяющиеся закономерности в последовательности, в зависимости от конкретной системы и сайта.
Раньше считалось, что экологическая сукцессия имеет стабильную конечную стадию, называемую кульминация, иногда называемые «потенциальной растительностью» участка, и формируются в основном под влиянием местного климата. Современные экологи отказались от этой идеи в пользу неравновесных представлений о динамике экосистем. Большинство природных экосистем испытывают нарушение со скоростью, которая делает недостижимым "кульминационный момент" сообщества. Изменение климата часто происходит с частотой и частотой, достаточными для предотвращения наступления климакса. Дополнения к доступным пулам видов за счет расширения ареала и введения также может постоянно изменять сообщества.
Развитие некоторых атрибутов экосистемы, таких как свойства почвы и питательные циклы, оба находятся под влиянием собственности сообщества и, в свою очередь, влияют на дальнейшее сукцессионное развитие. Этот процесс обратной связи может происходить только на протяжении столетий или тысячелетий. В сочетании с стохастический природа возмущающих событий и других долгосрочных (например, климатических) изменений, такая динамика делает сомнительным, будет ли когда-либо применяться концепция «кульминации» или она особенно полезна при рассмотрении фактической растительности.
Типы
Первичная преемственность
Сукцессионная динамика, начинающаяся с заселения территории, которая ранее не была занята экологическим сообществом, такая как недавно обнаженные поверхности скал или песка, потоки лавы, недавно обнаженные ледниковые холмы и т. Д., Называются первичной сукцессией. К стадиям первичной сукцессии относятся микроорганизмы-пионеры,[14] растения (лишайники и мхи), травянистые растения, более мелкие кусты и деревья. Животные начинают возвращаться, когда есть пища для них. Когда это полностью функционирующая экосистема, она достигает кульминационной стадии сообщества.[15]
Вторичное правопреемство
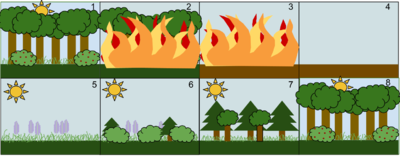
1. Устойчивое сообщество лиственных лесов
2. Возмущение, например лесной пожар, уничтожает лес.
3. Огонь сжигает лес дотла.
4. Огонь оставляет после себя пустую, но не уничтоженную землю.
5. Сначала отрастают травы и другие травянистые растения.
6. Небольшие кусты и деревья начинают заселять территорию.
7. Быстрорастущие вечнозеленые деревья развиваются в полной мере, тогда как теневыносливые деревья развиваются в подлеске.
8. Недолговечные вечнозеленые деревья, не переносящие тени, умирают, когда большие лиственные деревья покрывают их. Экосистема вернулась в то же состояние, в котором она начиналась.
Сукцессионная динамика после серьезного нарушения или удаления ранее существовавшего сообщества называется вторичной сукцессией. На динамику вторичной сукцессии сильно влияют условия, предшествующие возмущению, включая развитие почвы, семенные банки, остаточное органическое вещество и остаточные живые организмы. Из-за остаточной фертильности и ранее существовавших организмов изменение сообщества на ранних стадиях вторичной сукцессии может быть относительно быстрым.
Вторичная сукцессия наблюдается и изучается гораздо чаще, чем первичная. Наиболее распространенные типы вторичной сукцессии включают реакцию на естественные возмущения, такие как пожар, наводнение и сильные ветры, а также на антропогенные нарушения, такие как лесозаготовки и сельское хозяйство. Во вторичной сукцессии почвы и организмы необходимо оставить нетронутыми, чтобы у нового материала был путь к восстановлению.[4]
Например, во фрагментированной старой полевой среде обитания, созданной в восточной части Канзаса, древесные растения «быстрее колонизировались (на единицу площади) на больших и близлежащих территориях. патчи ".[16]

Вторичная преемственность может быстро изменить ландшафт. В 1900-х годах Национальный парк Акадия произошел лесной пожар, уничтоживший большую часть ландшафта. Первоначально в ландшафте росли вечнозеленые деревья. После пожара на этом участке росли кусты не менее года. В конце концов, вместо вечнозеленых растений начали расти лиственные деревья.[15]
Вторичная преемственность происходила в Национальный парк Шенандоа после наводнения 1995 г. Мурмана и реки Рапидан, уничтожившие растительный и животный мир.[17]
Сезонная и циклическая динамика
В отличие от вторичной сукцессии, эти типы изменения растительности не зависят от беспокойство но являются периодическими изменениями, возникающими из-за изменчивых взаимодействий видов или повторяющихся событий. Эти модели изменяют концепция кульминации к одному из динамических состояний.
Причины сукцессии растений
Аутогенная сукцессия могут быть вызваны изменениями в почве, вызванными присутствующими там организмами. Эти изменения включают накопление органических веществ в подстилке или гуминовом слое, изменение питательных веществ в почве или изменение pH почвы из-за растущих там растений. Структура самих растений также может изменить сообщество. Например, когда более крупные виды, такие как деревья, созревают, они создают тень на развивающейся лесной подстилке, которая, как правило, исключает светолюбивые виды. Теневыносливые виды будут вторгаться в этот район.
Аллогенная преемственность вызвано внешними воздействиями окружающей среды, а не растительностью. Например, изменения почвы из-за эрозии, выщелачивания или отложения ила и глин могут изменить содержание питательных веществ и водные отношения в экосистемах. Животные также играют важную роль в аллогенных изменениях, поскольку они опылители, разносчики семян и травоядные. Они также могут увеличивать содержание питательных веществ в почве в определенных областях или перемещать почву (как это делают термиты, муравьи и кроты), создавая пятна в среде обитания. Это может создать участки регенерации, благоприятствующие определенным видам.
Климатические факторы могут быть очень важными, но в гораздо более длительном масштабе, чем любые другие. Изменения температуры и режима осадков будут способствовать изменениям в сообществах. По мере потепления климата в конце каждого ледникового периода происходили большие сукцессионные изменения. Растительность тундры и голые ледниковые отложения сменились смешанными широколиственными лесами. В парниковый эффект повышение температуры может привести к глубоким аллогенным изменениям в следующем столетии. Геологические и климатические катастрофы, такие как извержения вулканов, землетрясения, лавины, метеоры, наводнения, пожары и сильный ветер, также вызывают аллогенные изменения.
Механизмы
В 1916 г. Фредерик Клементс опубликовал описательную теорию сукцессии и выдвинул ее как общую экологическую концепцию.[9] Его теория преемственности оказала сильное влияние на экологическую мысль. Концепцию Клементса принято называть классической. экологическая теория По словам Клементса, преемственность - это процесс, включающий несколько этапов:[9][страница нужна ]
- Nudation: преемственность начинается с развития голого участка, называемого Nudation (нарушение).[9]
- Миграция: относится к прибытию пропагулы.[9]
- Ecesis: включает укоренение и начальный рост растительности.[9]
- Конкуренция: по мере того, как растительность становится устойчивой, растет и распространяется, различные виды начинают конкурировать за пространство, свет и питательные вещества.[9]
- Реакция: во время этой фазы аутогенные изменения, такие как накопление гумуса, влияют на среду обитания, и одно растительное сообщество сменяет другое.[9]
- Стабилизация: формируется якобы стабильное климаксное сообщество.[9]
Серальные сообщества


Семейное сообщество - это промежуточный этап в экосистеме, продвигающейся к своему климакс сообщество. Во многих случаях до наступления климакса проходит более одной стадии развития.[18] А Prisere представляет собой набор серов, составляющих развитие области от не покрытых растительностью поверхностей до климаксового сообщества. В зависимости от субстрата и климата встречаются разные серы.
Изменения в жизни животных
Теория сукцессии была разработана в первую очередь ботаниками. Изучение преемственности применимо ко всему экосистемы инициирован в трудах Рамон Маргалеф, пока Евгений Одум Публикация Стратегия развития экосистемы считается его формальной отправной точкой.[19]
Жизнь животных также претерпевает изменения с изменением сообществ. В лишайниковой стадии фауна редка. Он состоит из нескольких клещей, муравьев и пауков, обитающих в трещинах и щелях. Качественный рост фауны происходит на стадии разнотравья. Животные, обнаруженные на этой стадии, включают нематод, личинок насекомых, муравьев, пауков, клещей и т. Д. Популяция животных увеличивается и диверсифицируется с развитием сообщества лесных климаксов. Фауна состоит из беспозвоночных, таких как слизни, улитки, черви, многоножки, многоножки, муравьи, жуки; и позвоночные животные, такие как белки, лисы, мыши, кроты, змеи, различные птицы, саламандры и лягушки.
Микросессия
Преемственность микроорганизмы включая грибы и бактерии происходящие в пределах микроместа обитания, известны как микросцессии или серулы. Как и в случае с растениями, микробная сукцессия может происходить в новых доступных средах обитания (первичное правопреемство ), такие как поверхности листьев растений, недавно обнаженные каменные поверхности (например, ледниковый тилль) или кишки новорожденных животных,[14] а также на нарушенных сообществах (вторичная последовательность ), например, растущие на недавно мертвых деревьях или помете животных. Сообщества микробов также могут меняться из-за продуктов, выделяемых присутствующими бактериями. Изменения pH в среде обитания могут создать идеальные условия для обитания в этом районе нового вида. В некоторых случаях новые виды могут превзойти нынешние по питательным веществам, что приведет к гибели основных видов. Изменения также могут происходить в результате микробной сукцессии с вариациями доступности воды и температуры. Теории макроэкология только недавно были применены к микробиология и еще так много предстоит понять об этой растущей области. Недавнее исследование микробной сукцессии оценило баланс между стохастический и детерминированные процессы в бактериальной колонизации солончаков хронопоследовательность. Результаты этого исследования показывают, что, как и в макросессии, ранняя колонизация (первичное правопреемство ) в основном зависит от стохастичности, в то время как на вторичную последовательность этих бактериальных сообществ сильнее влияют детерминированные факторы.[20]
Концепция кульминации
Согласно классической экологическая теория, последовательность прекращается, когда сера достигает равновесия или устойчивого состояния с физической и биотической средой. За исключением серьезных нарушений, он будет сохраняться бесконечно. Эта конечная точка преемственности называется кульминацией.
Climax сообщество
Конечное или стабильное сообщество в серии - это климакс сообщество или же климатическая растительность. Он самовоспроизводящийся и находится в равновесии с физической средой обитания. В климаксовом сообществе нет чистого годового накопления органических веществ. Годовое производство и использование энергии в таком сообществе сбалансировано.
Характеристики
- Растительность толерантна к условиям окружающей среды.
- Он имеет большое разнообразие видов, хорошо дренированную пространственную структуру и сложные пищевые цепи.
- Экосистема климакс сбалансирована. Существует равновесие между валовая первичная продукция и общее дыхание, между энергией, используемой солнечным светом, и энергией, выделяемой при разложении, между поглощением питательных веществ из почвы и возвратом питательных веществ из подстилки в почву.
- Люди в стадии кульминации заменяются другими такого же типа. Таким образом, видовой состав сохраняет равновесие.
- Это показатель климата местности. Форма жизни или роста указывает на климатический тип.
Типы климакса
- Климатический климакс
- Если существует только одна кульминация и развитие климаксового сообщества контролируется климатом региона, это называется климатической кульминацией. Например, развитие климаксового сообщества кленово-букового на влажной почве. Климатический климат является теоретическим и развивается там, где физические условия субстрата не являются настолько экстремальными, чтобы изменять влияние преобладающего регионального климата.
- Эдафическая кульминация
- Когда в регионе существует более одного климаксового сообщества, измененного местными условиями субстрата, такими как влажность почвы, питательные вещества почвы, топография, воздействие на склоны, пожары и активность животных, это называется эдафическая кульминация. Последовательность заканчивается эдафической кульминацией, когда рельеф, почва, вода, огонь или другие нарушения таковы, что климатическая кульминация не может развиться.
- Катастрофическая кульминация
- Растительность кульминации уязвима для катастрофического события, такого как лесной пожар. Например, в Калифорния, чапараль растительность - это последняя растительность. Лесной пожар удаляет зрелую растительность и разлагатели. Затем следует быстрое развитие травянистой растительности, пока не восстановится доминирование кустарника. Это известно как катастрофическая кульминация.
- Дисклимакс
- Когда устойчивое сообщество, которое не является климатической или эдафической кульминацией для данного участка, поддерживается человеком или его домашними животными, это обозначается как Disclimax (кульминация беспокойства) или антропогенный субклимакс (вызванный человеком). Например, чрезмерный выпас на складах могут образовывать сообщества кустарников и кактусов в пустыне, где местный климат фактически позволяет пастбищам сохраняться.
- Субклимакс
- Продолжительная стадия, предшествующая климатической кульминации, - это субклимакс.
- Предклимакс и постклимакс
- В определенных районах в одинаковых климатических условиях развиваются разные климаксовые сообщества. Если в сообществе есть формы жизни более низкие, чем в ожидаемой климатической кульминации, это называется преклимакс; сообщество, в котором формы жизни выше, чем в ожидаемой климатической кульминации, постклимакс. Полосы преклимакса развиваются в менее влажных и более жарких областях, тогда как пряди постклимакса развиваются в более влажных и прохладных областях, чем в окружающем климате.
Теории
Существует три школы интерпретаций, объясняющих концепцию кульминации:
- Теория моноклимакс или климатического климакс была выдвинута Clements (1916) и признает только одну кульминацию, характеристики которой определяются исключительно климатом (климатическая климакс). Процессы сукцессии и модификации окружающей среды преодолевают влияние различий в топографии, материнском материале почвы и других факторах. Вся территория будет покрыта однородным растительным сообществом. Сообщества, отличные от кульминации, связаны с ним и распознаются как субклимакс, постклимакс и дисклимакс.
- Теория поликлимакса была развита Тэнсли (1935). Он предполагает, что кульминационная фаза растительности региона состоит из более чем одной кульминационной точки, контролируемой влажностью почвы, питательными веществами почвы, топографией, экспозицией склонов, пожарами и деятельностью животных.
- Теория паттернов кульминации была предложена Уиттакером (1953). Теория паттернов климакса признает множество кульминаций, определяемых реакцией популяций видов на биотические и абиотические условия. Согласно этой теории, общая среда экосистемы определяет состав, видовую структуру и баланс климаксового сообщества. Окружающая среда включает в себя реакцию видов на влажность, температуру и питательные вещества, их биотические отношения, наличие флоры и фауны для колонизации территории, случайное распространение семян и животных, почвы, климат и такие нарушения, как огонь и ветер. Характер кульминационной растительности будет меняться по мере изменения окружающей среды. Сообщество климакса представляет собой образец популяций, который соответствует образцу окружающей среды и изменяется вместе с ним. Центральным и наиболее распространенным сообществом является климатический климакс.
Теория альтернативные стабильные состояния предполагает, что существует не одна конечная точка, а множество, которые переходят друг в друга в течение экологического времени.
Наследование леса

Леса, являясь экологической системой, подвержены процессу сукцессии видов.[21] Существуют «оппортунистические» или «пионерные» виды, которые производят большое количество семян, которые разносятся ветром, и поэтому могут колонизировать большие пустые участки. Они способны прорастать и расти под прямыми солнечными лучами. Как только они создали закрытый навес отсутствие прямого солнечного излучения на почве затрудняет развитие их собственных саженцев. Тогда это возможность для теневыносливый виды, чтобы утвердиться под защитой пионеров. Когда пионеры умирают, их заменяют теневыносливые виды. Эти виды способны расти под пологом, а значит, при отсутствии катастроф останутся. По этой причине тогда говорят, что стоять достигла своего апогея. Когда случается катастрофа, для пионеров снова открывается возможность, при условии, что они присутствуют или находятся в разумных пределах.
Примером первопроходцев в лесах северо-востока Северной Америки являются Betula papyrifera (Белая береза ) и Prunus serotina (Черная вишня ), которые особенно хорошо приспособлены для использования больших пробелов в лесных пологах, но не переносят тени и в конечном итоге заменяются другими теневыносливый виды при отсутствии нарушений, создающих такие разрывы.
Вещи в природе не черно-белые, и есть промежуточные стадии. Поэтому нормально, что между двумя крайностями света и тени существует градиент, и есть виды, которые могут действовать как первопроходцы или терпимые, в зависимости от обстоятельств. Чрезвычайно важно знать толерантность видов, чтобы практиковать эффективное лесоводство.
Смотрите также
- Модель экологической сукцессии Коннелла – Слейера
- Циклическая последовательность
- Экологическая устойчивость
- Гипотеза промежуточного возмущения
Рекомендации
- ^ Sahney, S .; Бентон, М.Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен». Труды Королевского общества B: биологические науки. 275 (1636): 759–65. Дои:10.1098 / rspb.2007.1370. ЧВК 2596898. PMID 18198148.
- ^ «Виртуальная природная тропа в Пенсильвании в Нью-Кенсингтоне». Государственный университет Пенсильвании. Получено 10 октября, 2013.
- ^ а б Смит, С. и Марк, С. (2009). Исторические корни охраны природы в регионе Северо-Западная Индиана / Чикаголенд: от науки к сохранению. Журнал Южного берега, 3. «Архивная копия». Архивировано из оригинал на 2016-01-01. Получено 2015-11-22.CS1 maint: заархивированная копия как заголовок (связь)
- ^ а б Череда лесных деревьев и диких яблок. Archive.org. Проверено 12 апреля 2014.
- ^ Торо, Х. Д. (2013). Очерки: Полностью аннотированное издание (Дж. С. Крамер, ред.). Нью-Хейвен, Коннектикут: Издательство Йельского университета.
- ^ Баззаз, Ф.А. (1996). Растения в меняющейся среде. Великобритания: Издательство Кембриджского университета. п. 3. ISBN 9-780521-398435.
- ^ Э.К. Коулз (1899). "Экологические связи растительности песчаных дюн озера Мичиган. Часть I. Географические связи флоры дюн". Ботанический вестник. Издательство Чикагского университета. 27 (2): 95–117. Дои:10.1086/327796. S2CID 84315469.
- ^ Шонс, Мэри. "Генри Чендлер Коулз". Национальная география. Получено 25 июн 2014.
- ^ а б c d е ж грамм час я Клементс, Фредерик Э. (1916) Последовательность растений: анализ развития растительности
- ^ Глисон, Генри А. (1926) Индивидуалистическая концепция ассоциации растений. Бюллетень Ботанического клуба Торри
- ^ Коулз, Генри С. (1911) Причины вегетационных циклов. Анналы Ассоциации американских географов, 1 (1): 3-20 [1]
- ^ Баззаз, Ф.А. (1996). Растения в меняющейся среде. Великобритания: Издательство Кембриджского университета. С. 4–5. ISBN 9-780521-398435.
- ^ Баззаз, Ф.А. (1996). Растения в меняющейся среде. Великобритания: Издательство Кембриджского университета. п. 1. ISBN 9-780521-398435.
- ^ а б Ортис-Альварес, Рюдигер; Фирер, Ной; де лос Риос, Асунсьон; Casamayor, Emilio O .; Барберан, Альберт (2018). «Последовательные изменения таксономической структуры и функциональных характеристик бактериальных сообществ в ходе первичной сукцессии». Журнал ISME. 12 (7): 1658–1667. Дои:10.1038 / с41396-018-0076-2. ISSN 1751-7370. ЧВК 6018800. PMID 29463893.
- ^ а б Редакторы, Б.Д. (31.01.2017). «Экологическая преемственность - определение, виды и примеры». Биологический словарь. Получено 2019-05-08.CS1 maint: дополнительный текст: список авторов (связь)
- ^ Cook, W.M .; Yao, J .; Foster, B.L .; Holt, R.D .; Патрик, Л. «Вторичная последовательность в экспериментально фрагментированном ландшафте: модели сообщества в пространстве и времени». Министерство сельского хозяйства США. Получено 2013-09-30.
- ^ Баниски, Сэнди (3 июля 1995 г.). «Наводнения меняют облик парка Шенандоа». Балтимор Сан. Получено 2019-07-05.
- ^ Майкл Дж. Барбур и Уильям Дуайт Биллингс (2000) Наземная растительность Северной Америки, Cambridge University Press, 708 стр. ISBN 0-521-55986-3, ISBN 978-0-521-55986-7
- ^ Баззаз, Ф.А. (1996). Растения в изменяющейся среде. Издательство Кембриджского университета. п. 4. ISBN 9-780521-398435.
- ^ Дини-Андреоте, Франсиско; Стеген, Джеймс; Дирк ван Эльзас, Ян; Фалькао Саллес, Жоана (17 марта 2015 г.). «Механизмы распутывания, которые обеспечивают баланс между стохастическими и детерминированными процессами в микробной последовательности». PNAS. 112 (11): E1326 – E1332. Дои:10.1073 / pnas.1414261112. ЧВК 4371938. PMID 25733885.
- ^ Макэвой, Том, Положительное влияние на лесное хозяйство, стр. 32 «Преемственность видов и толерантность», Island Press, 2004 г.
дальнейшее чтение
- Коннелл, Дж. Х.; Р. О. Слатьер (1977). «Механизмы сукцессии в естественных сообществах и их роль в стабильности и организации сообществ». Американский натуралист. 111 (982): 1119–44. Дои:10.1086/283241. S2CID 3587878.
внешняя ссылка
- Научная помощь: преемственность Объяснение преемственности для старшеклассников.
- Биографический очерк Генри Чендлера Коулза.
- Робберт Мерфи видит в значительной степени идеологическую, а не научную основу для неблагосклонного отношения к преемственности со стороны нынешней экологической ортодоксии и пытается восстановить преемственность с помощью целостных и телеологических аргументов.
- https://biologydictionary.net/ecological-succession/
| Авторитетный контроль |
|---|