РНК - RNA

Рибонуклеиновая кислота (РНК) это полимерный молекула, необходимая для различных биологических ролей в кодирование, расшифровка, регулирование и выражение из гены. РНК и ДНК находятся нуклеиновые кислоты. Вместе с липиды, белки, и углеводы, нуклеиновые кислоты составляют одну из четырех основных макромолекулы необходим для всех известных форм жизнь. Подобно ДНК, РНК собирается в виде цепочки нуклеотиды, но в отличие от ДНК, РНК встречается в природе в виде одинарной цепи, свернутой сама по себе, а не в виде парной двойной цепи. Использование клеточных организмов информационная РНК (мРНК) для передачи генетической информации (используя азотистые основания из гуанин, урацил, аденин, и цитозин, обозначаемые буквами G, U, A и C), который направляет синтез определенных белков. Много вирусы кодируют свою генетическую информацию с помощью РНК геном.
Некоторые молекулы РНК играют активную роль в клетках, катализируя биологические реакции, контролируя экспрессия гена, или восприятие и передача ответов на клеточные сигналы. Одним из таких активных процессов является синтез белка, универсальная функция, в которой молекулы РНК направляют синтез белков на рибосомы. Этот процесс использует переносить РНК (тРНК) молекул для доставки аминокислоты к рибосоме, где рибосомная РНК (рРНК) затем связывает аминокислоты вместе с образованием кодированных белков.
Сравнение с ДНК

Подобно ДНК, большинство биологически активных РНК, в том числе мРНК, тРНК, рРНК, мяРНК, и другие некодирующие РНК, содержат самокомплементарные последовательности, которые позволяют частям РНК складываться[5] и соединяются с собой, чтобы сформировать двойные спирали. Анализ этих РНК показал, что они высоко структурированы. В отличие от ДНК, их структуры не состоят из длинных двойных спиралей, а скорее представляют собой совокупность коротких спиралей, упакованных вместе в структуры, похожие на белки.
Таким образом, РНК могут достичь химического катализ (как ферменты).[6] Например, определение структуры рибосомы - комплекса РНК-белок, катализирующего образование пептидной связи - показало, что ее активный центр полностью состоит из РНК.[7]
Структура
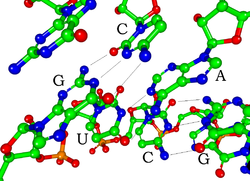
Каждый нуклеотид в РНК содержит рибоза сахар, с номерами атомов углерода от 1 до 5. База прикреплена к позиции 1 ', как правило, аденин (А), цитозин (С), гуанин (G), или урацил (U). Аденин и гуанин являются пурины, цитозин и урацил являются пиримидины. А фосфат группа прикреплена к позиции 3 'одной рибозы и позиции 5' следующей. Каждая фосфатная группа имеет отрицательный заряд, что делает РНК заряженной молекулой (полианионом). Базы формы водородные связи между цитозином и гуанином, между аденином и урацилом и между гуанином и урацилом.[8] Однако возможны и другие взаимодействия, такие как связывание группы оснований аденина друг с другом в выпуклости,[9]или GNRA тетрапетля который имеет пару оснований гуанин-аденин.[8]

Важным структурным компонентом РНК, который отличает ее от ДНК, является наличие гидроксил группа в положении 2 'сахара рибозы. Наличие этой функциональной группы заставляет спираль в основном принимать А-образная геометрия,[10] хотя в контексте однонитевых динуклеотидов РНК редко может также принимать B-форму, наиболее часто наблюдаемую в ДНК.[11] Геометрия А-образной формы дает очень глубокую и узкую большую канавку и неглубокую и широкую малую канавку.[12] Второе следствие присутствия 2'-гидроксильной группы состоит в том, что в конформационно гибких областях молекулы РНК (то есть, не участвующих в образовании двойной спирали) она может химически атаковать соседнюю фосфодиэфирную связь, чтобы расщепить основную цепь.[13]
РНК транскрибируется всего четырьмя основаниями (аденином, цитозином, гуанином и урацилом),[14] но эти основания и присоединенные сахара могут быть модифицированы различными способами по мере созревания РНК. Псевдоуридин (Ψ), в котором связь между урацилом и рибозой изменена с связи C – N на связь C – C, и риботимидин (T) находятся в разных местах (наиболее заметные из них находятся в петле TΨC тРНК ).[15] Другое известное модифицированное основание - это гипоксантин, дезаминированное основание аденина, которое нуклеозид называется инозин (Я). Инозин играет ключевую роль в гипотеза колебания из генетический код.[16]
Существует более 100 других модифицированных нуклеозидов природного происхождения.[17] Наибольшее конструктивное разнообразие модификаций можно найти в тРНК,[18] а псевдоуридин и нуклеозиды с 2'-O-метилрибоза часто присутствующие в рРНК являются наиболее распространенными.[19] Конкретные роли многих из этих модификаций в РНК до конца не изучены. Однако примечательно, что в рибосомной РНК многие посттранскрипционные модификации происходят в высокофункциональных областях, таких как центр пептидилтрансферазы и интерфейс субъединицы, подразумевая, что они важны для нормального функционирования.[20]
Функциональная форма одноцепочечных молекул РНК, как и белки, часто требует определенного третичная структура. Строительные леса для этой конструкции предоставлены вторичный структурный элементы, которые являются водородными связями внутри молекулы. Это приводит к нескольким узнаваемым «доменам» вторичной структуры, таким как петли для шпилек, выпуклости и внутренние петли.[21] Поскольку РНК заряжена, ионы металлов, такие как Mg2+ необходимы для стабилизации многих вторичных и третичные структуры.[22]
Встречающиеся в природе энантиомер РНК D-РНК состоит из D-рибонуклеотиды. Все центры хиральности расположены в D-рибоза. Используя L-рибоза а точнее L-рибонуклеотиды, L-РНК может быть синтезирована. L-РНК намного более устойчива против деградации РНКаза.[23]
Подобно другим структурированным биополимерам, таким как белки, можно определить топологию свернутой молекулы РНК. Это часто делается на основе расположения внутрицепочечных контактов внутри свернутой РНК, что называется топология схемы.
Синтез
Синтез РНК обычно катализируется ферментом -РНК-полимераза - использование ДНК в качестве матрицы, процесс, известный как транскрипция. Инициирование транскрипции начинается со связывания фермента с промоутер последовательность в ДНК (обычно находится «перед геном»). Двойная спираль ДНК разматывается геликаза активность фермента. Затем фермент продвигается вдоль цепочки матрицы в направлении от 3 ’к 5’, синтезируя комплементарную молекулу РНК с удлинением в направлении от 5 ’к 3’. Последовательность ДНК также определяет, где произойдет остановка синтеза РНК.[24]
Первичная стенограмма РНК часто модифицированный ферментами после транскрипции. Например, поли (А) хвост и Крышка 5 футов добавляются к эукариотическим пре-мРНК и интроны удаляются сплайсосома.
Также есть ряд РНК-зависимые РНК-полимеразы которые используют РНК в качестве матрицы для синтеза новой цепи РНК. Например, ряд РНК-вирусов (таких как полиовирус) используют этот тип фермента для репликации своего генетического материала.[25] Кроме того, РНК-зависимая РНК-полимераза является частью РНК-интерференция путь во многих организмах.[26]
Типы РНК
Обзор

Информационная РНК (мРНК) - это РНК, передающая информацию от ДНК к рибосома, сайты синтеза белка (перевод ) в ячейке. Кодирующая последовательность мРНК определяет аминокислота последовательность в белок что производится.[27] Однако многие РНК не кодируют белок (у эукариот около 97% транскрипционной продукции не кодирует белок.[28][29][30][31]).
Эти так называемые некодирующие РНК («нкРНК») могут кодироваться собственными генами (генами РНК), но также могут происходить из мРНК интроны.[32] Наиболее яркими примерами некодирующих РНК являются: переносить РНК (тРНК) и рибосомная РНК (рРНК), обе из которых участвуют в процессе трансляции.[4] Есть также некодирующие РНК, участвующие в регуляции генов, Обработка РНК и другие роли. Некоторые РНК способны катализировать химические реакции, такие как резка и перевязка другие молекулы РНК,[33] и катализ пептидная связь формирование в рибосома;[7] они известны как рибозимы.
В длину
По длине цепи РНК в РНК входят малая РНК и длинная РНК.[34] Обычно, малые РНК короче 200нт в длину, а длинные РНК более 200нт длинный.[35] Длинные РНК, также называемые большими РНК, в основном включают: длинная некодирующая РНК (днРНК) и мРНК. Малые РНК в основном включают 5.8S рибосомная РНК (рРНК), 5S рРНК, переносить РНК (тРНК), микроРНК (миРНК), малая интерферирующая РНК (миРНК), малая ядрышковая РНК (мяРНК), Piwi-взаимодействующая РНК (пиРНК), малая РНК на основе тРНК (цРНК)[36] и малая РНК, происходящая от рДНК (srRNA).[37]Есть определенные исключения, как в случае с 5S рРНК членов рода Галококк (Археи ), которые имеют вставку, увеличивая ее размер.[38][39][40]
В переводе
Посланник РНК (мРНК) несет информацию о последовательности белка в рибосомы, фабрики синтеза белка в клетке. это закодированный так что каждые три нуклеотида (a кодон ) соответствует одной аминокислоте. В эукариотический клетки, как только мРНК-предшественник (пре-мРНК) была транскрибирована с ДНК, она процессируется до зрелой мРНК. Это устраняет его интроны - некодирующие участки пре-мРНК. Затем мРНК экспортируется из ядра в цитоплазму, где она связывается с рибосомами и переведено в соответствующую белковую форму с помощью тРНК. В прокариотических клетках, которые не имеют отсеков ядра и цитоплазмы, мРНК может связываться с рибосомами, пока она транскрибируется с ДНК. Через некоторое время сообщение распадается на составляющие его нуклеотиды с помощью рибонуклеазы.[27]
Передача РНК (тРНК) представляет собой небольшую цепь РНК длиной около 80 нуклеотиды который передает определенную аминокислоту растущему полипептид цепь на рибосомном сайте синтеза белка во время трансляции. Он имеет участки для присоединения аминокислот и антикодон регион для кодон распознавание, которое связывается с определенной последовательностью в цепи информационной РНК посредством водородных связей.[32]
Рибосомная РНК (рРНК) является каталитическим компонентом рибосом. Рибосомы эукариот содержат четыре различных молекулы рРНК: 18S, 5,8S, 28S и 5S рРНК. Три молекулы рРНК синтезируются в ядрышко, и один синтезируется в другом месте. В цитоплазме рибосомная РНК и белок объединяются, образуя нуклеопротеин, называемый рибосомой. Рибосома связывает мРНК и осуществляет синтез белка. Несколько рибосом могут быть присоединены к одной мРНК в любое время.[27] Почти вся РНК, обнаруженная в типичной эукариотической клетке, является рРНК.
Трансфер-мессенджер РНК (тмРНК) встречается во многих бактерии и пластиды. Он маркирует белки, кодируемые мРНК, у которых отсутствуют стоп-кодоны для деградации, и предотвращает остановку рибосомы.[41]
Регуляторная РНК
Самые ранние известные регуляторы экспрессия гена были белки, известные как репрессоры и активаторы, регуляторы со специфическими короткими сайтами связывания внутри усилитель регионы рядом с регулируемыми генами.[42] Совсем недавно было обнаружено, что РНК также регулируют гены. У эукариот существует несколько видов РНК-зависимых процессов, регулирующих экспрессию генов в различных точках, например: РНКи подавляющие гены посттранскрипция союзник, длинные некодирующие РНК отключение блоков хроматин эпигенетически, и энхансерные РНК индуцирование повышенной экспрессии генов.[43] В дополнение к этим механизмам у эукариот как бактерии, так и археи, как было обнаружено, широко используют регуляторные РНК. Бактериальная малая РНК и CRISPR system являются примерами таких систем прокариотической регуляторной РНК.[44] Fire и Mello были награждены премией 2006 года. Нобелевская премия по физиологии и медицине для открытия микроРНК (miRNA), специфические короткие молекулы РНК, которые могут образовывать пары оснований с мРНК.[45]
РНК-интерференция миРНК
Уровни посттранскрипционной экспрессии многих генов можно контролировать с помощью РНК-интерференция, в котором миРНК, специфические короткие молекулы РНК, соединяются с участками мРНК и направляют их для деградации.[46] Этот антисмысловой -основанный процесс включает шаги, которые сначала обрабатывают РНК, чтобы она могла пара оснований с областью его целевой мРНК. Как только происходит спаривание оснований, другие белки направляют мРНК на разрушение нуклеазы.[43] Огонь и Мелло были награждены премией 2006 г. Нобелевская премия по физиологии и медицине за это открытие.[45]
Длинные некодирующие РНК
Далее, чтобы связать с регулированием, были Xist и другие длинные некодирующие РНК связана с Инактивация Х-хромосомы. Их роли, поначалу загадочные, показали Джинни Т. Ли и другие, чтобы быть заглушить блоков хроматина за счет привлечения Поликомб комплекс так, чтобы информационная РНК не могла быть транскрибирована с них.[47] Дополнительные днРНК, которые в настоящее время определяются как РНК из более чем 200 пар оснований, которые, по-видимому, не обладают кодирующим потенциалом,[48] были обнаружены связанные с регулированием стволовая клетка плюрипотентность и деление клеток.[48]
Энхансерные РНК
Третья основная группа регуляторных РНК называется энхансерные РНК.[48] В настоящее время неясно, являются ли они уникальной категорией РНК разной длины или представляют собой отдельное подмножество днРНК. Во всяком случае, они переписаны с усилители, которые представляют собой известные регуляторные сайты в ДНК рядом с регулируемыми ими генами.[48][49] Они активируют транскрипцию гена (ов) под контролем энхансера, с которого они транскрибируются.[48][50]
Регуляторная РНК у прокариот
Сначала считалось, что регуляторная РНК - это эукариотический феномен, часть объяснения того, почему у высших организмов наблюдается гораздо больше транскрипции, чем предполагалось. Но как только исследователи начали искать возможные регуляторы РНК в бактериях, они обнаружили и там, что называется малая РНК (мРНК).[51][44] В настоящее время повсеместная природа систем регуляции РНК генов обсуждается как подтверждение Мир РНК теория.[43][52] Бактериальные малые РНК обычно действуют через антисмысловой спаривание с мРНК для подавления ее трансляции, либо влияя на стабильность, либо влияя на способность цис-связывания.[43] Рибопереключатели также были обнаружены. Это цис-действующие регуляторные последовательности РНК, действующие аллостерически. Они меняют форму, когда связываются метаболиты так что они приобретают или теряют способность связывать хроматин для регулирования экспрессии генов.[53][54]
У архей также есть системы регуляторных РНК.[55] Система CRISPR, недавно использовавшаяся для редактирования ДНК на месте, действует через регуляторные РНК у архей и бактерий, обеспечивая защиту от вирусных захватчиков.[43][56]
При обработке РНК

Многие РНК участвуют в модификации других РНК.Интроны находятся сращенный снаружи пре-мРНК к сплайсосомы, которые содержат несколько малые ядерные РНК (мяРНК),[4] или интроны могут быть рибозимами, которые сплайсируются сами по себе.[57]РНК также можно изменить, модифицировав ее нуклеотиды на нуклеотиды, отличные от А, C, грамм и U У эукариот модификации нуклеотидов РНК, как правило, направляются малые ядрышковые РНК (мяРНК; 60–300 н.),[32] найдено в ядрышко и кахальные тела. snoRNA связываются с ферментами и направляют их к месту на РНК путем спаривания оснований с этой РНК. Затем эти ферменты выполняют модификацию нуклеотидов. рРНК и тРНК сильно модифицируются, но мяРНК и мРНК также могут быть мишенью для модификации оснований.[58][59] РНК также может быть метилирована.[60][61]
РНК геномы
Подобно ДНК, РНК может нести генетическую информацию. РНК-вирусы имеют геномы состоит из РНК, кодирующей ряд белков. Вирусный геном реплицируется некоторыми из этих белков, в то время как другие белки защищают геном, когда вирусная частица перемещается к новой клетке-хозяину. Вироиды являются еще одной группой патогенов, но они состоят только из РНК, не кодируют какой-либо белок и реплицируются полимеразой клетки растения-хозяина.[62]
В обратной транскрипции
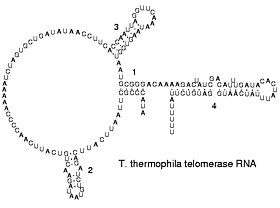
Вирусы с обратной транскрипцией реплицируют свои геномы с помощью обратная расшифровка Копии ДНК с их РНК; эти копии ДНК затем транскрибируются в новую РНК. Ретротранспозоны также распространяются путем копирования ДНК и РНК друг с друга,[63] и теломераза содержит РНК, которая используется в качестве шаблона для построения концов хромосом эукариот.[64]
Двухцепочечная РНК

Двухцепочечная РНК (дцРНК) - это РНК с двумя комплементарными цепями, подобная ДНК, обнаруженной во всех клетках, но с заменой тимина на урацил. дцРНК образует генетический материал некоторых вирусы (двухцепочечные РНК-вирусы ). Двухцепочечная РНК, такая как вирусная РНК или миРНК, может вызвать РНК-интерференция в эукариоты, а также интерферон ответ в позвоночные.[65][66][67][68]
Круговая РНК
В конце 1970-х годов было показано, что существует однониточная ковалентно замкнутая, то есть кольцевая форма РНК, экспрессируемая во всем царстве животных и растений (см. circRNA ).[69] Считается, что circRNA возникают в результате реакции "обратного сплайсинга", когда сплайсосома присоединяет нижележащий донор к вышестоящему акцепторному сайту сплайсинга. До сих пор функция circRNAs в значительной степени неизвестна, хотя на нескольких примерах была продемонстрирована губчатая активность микроРНК.
Ключевые открытия в биологии РНК

Исследования РНК привели ко многим важным биологическим открытиям и многочисленным Нобелевским премиям. Нуклеиновые кислоты были открыты в 1868 г. Фридрих Мишер, который назвал материал нуклеином, поскольку он был обнаружен в ядро.[70] Позже было обнаружено, что прокариотические клетки, не имеющие ядра, также содержат нуклеиновые кислоты. О роли РНК в синтезе белка подозревали еще в 1939 году.[71] Северо-Очоа выиграл 1959 г. Нобелевская премия по медицине (поделился с Артур Корнберг ) после того, как он обнаружил фермент, который может синтезировать РНК в лаборатории.[72] Однако фермент, открытый Очоа (полинуклеотидфосфорилаза Позже было показано, что он отвечает за деградацию РНК, а не за синтез РНК. В 1956 году Алекс Рич и Дэвид Дэвис гибридизировали две отдельные цепи РНК, чтобы сформировать первый кристалл РНК, структуру которого можно было определить с помощью рентгеновской кристаллографии.[73]
Последовательность из 77 нуклеотидов тРНК дрожжей была обнаружена Роберт В. Холли в 1965 г.,[74] выиграв Холли Нобелевская премия по медицине 1968 года (поделился с Хар Гобинд Кхорана и Маршалл Ниренберг ).
В начале 1970-х гг. ретровирусы и обратная транскриптаза были открыты, впервые продемонстрировав, что ферменты могут копировать РНК в ДНК (противоположность обычному пути передачи генетической информации). Для этой работы Дэвид Балтимор, Ренато Дульбекко и Говард Темин были удостоены Нобелевской премии в 1975 г. В 1976 г. Уолтер Фирс и его команда определили первую полную нуклеотидную последовательность генома РНК-вируса, бактериофаг MS2.[75]
В 1977 г. интроны и Сплайсинг РНК были обнаружены как в вирусах млекопитающих, так и в клеточных генах, в результате чего Нобелевская премия 1993 г. Филип Шарп и Ричард Робертс.Каталитические молекулы РНК (рибозимы ) были обнаружены в начале 1980-х, что привело к присуждению Нобелевской премии 1989 г. Томас Чех и Сидни Альтман. В 1990 году его нашли в Петуния что введенные гены могут заставить замолчать аналогичные гены самого растения, что, как теперь известно, является результатом РНК-интерференция.[76][77]
Примерно в то же время РНК длиной 22 н. микроРНК, было обнаружено, что они играют роль в разработка из C. elegans.[78]Исследования интерференции РНК были удостоены Нобелевской премии. Эндрю Файер и Крейг Мелло в 2006 г., а еще одна Нобелевская премия была присуждена за исследования транскрипции РНК в Роджер Корнберг в том же году. Открытие регуляторных РНК генов привело к попыткам разработать препараты на основе РНК, такие как миРНК, чтобы замолчать гены.[79] В дополнение к Нобелевской премии, присужденной за исследования РНК в 2009 году, она была присуждена Венки Рамакришнану, Тому Стейтцу и Аде Йонат за выяснение атомной структуры рибосомы.
Актуальность для пребиотической химии и абиогенеза
В 1968 г. Карл Вёзе предположил, что РНК может быть катализатором, и предположил, что самые ранние формы жизни (самовоспроизводящиеся молекулы) могли полагаться на РНК как для передачи генетической информации, так и для катализирования биохимических реакций - Мир РНК.[80][81]
В марте 2015 г. ДНК и РНК нуклеотиды, включая урацил, цитозин и тимин, как сообщается, были сформированы в лаборатории под космическое пространство условий, используя стартовые химикаты, такие как пиримидин, органическое соединение обычно встречается в метеориты. Пиримидин, как полициклические ароматические углеводороды (ПАУ), является одним из самых богатых углеродом соединений, обнаруженных в Вселенная и, возможно, были сформированы в красные гиганты или в межзвездная пыль и газовые облака.[82]
Смотрите также
Рекомендации
- ^ «РНК: универсальная молекула». Университет Юты. 2015.
- ^ «Нуклеотиды и нуклеиновые кислоты» (PDF). Калифорнийский университет в Лос-Анджелесе. Архивировано из оригинал (PDF) на 2015-09-23. Получено 2015-08-26.
- ^ Шукла Р.Н. (2014). Анализ хромосом. ISBN 978-93-84568-17-7.
- ^ а б c Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). Биохимия (5-е изд.). WH Freeman and Company. С. 118–19, 781–808. ISBN 978-0-7167-4684-3. OCLC 179705944.
- ^ Tinoco I, Bustamante C (октябрь 1999 г.). «Как складывается РНК». Журнал молекулярной биологии. 293 (2): 271–81. Дои:10.1006 / jmbi.1999.3001. PMID 10550208.
- ^ Хиггс П.Г. (август 2000 г.). «Вторичная структура РНК: физические и вычислительные аспекты». Ежеквартальные обзоры биофизики. 33 (3): 199–253. Дои:10.1017 / S0033583500003620. PMID 11191843.
- ^ а б Ниссен П., Хансен Дж., Бан Н., Мур ПБ, Стейтц Т.А. (август 2000 г.). «Структурные основы активности рибосом в синтезе пептидных связей». Наука. 289 (5481): 920–30. Bibcode:2000Sci ... 289..920N. Дои:10.1126 / science.289.5481.920. PMID 10937990.
- ^ а б Ли Дж. К., Гутелл Р. Р. (декабрь 2004 г.). «Разнообразие конформаций пар оснований и их появление в структуре рРНК и структурных мотивах РНК». Журнал молекулярной биологии. 344 (5): 1225–49. Дои:10.1016 / j.jmb.2004.09.072. PMID 15561141.
- ^ Барцишевский Дж, Фредерик Б., Кларк С. (1999). Биохимия и биотехнология РНК. Springer. С. 73–87. ISBN 978-0-7923-5862-6. OCLC 52403776.
- ^ Салазар М., Федоров О.Ю., Миллер Дж. М., Рибейро Н.С., Рейд Б.Р. (апрель 1993 г.). «Нить ДНК в гибридных дуплексах DNA.RNA не является ни B-формой, ни A-формой в растворе». Биохимия. 32 (16): 4207–15. Дои:10.1021 / bi00067a007. PMID 7682844.
- ^ Седова А., Банавали Н.К. (февраль 2016 г.). «РНК приближается к B-форме в контекстах сложенных одноцепочечных динуклеотидов». Биополимеры. 105 (2): 65–82. Дои:10.1002 / bip.22750. PMID 26443416. S2CID 35949700.
- ^ Герман Т., Патель DJ (март 2000 г.). «РНК выступает как архитектурный мотив и мотив узнавания». Структура. 8 (3): R47–54. Дои:10.1016 / S0969-2126 (00) 00110-6. PMID 10745015.
- ^ Миккола С., Стенман Э., Нурми К., Юсефи-Салакде Э., Стрёмберг Р., Лённберг Х. (1999). «Механизм расщепления фосфодиэфирных связей РНК, стимулируемого ионом металла, включает в себя общий кислотный катализ акво-ионом металла при уходе уходящей группы». Журнал химического общества, Perkin Transactions 2 (8): 1619–26. Дои:10.1039 / a903691a.
- ^ Янковский Я.А., Полак Я.М. (1996). Клинический анализ генов и манипуляции с ними: инструменты, методы и устранение неполадок. Издательство Кембриджского университета. п.14. ISBN 978-0-521-47896-0. OCLC 33838261.
- ^ Yu Q, Morrow CD (май 2001 г.). «Идентификация критических элементов в акцепторном стволе тРНК и петле Т (Psi) C, необходимых для инфекционности вируса иммунодефицита человека типа 1». Журнал вирусологии. 75 (10): 4902–6. Дои:10.1128 / JVI.75.10.4902-4906.2001. ЧВК 114245. PMID 11312362.
- ^ Эллиотт М.С., Трюин Р.В. (февраль 1984 г.). «Биосинтез инозина в транспортной РНК ферментативной вставкой гипоксантина». Журнал биологической химии. 259 (4): 2407–10. PMID 6365911.
- ^ Кантара В.А., Крейн П.Ф., Розенски Дж., Макклоски Дж. А., Харрис К.А., Чжан Х, Вендейш Ф.А., Фабрис Д., Агрис П.Ф. (январь 2011 г.). «База данных модификаций РНК, RNAMDB: обновление 2011 г.». Исследования нуклеиновых кислот. 39 (Выпуск базы данных): D195-201. Дои:10.1093 / nar / gkq1028. ЧВК 3013656. PMID 21071406.
- ^ Söll D, RajBhandary U (1995). TRNA: структура, биосинтез и функция. ASM Press. п. 165. ISBN 978-1-55581-073-3. OCLC 183036381.
- ^ Поцелуй Т (июль 2001 г.). «Небольшая ядрышковая РНК-управляемая посттранскрипционная модификация клеточных РНК». Журнал EMBO. 20 (14): 3617–22. Дои:10.1093 / emboj / 20.14.3617. ЧВК 125535. PMID 11447102.
- ^ Король Т.Х., Лю Б., Маккалли Р.Р., Фурнье MJ (февраль 2003 г.). «Структура и активность рибосом изменены в клетках, лишенных snoRNP, которые образуют псевдоуридины в центре пептидилтрансферазы». Молекулярная клетка. 11 (2): 425–35. Дои:10.1016 / S1097-2765 (03) 00040-6. PMID 12620230.
- ^ Мэтьюз Д.Х., Дисней Мэриленд, Чайлдс Дж.Л., Шредер С.Дж., Цукер М., Тернер Д.Х. (май 2004 г.). «Включение ограничений химической модификации в алгоритм динамического программирования для прогнозирования вторичной структуры РНК». Труды Национальной академии наук Соединенных Штатов Америки. 101 (19): 7287–92. Bibcode:2004ПНАС..101.7287М. Дои:10.1073 / pnas.0401799101. ЧВК 409911. PMID 15123812.
- ^ Тан З.Д., Чен С.Дж. (июль 2008 г.). «Солевая зависимость стабильности шпильки нуклеиновой кислоты». Биофизический журнал. 95 (2): 738–52. Bibcode:2008BpJ .... 95..738T. Дои:10.1529 / biophysj.108.131524. ЧВК 2440479. PMID 18424500.
- ^ Фатер А., Клуссманн С. (январь 2015 г.). «Превращение зеркальных олигонуклеотидов в лекарства: эволюция терапии Spiegelmer (®)». Открытие наркотиков сегодня. 20 (1): 147–55. Дои:10.1016 / j.drudis.2014.09.004. PMID 25236655.
- ^ Нудлер Э., Готтесман М.Э. (август 2002 г.). «Терминация транскрипции и анти-терминация в E. coli». Гены в клетки. 7 (8): 755–68. Дои:10.1046 / j.1365-2443.2002.00563.x. PMID 12167155. S2CID 23191624.
- ^ Хансен Дж. Л., Лонг А. М., Шульц СК (август 1997 г.). «Структура РНК-зависимой РНК-полимеразы полиовируса». Структура. 5 (8): 1109–22. Дои:10.1016 / S0969-2126 (97) 00261-X. PMID 9309225.
- ^ Алквист П. (май 2002 г.). «РНК-зависимые РНК-полимеразы, вирусы и сайленсинг РНК». Наука. 296 (5571): 1270–73. Bibcode:2002Наука ... 296.1270A. Дои:10.1126 / science.1069132. PMID 12016304. S2CID 42526536.
- ^ а б c Купер Г.К., Хаусман Р.Е. (2004). Клетка: молекулярный подход (3-е изд.). Синауэр. С. 261–76, 297, 339–44. ISBN 978-0-87893-214-6. OCLC 174924833.
- ^ Мэттик Дж. С., Гаген М. Дж. (Сентябрь 2001 г.). «Эволюция управляемых многозадачных генных сетей: роль интронов и других некодирующих РНК в развитии сложных организмов». Молекулярная биология и эволюция. 18 (9): 1611–30. Дои:10.1093 / oxfordjournals.molbev.a003951. PMID 11504843.
- ^ Mattick JS (ноябрь 2001 г.). «Некодирующие РНК: архитекторы эукариотической сложности». EMBO отчеты. 2 (11): 986–91. Дои:10.1093 / embo-reports / kve230. ЧВК 1084129. PMID 11713189.
- ^ Маттик JS (октябрь 2003 г.). «Бросить вызов догме: скрытый слой небелковых РНК в сложных организмах» (PDF). BioEssays. 25 (10): 930–39. CiteSeerX 10.1.1.476.7561. Дои:10.1002 / bies.10332. PMID 14505360. Архивировано из оригинал (PDF) на 2009-03-06.
- ^ Мэттик Дж. С. (октябрь 2004 г.). «Скрытая генетическая программа сложных организмов». Scientific American. 291 (4): 60–67. Bibcode:2004SciAm.291d..60M. Дои:10.1038 / Scientificamerican1004-60. PMID 15487671.[мертвая ссылка ]
- ^ а б c Вирта В. (2006). Майнинг транскриптома - методы и приложения. Стокгольм: Школа биотехнологии Королевского технологического института. ISBN 978-91-7178-436-0. OCLC 185406288.
- ^ Росси Дж. Дж. (Июль 2004 г.). «Рибозимная диагностика достигает совершеннолетия». Химия и биология. 11 (7): 894–95. Дои:10.1016 / j.chembiol.2004.07.002. PMID 15271347.
- ^ Сторц G (май 2002 г.). «Расширяющаяся вселенная некодирующих РНК». Наука. 296 (5571): 1260–63. Bibcode:2002Наука ... 296.1260S. Дои:10.1126 / science.1072249. PMID 12016301. S2CID 35295924.
- ^ Fatica A, Bozzoni I (январь 2014 г.). «Длинные некодирующие РНК: новые игроки в дифференцировке и развитии клеток». Природа Обзоры Генетика. 15 (1): 7–21. Дои:10.1038 / nrg3606. PMID 24296535. S2CID 12295847.[постоянная мертвая ссылка ]
- ^ Chen Q, Yan M, Cao Z, Li X, Zhang Y, Shi J и др. (Январь 2016 г.). «ЦРНК сперматозоидов способствуют наследованию приобретенных метаболических нарушений из поколения в поколение» (PDF). Наука. 351 (6271): 397–400. Bibcode:2016Научный ... 351..397C. Дои:10.1126 / science.aad7977. PMID 26721680. S2CID 21738301.
- ^ Вэй Х, Чжоу Б., Чжан Ф, Ту И, Ху И, Чжан Б., Чжай К. (2013). «Профилирование и идентификация малых РНК, полученных из рДНК, и их потенциальных биологических функций». PLOS ONE. 8 (2): e56842. Bibcode:2013PLoSO ... 856842W. Дои:10.1371 / journal.pone.0056842. ЧВК 3572043. PMID 23418607.
- ^ Люерсен К.Р., Николсон Д.Е., Юбэнкс, округ Колумбия, Fox GE (1981). «Архебактериальная 5S рРНК содержит длинную инсерционную последовательность». Природа. 293 (Pt 12): 755–756. Дои:10.1099/00221287-145-12-3565. PMID 6169998.
- ^ Стэн-Лоттер Х, Макдженити Т.Дж., Легат А, Деннер Э.Б., Глейзер К., Стеттер К.О., Ваннер Г. (1999). «Очень похожие штаммы Halococcus salifodinae обнаружены в географически разделенных месторождениях пермотриасовой соли». Микробиология. 145 (Pt 12): 3565–3574. Дои:10.1099/00221287-145-12-3565. PMID 10627054.
- ^ Тирумалай MR, Kaelber JT, Park DR, Tran Q, Fox GE (август 2020 г.). «Визуализация с помощью криоэлектронной микроскопии большой вставки в 5S рибосомной РНК чрезвычайно галофильного архея» Halococcus morrhuae". FEBS Open Bio. 10 (10): 1938–1946. Дои:10.1002/2211-5463.12962. ЧВК 7530397. PMID 32865340.
- ^ Гено де Новоа П., Уильямс КП (январь 2004 г.). «Сайт тмРНК: редуктивная эволюция тмРНК в пластидах и других эндосимбионтах». Исследования нуклеиновых кислот. 32 (Выпуск базы данных): D104–08. Дои:10.1093 / нар / гх102. ЧВК 308836. PMID 14681369.
- ^ Джейкоб Ф, Монод Дж (1961). «Генетические регуляторные механизмы в синтезе белков». Журнал молекулярной биологии. 3 (3): 318–56. Дои:10.1016 / с0022-2836 (61) 80072-7. PMID 13718526.
- ^ а б c d е Моррис К., Мэттик Дж. (2014). «Рост регуляторной РНК». Природа Обзоры Генетика. 15 (6): 423–37. Дои:10.1038 / nrg3722. ЧВК 4314111. PMID 24776770.
- ^ а б Готтесман С (2005). «Микроорганизмы для микробов: некодирующие регуляторные РНК в бактериях». Тенденции в генетике. 21 (7): 399–404. Дои:10.1016 / j.tig.2005.05.008. PMID 15913835.
- ^ а б «Нобелевская премия по физиологии и медицине 2006 г.». Nobelprize.org. Nobel Media AB 2014. Интернет. 6 августа 2018 г. http://www.nobelprize.org/nobel_prizes/medicine/laureates/2006
- ^ Fire и др. (1998). «Мощное и специфическое генетическое вмешательство с помощью двухцепочечной РНК у Ceanorhabditis elegans». Природа. 391 (6669): 806–11. Bibcode:1998Натура.391..806F. Дои:10.1038/35888. PMID 9486653. S2CID 4355692.
- ^ Чжао Дж., Сун Б.К., Эрвин Дж. А., Сон Дж. Дж., Ли Дж. Т. (2008). «Белки Polycomb, нацеленные коротким повтором РНК на X-хромосому мыши». Наука. 322 (5902): 750–56. Bibcode:2008Sci ... 322..750Z. Дои:10.1126 / science.1163045. ЧВК 2748911. PMID 18974356.
- ^ а б c d е Ринн Дж. Л., Чанг Х. Ю. (2012). «Регуляция генома длинными некодирующими РНК». Анну. Преподобный Biochem. 81: 1–25. Дои:10.1146 / annurev-biochem-051410-092902. ЧВК 3858397. PMID 22663078.
- ^ Тафт Р. Дж., Каплан CD, Саймонс С., Мэттик Дж. С. (2009). «Эволюция, биогенез и функция промоторных РНК». Клеточный цикл. 8 (15): 2332–38. Дои:10.4161 / cc.8.15.9154. PMID 19597344.
- ^ Ором У.А., Дерриен Т., Берингер М., Гумиредди К., Гардини А. и др. (2010). "'Длинные некодирующие РНК с энхансер-подобной функцией в клетках человека ». Клетка. 143 (1): 46–58. Дои:10.1016 / j.cell.2010.09.001. ЧВК 4108080. PMID 20887892.
- ^ EGH Вагнер, П. Ромби. (2015). «Малые РНК у бактерий и архей: кто они, что они делают и как они это делают». Достижения в генетике (Том 90, стр. 133–208).
- ^ J.W. Нельсон, Р.Р. Брейкер (2017) «Утерянный язык мира РНК».Sci. Сигнал.10, eaam8812 1–11.
- ^ Винклеф WC (2005). «Рибопереключатели и роль некодирующих РНК в бактериальном метаболическом контроле». Curr. Мнение. Chem. Биол. 9 (6): 594–602. Дои:10.1016 / j.cbpa.2005.09.016. PMID 16226486.
- ^ Такер Б.Дж., Брейкер Р.Р. (2005). «Рибопереключатели как универсальные элементы контроля генов». Curr. Мнение. Struct. Биол. 15 (3): 342–48. Дои:10.1016 / j.sbi.2005.05.003. PMID 15919195.
- ^ Mojica FJ, Diez-Villasenor C, Soria E, Juez G (2000). "" «Биологическое значение семейства регулярно расположенных повторов в геномах архей, бактерий и митохондрий». Мол. Микробиол. 36 (1): 244–46. Дои:10.1046 / j.1365-2958.2000.01838.x. PMID 10760181. S2CID 22216574.
- ^ Браунс С., Джор М.М., Лундгрен М., Вестра Е., Слайкхейс Р., Снейдерс А., Дикман М., Макарова К., Кунин Е., Der Oost JV (2008). «Малые РНК CRISPR направляют противовирусную защиту прокариот». Наука. 321 (5891): 960–64. Bibcode:2008Sci ... 321..960B. Дои:10.1126 / science.1159689. ЧВК 5898235. PMID 18703739.
- ^ Steitz TA, Steitz JA (июль 1993 г.). «Общий двухметаллический ионный механизм каталитической РНК». Труды Национальной академии наук Соединенных Штатов Америки. 90 (14): 6498–502. Bibcode:1993ПНАС ... 90.6498С. Дои:10.1073 / пнас.90.14.6498. ЧВК 46959. PMID 8341661.
- ^ Се Дж, Чжан М., Чжоу Т., Хуа Х, Тан Л., Ву В. (январь 2007 г.). «Sno / scaRNAbase: курируемая база данных для малых ядрышковых РНК и РНК, специфичных для тельца кахаля». Исследования нуклеиновых кислот. 35 (Выпуск базы данных): D183–87. Дои:10.1093 / nar / gkl873. ЧВК 1669756. PMID 17099227.
- ^ Омер А.Д., Зиеше С., Декатур В.А., Фурнье М.Дж., Деннис П.П. (май 2003 г.). «Машины, модифицирующие РНК в архее». Молекулярная микробиология. 48 (3): 617–29. Дои:10.1046 / j.1365-2958.2003.03483.x. PMID 12694609. S2CID 20326977.
- ^ Кавай Дж., Николозо М., Bachellerie JP (октябрь 1996 г.). «Направленное метилирование рибозы РНК in vivo, направляемое специальными антисмысловыми руководствами по РНК». Природа. 383 (6602): 732–35. Bibcode:1996Натура.383..732C. Дои:10.1038 / 383732a0. PMID 8878486. S2CID 4334683.
- ^ Kiss-László Z, Henry Y, Bachellerie JP, Caizergues-Ferrer M, Kiss T (июнь 1996 г.). «Сайт-специфическое метилирование рибозы прерибосомальной РНК: новая функция для малых ядрышковых РНК». Клетка. 85 (7): 1077–88. Дои:10.1016 / S0092-8674 (00) 81308-2. PMID 8674114. S2CID 10418885.
- ^ Дарос Дж. А., Елена С. Ф., Флорес Р. (июнь 2006 г.). «Вироиды: нить Ариадны в лабиринте РНК». EMBO отчеты. 7 (6): 593–98. Дои:10.1038 / sj.embor.7400706. ЧВК 1479586. PMID 16741503.
- ^ Kalendar R, Vicient CM, Peleg O, Anamthawat-Jonsson K, Bolshoy A, Schulman AH (март 2004 г.). «Большие производные ретротранспозона: многочисленные, консервативные, но неавтономные ретроэлементы ячменя и родственных геномов». Генетика. 166 (3): 1437–50. Дои:10.1534 / genetics.166.3.1437. ЧВК 1470764. PMID 15082561.
- ^ Подлевский JD, Блей CJ, Omana RV, Qi X, Chen JJ (январь 2008 г.). «База данных теломеразы». Исследования нуклеиновых кислот. 36 (Выпуск базы данных): D339–43. Дои:10.1093 / нар / гкм700. ЧВК 2238860. PMID 18073191.
- ^ Блевинс Т., Раджешваран Р., Шивапрасад П.В., Бекназариантс Д., Си-Аммор А., Парк Х.С., Васкес Ф., Робертсон Д., Мейнс Ф., Хон Т., Пуггин М.М. (2006). «Четыре дайсера растений опосредуют биогенез вирусной малой РНК и подавление индуцированного ДНК вируса». Исследования нуклеиновых кислот. 34 (21): 6233–46. Дои:10.1093 / нар / gkl886. ЧВК 1669714. PMID 17090584.
- ^ Яна С., Чакраборти С., Нанди С., Деб Дж. К. (ноябрь 2004 г.). «РНК-интерференция: потенциальные терапевтические мишени». Прикладная микробиология и биотехнология. 65 (6): 649–57. Дои:10.1007 / s00253-004-1732-1. PMID 15372214. S2CID 20963666.
- ^ Шульц У., Касперс Б., Стахели П. (май 2004 г.). «Интерфероновая система позвоночных, не являющихся млекопитающими». Развитие и сравнительная иммунология. 28 (5): 499–508. Дои:10.1016 / j.dci.2003.09.009. PMID 15062646.
- ^ Уайтхед К.А., Дальман Дж. Э., Лангер Р.С., Андерсон Д.Г. (2011). «Молчание или стимуляция? SiRNA доставки и иммунной системы». Ежегодный обзор химической и биомолекулярной инженерии. 2: 77–96. Дои:10.1146 / annurev-chembioeng-061010-114133. PMID 22432611.
- ^ Hsu MT, Coca-Prados M (июль 1979 г.). «Электронно-микроскопические доказательства круговой формы РНК в цитоплазме эукариотических клеток». Природа. 280 (5720): 339–40. Bibcode:1979Натура.280..339H. Дои:10.1038 / 280339a0. PMID 460409. S2CID 19968869.
- ^ Dahm R (февраль 2005 г.). «Фридрих Мишер и открытие ДНК». Биология развития. 278 (2): 274–88. Дои:10.1016 / j.ydbio.2004.11.028. PMID 15680349.
- ^ Касперссон Т., Шульц Дж. (1939). «Пентозные нуклеотиды в цитоплазме растущих тканей». Природа. 143 (3623): 602–03. Bibcode:1939 г.Натура.143..602С. Дои:10.1038 / 143602c0. S2CID 4140563.
- ^ Очоа С (1959). «Ферментативный синтез рибонуклеиновой кислоты» (PDF). Нобелевская лекция.
- ^ Рич А., Дэвис Д. (1956). «Новая двухцепочечная спиральная структура: полиадениловая кислота и полиуридиловая кислота». Журнал Американского химического общества. 78 (14): 3548–49. Дои:10.1021 / ja01595a086.
- ^ Холли Р.В. и др. (Март 1965 г.). «Строение рибонуклеиновой кислоты». Наука. 147 (3664): 1462–65. Bibcode:1965Научный ... 147.1462H. Дои:10.1126 / science.147.3664.1462. PMID 14263761. S2CID 40989800.
- ^ Fiers W, et al. (Апрель 1976 г.). «Полная нуклеотидная последовательность РНК бактериофага MS2: первичная и вторичная структура гена репликазы». Природа. 260 (5551): 500–07. Bibcode:1976Натура.260..500F. Дои:10.1038 / 260500a0. PMID 1264203. S2CID 4289674.
- ^ Napoli C, Lemieux C, Jorgensen R (апрель 1990 г.). «Введение химерного гена халкон-синтазы в петунию приводит к обратимой совместной супрессии гомологичных генов в транс». Растительная клетка. 2 (4): 279–89. Дои:10.1105 / tpc.2.4.279. ЧВК 159885. PMID 12354959.
- ^ Дафни-Елин М., Чунг С.М., Франкман Э.Л., Цфира Т. (декабрь 2007 г.). «Векторы интерференции РНК pSAT: модульная серия для подавления множественных генов в растениях». Физиология растений. 145 (4): 1272–81. Дои:10.1104 / стр.107.106062. ЧВК 2151715. PMID 17766396.
- ^ Рувкун Г. (октябрь 2001 г.). «Молекулярная биология. Взгляд на крошечный мир РНК». Наука. 294 (5543): 797–99. Дои:10.1126 / science.1066315. PMID 11679654. S2CID 83506718.
- ^ Fichou Y, Férec C (декабрь 2006 г.). «Возможности олигонуклеотидов для терапевтического применения». Тенденции в биотехнологии. 24 (12): 563–70. Дои:10.1016 / j.tibtech.2006.10.003. PMID 17045686.
- ^ Зиберт С (2006). «Общие свойства структуры последовательностей и стабильные участки во вторичных структурах РНК» (PDF). Диссертация, Университет Альберта Людвига, Фрайбург-им-Брайсгау. п. 1. Архивировано из оригинал (PDF) 9 марта 2012 г.
- ^ Сатмари Э (июнь 1999 г.). «Происхождение генетического кода: аминокислоты как кофакторы в мире РНК». Тенденции в генетике. 15 (6): 223–29. Дои:10.1016 / S0168-9525 (99) 01730-8. PMID 10354582.
- ^ Марлер Р. (3 марта 2015 г.). «НАСА Эймс воспроизводит строительные блоки жизни в лаборатории». НАСА. Получено 5 марта 2015.
внешняя ссылка
- Сайт RNA World Коллекция ссылок (структуры, последовательности, инструменты, журналы)
- База данных нуклеиновых кислот Изображения ДНК, РНК и комплексов.
- Семинар Анны Мари Пайл: структура, функция и распознавание РНК
| Ключевые компоненты | |
|---|---|
| Поля | |
| Археогенетика из | |
| похожие темы | |
| Списки | |
| |
Виды нуклеиновые кислоты | |||||||
|---|---|---|---|---|---|---|---|
| Избиратели | |||||||
| Рибонуклеиновые кислоты (кодирование, некодирование ) |
| ||||||
| Дезоксирибонуклеиновые кислоты | |||||||
| Аналоги | |||||||
| Клонирование векторов | |||||||
| |||||||
| Авторитетный контроль |
|---|