Биология развития - Developmental biology
Биология развития это изучение процесса, посредством которого животные и растения расти и развиваться. Биология развития также включает в себя биологию регенерация, бесполое размножение, метаморфоза, а рост и дифференциация стволовые клетки во взрослом организме.
Перспективы
Основные процессы, задействованные в эмбриональное развитие животных: формирование рисунка ткани (через региональная спецификация и с рисунком дифференциация клеток ); рост тканей; и ткань морфогенез.
- Региональная спецификация относится к процессам, которые создают пространственный узор в шаре или листе изначально похожих ячеек. Обычно это связано с действием цитоплазматические детерминанты, расположенных в частях оплодотворенной яйцеклетки, и индуктивных сигналов, исходящих от сигнальных центров эмбриона. Ранние этапы региональная спецификация не генерируют функционально дифференцированные клетки, а клеточные популяции, обязанные развиваться в определенной области или части организма. Они определяются выражением конкретных комбинаций факторы транскрипции.
- Дифференциация клеток относится, в частности, к образованию функциональных типов клеток, таких как нервные, мышечные, секреторные эпителии и т.д. Дифференцированные клетки содержат большое количество специфических белков, связанных с функцией клетки.
- Морфогенез относится к формированию трехмерной формы. В основном он включает в себя организованные движения клеточных слоев и отдельных клеток. Морфогенез важен для создания трех зародышевых листков раннего эмбриона (эктодерма, мезодерма и энтодерма ) и для создания сложных структур в процессе развития органов.
- Рост тканей включает как общее увеличение размера ткани, так и дифференциальный рост частей (аллометрия ), который способствует морфогенезу. Рост в основном происходит за счет распространение клеток но также за счет изменения размера клеток или отложения внеклеточных материалов.
Процессы развития растений сходны с процессами развития животных. Однако клетки растений в большинстве своем неподвижны, поэтому морфогенез достигается за счет дифференциального роста без движений клеток. Кроме того, индуктивные сигналы и задействованные гены отличаются от тех, которые контролируют развитие животных.
Процессы развития
Дифференциация клеток
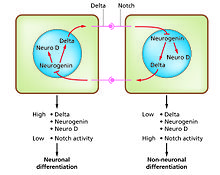
Дифференциация клеток это процесс, при котором в процессе развития возникают разные функциональные типы клеток. Например, нейроны, мышечные волокна и гепатоциты (клетки печени) являются хорошо известными типами дифференцированных клеток. Дифференцированные клетки обычно производят большое количество небольшого количества белков, необходимых для их конкретной функции, и это придает им характерный внешний вид, позволяющий распознавать их под световым микроскопом. Гены, кодирующие эти белки, очень активны. Обычно их хроматин структура очень открыта, что обеспечивает доступ для ферментов транскрипции, а определенные факторы транскрипции связываются с регуляторными последовательностями в ДНК, чтобы активировать экспрессию генов.[1][2] Например, NeuroD является ключевым фактором транскрипции для дифференцировки нейронов, миогенин для дифференциации мышц, и HNF4 дифференцировка гепатоцитов. Дифференцировка клеток обычно является заключительной стадией развития, которой предшествуют несколько состояний приверженности, которые не дифференцируются визуально. Одна ткань, образованная из одного типа клеток-предшественников или стволовых клеток, часто состоит из нескольких типов дифференцированных клеток. Контроль их образования включает в себя процесс бокового торможения,[3] на основе свойств Notch сигнальный путь.[4] Например, в нервной пластинке эмбриона эта система действует, чтобы генерировать популяцию нейрональных клеток-предшественников, в которых NeuroD экспрессируется в высокой степени.
Регенерация
Регенерация указывает на возможность отрастить недостающую часть.[5] Это очень распространено среди растений, которые демонстрируют непрерывный рост, а также среди колониальных животных, таких как гидроиды и асцидии. Но наибольший интерес биологи развития проявили к регенерации частей у свободноживущих животных. В частности, четыре модели были предметом большого исследования. Два из них обладают способностью регенерировать целые тела: Гидра, который может регенерировать любую часть полипа из небольшого фрагмента,[6] и планарий черви, которые обычно могут восстанавливать как головы, так и хвосты.[7] В обоих этих примерах непрерывный оборот клеток поддерживается стволовые клетки и, по крайней мере, у планарий, по крайней мере, некоторые стволовые клетки оказались плюрипотентный.[8] Две другие модели демонстрируют только дистальную регенерацию придатков. Это придатки насекомых, обычно ноги гемиметаболических насекомых, таких как сверчок,[9] и конечности уродельные амфибии.[10] В настоящее время имеется значительный объем информации о регенерации конечностей земноводных, и известно, что каждый тип клеток регенерирует сам себя, за исключением соединительных тканей, где происходит значительное взаимное преобразование хряща, дермы и сухожилий. Что касается структуры структур, то это контролируется реактивацией сигналов, активных в эмбрионе. До сих пор ведутся споры по поводу старого вопроса о том, является ли регенерация «изначальным» или «адаптивным» свойством.[11] Если первое верно, то с улучшенными знаниями мы можем ожидать, что сможем улучшить регенеративную способность у людей. В последнем случае предполагается, что каждый случай регенерации возник в результате естественного отбора в условиях, специфичных для данного вида, поэтому никаких общих правил не ожидается.
Эмбриональное развитие животных
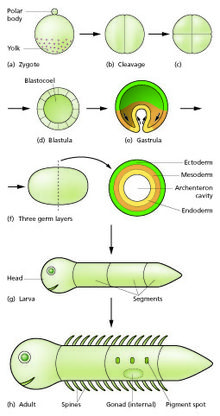

Сперма и яйцеклетка сливаются в процессе оплодотворения, образуя оплодотворенную яйцеклетку, или зигота.[12] Это претерпевает период делений, чтобы сформировать шар или лист подобных клеток, называемый бластула или же бластодерма. Эти деления клеток, как правило, происходят быстро без роста, поэтому дочерние клетки составляют половину размера материнской клетки, а весь эмбрион остается примерно того же размера. Они называются расщепление подразделения.
Мышь эпибласт изначальный стволовые клетки (см. рисунок: «Начальные стадии человеческого эмбриогенез ”) Пройти обширную эпигенетический перепрограммирование.[13] Этот процесс включает геном -широкий Деметилирование ДНК, хроматин реорганизация и эпигенетический стирание отпечатка, ведущее к тотипотентность.[13] Деметилирование ДНК осуществляется с помощью процесса, в котором используется ДНК. базовая эксцизионная пластика путь.[14]
Морфогенетические движения превращают клеточную массу в трехслойную структуру, состоящую из многоклеточных листов, называемых эктодерма, мезодерма и энтодерма. Эти листы известны как ростковые отростки. Это процесс гаструляция. Во время расщепления и гаструляции происходят первые события региональной спецификации. В дополнение к образованию самих трех зародышевых листков они часто генерируют экстраэмбриональные структуры, такие как у млекопитающих плацента, необходимые для поддержки и питания эмбриона,[15] а также установить различия в приверженности вдоль переднезадней оси (голова, туловище и хвост).[16]
Региональная спецификация инициируется наличием цитоплазматические детерминанты в одной части зиготы. Клетки, содержащие детерминант, становятся сигнальным центром и излучают индуцирующий фактор. Поскольку индуцирующий фактор производится в одном месте, диффундирует и распадается, он образует градиент концентрации, высокий около ячеек-источников и низкий подальше.[17][18] Остальные клетки эмбриона, которые не содержат детерминант, способны реагировать на различные концентрации посредством активации определенных генов контроля развития. Это приводит к созданию ряда зон, расположенных на все большем расстоянии от центра сигнализации. В каждой зоне активируется своя комбинация генов, контролирующих развитие.[19] Эти гены кодируют факторы транскрипции которые активируют новые комбинации активности генов в каждом регионе. Помимо других функций, эти факторы транскрипции контролируют экспрессию генов, придающих специфические адгезионные и подвижные свойства клеткам, в которых они активны. Из-за этих различных морфогенетических свойств клетки каждого зародышевого листка перемещаются, чтобы сформировать листы, так что эктодерма заканчивается снаружи, мезодерма в середине и энтодерма внутри.[20][21] Морфогенетические движения не только изменяют форму и структуру эмбриона, но, вводя клеточные листы в новые пространственные отношения, они также делают возможными новые фазы передачи сигналов и реакции между ними.
Рост у эмбрионов в основном автономен.[22] Для каждой территории клеток скорость роста контролируется комбинацией активных генов. Свободноживущие эмбрионы не растут в массе, поскольку у них нет внешнего источника питания. Но эмбрионы, питаемые плацентой или экстраэмбриональным желтком, могут расти очень быстро, и изменения относительной скорости роста между частями этих организмов помогают сформировать окончательную общую анатомию.
Весь процесс нужно координировать во времени, и как это контролируется, непонятно. Могут быть главные часы, способные связываться со всеми частями эмбриона, которые контролируют ход событий, или время может зависеть просто от локальной причинной последовательности событий.[23]
Метаморфоза
Процессы развития очень очевидны в процессе метаморфоза. Это происходит у разных видов животных. Хорошо известные примеры наблюдаются у лягушек, которые обычно вылупляются как головастики и превращаются во взрослую лягушку, а также у некоторых насекомых, которые вылупляются как личинки, а затем превращаются во взрослую форму на стадии куколки.
Все вышеперечисленные процессы развития происходят во время метаморфоза. Примеры, которые были особенно хорошо изучены, включают потерю хвоста и другие изменения в головастике лягушки. Xenopus,[24][25] и биология имагинальных дисков, которые производят взрослые части тела мухи Drosophila melanogaster.[26][27]
Развитие растений
Растение разработка это процесс, посредством которого структуры возникают и созревают по мере роста растения. Это изучается в анатомия растений и физиология растений а также морфология растений.
На протяжении всей жизни растения постоянно производят новые ткани и структуры из меристемы[28] расположены на концах органов или между зрелыми тканями. Таким образом, у живого растения всегда есть зародышевые ткани. Напротив, животное эмбрион очень рано произведет все части тела, которые когда-либо будут у него в жизни. Когда животное рождается (или вылупляется из яйца), у него есть все части тела, и с этого момента оно будет только расти и становиться более зрелым.
Свойства организации, наблюдаемые в растении: эмерджентные свойства которые больше, чем сумма отдельных частей. «Сборка этих тканей и функций в единый многоклеточный организм дает не только характеристики отдельных частей и процессов, но также совершенно новый набор характеристик, которые нельзя было бы предсказать на основе исследования отдельных частей».[29]
Рост
А сосудистое растение начинается с одноклеточной зигота, образована оплодотворение яйцеклетки с помощью сперматозоида. С этого момента он начинает делиться, образуя растение. эмбрион через процесс эмбриогенез. Когда это происходит, полученные клетки организуются так, что один конец становится первым корнем, а другой конец - верхушкой побега. В семя растения, у эмбриона разовьется один или несколько «семенных листков» (семядоли ). К концу эмбриогенеза у молодого растения будут все части, необходимые для начала жизни.
Однажды эмбрион прорастает из своего семени или родительского растения он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенез. Новые корни растут из корня меристемы расположен на кончике корня, и из побега вырастают новые стебли и листья меристемы находится на кончике побега.[30] Ветвление происходит, когда небольшие скопления клеток, оставленные меристемой и еще не подвергшиеся клеточная дифференциация чтобы сформировать специализированную ткань, начинают расти как верхушка нового корня или побега. Рост из любой такой меристемы на кончике корня или побега называется первичный рост и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега из делений клеток в камбий.[31]
Помимо роста на клетка деление, растение может прорасти удлинение ячейки.[32] Это происходит, когда отдельные клетки или группы клеток становятся длиннее. Не все растительные клетки вырастают до одинаковой длины. Когда клетки на одной стороне ствола растут дольше и быстрее, чем клетки на другой стороне, в результате ствол будет изгибаться в сторону более медленно растущих клеток. Этот направленный рост может происходить через реакцию растения на определенный стимул, такой как свет (фототропизм ), сила тяжести (гравитропизм ), воды, (гидротропизм ) и физический контакт (тигмотропизм ).
Рост и развитие растений опосредуются специфическими гормоны растений и регуляторы роста растений (ГРР) (Росс и др., 1983).[33] Уровни эндогенных гормонов зависят от возраста растений, морозостойкости, состояния покоя и других метаболических состояний; фотопериод, засуха, температура и другие внешние условия окружающей среды; и экзогенные источники ГРР, например, применяемые извне и ризосферного происхождения.
Морфологическая вариация
Растения демонстрируют естественные вариации по форме и строению. В то время как все организмы различаются от человека к человеку, растения демонстрируют дополнительный тип изменений. В пределах одного человека повторяются части, которые могут отличаться по форме и структуре от других подобных частей. Это изменение наиболее легко увидеть на листьях растения, хотя другие органы, такие как стебли и цветы, могут иметь аналогичные изменения. У этой вариации есть три основные причины: позиционные эффекты, влияние окружающей среды и молодость.
Эволюция морфологии растений
Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в морфогенезе растений и их эволюции. Во время посадки растений появилось много новых семейств факторов транскрипции, которые преимущественно связаны с сетями многоклеточного развития, размножения и развития органов, внося вклад в более сложный морфогенез наземных растений.[34]
У большинства наземных растений есть общий предок - многоклеточные водоросли. Пример эволюции морфологии растений наблюдается у харофитов. Исследования показали, что харофиты обладают чертами, гомологичными наземным растениям. Существуют две основные теории эволюции морфологии растений: теория гомологии и теория антитезиса. Общепринятая теория эволюции морфологии растений - это теория антитезиса. Антитетическая теория утверждает, что множественные митотические деления, которые происходят до мейоза, вызывают развитие спорофита. Тогда спорофит будет развиваться как самостоятельный организм.[35]
Организмы модели развития
Большая часть исследований в области биологии развития за последние десятилетия сосредоточена на использовании небольшого количества модельные организмы. Оказалось, что механизмы развития в животном мире в значительной степени сохранены. На раннем этапе развития разные виды позвоночных используют по существу одни и те же индуктивные сигналы и одни и те же гены, кодирующие региональную идентичность. Даже беспозвоночные используют аналогичный набор сигналов и генов, хотя сформированные части тела значительно отличаются. У каждого модельного организма есть определенные экспериментальные преимущества, благодаря которым они стали популярными среди исследователей. В одном смысле они являются «моделями» всего животного царства, а в другом - «моделями» человеческого развития, которое трудно изучать напрямую как по этическим, так и по практическим причинам. Модельные организмы были наиболее полезны для выяснения широкой природы механизмов развития. Чем больше требуется деталей, тем больше они отличаются друг от друга и от человека.
Растения:
- Тале кресс (Arabidopsis thaliana)
Позвоночные:
- Лягушка: Xenopus (X.laevis и tropicalis).[36][37] Хороший запас эмбрионов. Особенно подходит для микрохирургии.
- Данио: Данио Рерио.[38] Хороший запас эмбрионов. Хорошо развита генетика.
- Курица: Gallus gallus.[39] Ранние стадии похожи на млекопитающих, но с микрохирургией легче. Бюджетный.
- Мышь: Mus musculus.[40] Млекопитающее с хорошо развитой генетикой.
Беспозвоночные:
- Плодовая муха: Drosophila melanogaster.[41] Хороший запас эмбрионов. Хорошо развита генетика.
- Нематода: Caenorhabditis elegans.[42] Хороший запас эмбрионов. Хорошо развита генетика. Бюджетный.
Также популярны для некоторых целей морские ежи[43] и асцидии.[44] Для изучения регенерации уродельные амфибии такой как аксолотль Амбистома мексиканская используются,[45] а также плоские черви, такие как Schmidtea mediterranea.[7] Органоиды также были продемонстрированы как эффективная модель для развития.[46] Развитие растений сосредоточено на кресс-салате. Arabidopsis thaliana как модельный организм.[47]
Смотрите также
Рекомендации
- ^ Ли Б., Кэри М., Уоркман Дж. Л. (февраль 2007 г.). «Роль хроматина при транскрипции». Клетка. 128 (4): 707–19. Дои:10.1016 / j.cell.2007.01.015. PMID 17320508.
- ^ Хайнцман Н.Д., Стюарт Р.К., Хон Дж., Фу Й., Чинг С.В., Хокинс Р.Д. и др. (Март 2007 г.). «Отчетливые и предсказуемые сигнатуры хроматина промоторов и энхансеров транскрипции в геноме человека». Природа Генетика. 39 (3): 311–8. Дои:10,1038 / ng1966. PMID 17277777.
- ^ Мейнхардт Х, Гирер А (2000). «Формирование паттерна путем локальной самоактивации и бокового торможения» (PDF). BioEssays. 22 (8): 753–760. CiteSeerX 10.1.1.477.439. Дои:10.1002 / 1521-1878 (200008) 22: 8 <753 :: help-bies9> 3.0.co; 2-z. PMID 10918306. В архиве (PDF) из оригинала от 27.10.2017.
- ^ Спринзак Д., Лакханпал А., Лебон Л., Сантат Л.А., Фонтес М.Э., Андерсон Г.А. и др. (Май 2010 г.). «Цис-взаимодействия между Notch и Delta генерируют взаимоисключающие состояния сигнализации». Природа. 465 (7294): 86–90. Bibcode:2010Натура.465 ... 86S. Дои:10.1038 / природа08959. ЧВК 2886601. PMID 20418862.
- ^ Карлсон Б.М. (2007). Принципы регенеративной биологии. Берлингтон, Массачусетс: Academic Press.
- ^ Bosch TC (март 2007 г.). «Почему полипы регенерируют, а мы нет: к клеточному и молекулярному каркасу для регенерации гидры». Биология развития. 303 (2): 421–33. Дои:10.1016 / j.ydbio.2006.12.012. PMID 17234176.
- ^ а б Reddien PW, Санчес Альварадо А (2004). «Основы регенерации планарий». Ежегодный обзор клеточной биологии и биологии развития. 20: 725–57. Дои:10.1146 / annurev.cellbio.20.010403.095114. PMID 15473858. S2CID 1320382.
- ^ Вагнер Д.Е., Ван И.Е., Reddien PW (май 2011 г.). «Клоногенные необласты - это плюрипотентные взрослые стволовые клетки, лежащие в основе регенерации планарии». Наука. 332 (6031): 811–6. Bibcode:2011Sci ... 332..811W. Дои:10.1126 / science.1203983. ЧВК 3338249. PMID 21566185.
- ^ Накамура Т., Мито Т., Бандо Т., Охучи Х., Нодзи С. (январь 2008 г.). «Рассекающая регенерация ног насекомых посредством РНК-интерференции». Клеточные и молекулярные науки о жизни. 65 (1): 64–72. Дои:10.1007 / s00018-007-7432-0. PMID 18030418.
- ^ Саймон А., Танака Э.М. (2013). «Регенерация конечностей». Междисциплинарные обзоры Wiley. Биология развития. 2 (2): 291–300. Дои:10.1002 / wdev.73. PMID 24009038.
- ^ Slack JM (2013). «Глава 20». Основная биология развития. Оксфорд: Wiley-Blackwell.
- ^ Юнгникель М.К., Саттон К.А., Флорман Х.М. (август 2003 г.). «В начале: уроки оплодотворения мышей и червей». Клетка. 114 (4): 401–4. Дои:10.1016 / s0092-8674 (03) 00648-2. PMID 12941269.
- ^ а б Хакетт Дж. А., Сенгупта Р., Зилич Дж. Дж., Мураками К., Ли С., Даун Т. А., Сурани М. А. (январь 2013 г.). «Динамика деметилирования ДНК зародышевой линии и стирание отпечатка с помощью 5-гидроксиметилцитозина». Наука. 339 (6118): 448–52. Bibcode:2013Наука ... 339..448H. Дои:10.1126 / science.1229277. ЧВК 3847602. PMID 23223451.
- ^ Хайкова П., Джеффрис С.Дж., Ли С., Миллер Н., Джексон С.П., Сурани М.А. (июль 2010 г.). «Полногеномное перепрограммирование в зародышевой линии мыши влечет за собой основной путь эксцизионной репарации». Наука. 329 (5987): 78–82. Bibcode:2010Sci ... 329 ... 78H. Дои:10.1126 / science.1187945. ЧВК 3863715. PMID 20595612.
- ^ Стивен Д.Х., изд. (1975). Сравнительная плацентация. Лондон: Academic Press.
- ^ Кимельман Д., Мартин Б.Л. (2012). «Передне-задний паттерн в раннем развитии: три стратегии». Междисциплинарные обзоры Wiley. Биология развития. 1 (2): 253–66. Дои:10.1002 / wdev.25. ЧВК 5560123. PMID 23801439.
- ^ Slack JM (1987). «Морфогенетические градиенты - прошлое и настоящее». Тенденции в биохимических науках. 12: 200–204. Дои:10.1016/0968-0004(87)90094-6.
- ^ Роджерс К.В., Шир А.Ф. (2011). «Градиенты морфогенов: от поколения к интерпретации». Ежегодный обзор клеточной биологии и биологии развития. 27: 377–407. Дои:10.1146 / annurev-cellbio-092910-154148. PMID 21801015. S2CID 21477124.
- ^ Dahmann C, Oates AC, Brand M (январь 2011 г.). «Формирование и поддержание границ в развитии тканей». Обзоры природы. Генетика. 12 (1): 43–55. Дои:10.1038 / nrg2902. PMID 21164524.
- ^ Хардин Дж., Уолстон Т. (август 2004 г.). «Модели морфогенеза: механизмы и механика перестройки клеток». Текущее мнение в области генетики и развития. 14 (4): 399–406. Дои:10.1016 / j.gde.2004.06.008. PMID 15261656.
- ^ Hammerschmidt M, Wedlich D (ноябрь 2008 г.). «Регулируемая адгезия как движущая сила гаструляционных движений». Разработка. 135 (22): 3625–41. Дои:10.1242 / dev.015701. PMID 18952908.
- ^ О'Фаррелл PH (2003). «Как многоклеточные животные достигают своего полного размера: естественная история размера». В зале MN, Raff M, Thomas G (ред.). Рост клеток: контроль размера клеток. Лабораторный пресс Колд-Спринг-Харбор. С. 1–21.
- ^ Мосс Э.Г., Ромер-Зайберт Дж. (2014). «Внутреннее время клетки в развитии животных». Междисциплинарные обзоры Wiley. Биология развития. 3 (5): 365–77. Дои:10.1002 / wdev.145. PMID 25124757.
- ^ Тата-младший (1996). «Метаморфозы амфибий: изысканная модель гормональной регуляции постэмбрионального развития позвоночных». Развитие, рост и дифференциация. 38 (3): 223–231. Дои:10.1046 / j.1440-169x.1996.t01-2-00001.x.
- ^ Браун Д.Д., Цай Л. (июнь 2007 г.). «Метаморфоза амфибии». Биология развития. 306 (1): 20–33. Дои:10.1016 / j.ydbio.2007.03.021. ЧВК 1945045. PMID 17449026.
- ^ Коэн С.М. (1993). «Развитие имагинального диска». В Bate M, Martinez-Arias M (ред.). Развитие Drosophila melanogaster. Колд Спринг Харбор Пресс.
- ^ Maves L, Schubiger G (октябрь 2003 г.). «Трансдетерминация в имагинальных дисках Drosophila: модель для понимания плюрипотентности и поддержания селекторного гена». Текущее мнение в области генетики и развития. 13 (5): 472–9. Дои:10.1016 / j.gde.2003.08.006. PMID 14550411.
- ^ Bäurle I, Laux T (октябрь 2003 г.). «Апикальные меристемы: фонтан молодости растений». Рассмотрение. BioEssays. 25 (10): 961–70. Дои:10.1002 / bies.10341. PMID 14505363.
- ^ Леопольд AC (1964). Рост и развитие растений. Нью-Йорк: Макгроу-Хилл. п.183.
- ^ Бренд U, Hobe M, Simon R (февраль 2001 г.). «Функциональные домены в меристемах побегов растений». Рассмотрение. BioEssays. 23 (2): 134–41. Дои:10.1002 / 1521-1878 (200102) 23: 2 <134 :: AID-BIES1020> 3.0.CO; 2-3. PMID 11169586.
- ^ Барлоу П. (май 2005 г.). «Узорчатое определение клеток в растительной ткани: вторичная флоэма деревьев». BioEssays. 27 (5): 533–41. Дои:10.1002 / bies.20214. PMID 15832381.
- ^ Пасифици Э., Ди Мамбро Р., Делло Иоио Р., Костантино П., Сабатини С. (август 2018 г.). «Корень арабидопсиса». Журнал EMBO. 37 (16). Дои:10.15252 / embj.201899134. ЧВК 6092616. PMID 30012836.
- ^ Росс С.Д., Фарис Р.П., Биндер В.Д. (1983). «Регуляторы роста и хвойные деревья: их физиология и потенциальное использование в лесном хозяйстве». В Никелле LG (ред.). Химические вещества, регулирующие рост растений. 2. Бока-Ратон, Флорида: CRC Press. С. 35–78.
- ^ Jin J, He K, Tang X, Li Z, Lv L, Zhao Y и др. (Июль 2015 г.). «Карта регуляторов транскрипции Arabidopsis выявляет отличительные функциональные и эволюционные особенности новых факторов транскрипции». Молекулярная биология и эволюция. 32 (7): 1767–73. Дои:10.1093 / molbev / msv058. ЧВК 4476157. PMID 25750178. В архиве из оригинала от 02.06.2016.
- ^ Pires, Nuno D .; Долан, Лиам (19 февраля 2012 г.). «Морфологическая эволюция наземных растений: новые конструкции со старыми генами». Философские труды Королевского общества B: биологические науки. 367 (1588): 508–518. Дои:10.1098 / rstb.2011.0252. ISSN 0962-8436. ЧВК 3248709. PMID 22232763.
- ^ Nieuwkoop PD, Faber J (1967). Нормальный стол Xenopus laevis (Даудин). Северная Голландия, Амстердам.
- ^ Харланд Р.М., Грейнджер Р.М. (декабрь 2011 г.). «Исследование Xenopus: метаморфозы генетики и геномики». Тенденции в генетике. 27 (12): 507–15. Дои:10.1016 / j.tig.2011.08.003. ЧВК 3601910. PMID 21963197.
- ^ Лоусон Н.Д., Вулф С.А. (июль 2011 г.). «Прямые и обратные генетические подходы для анализа развития позвоночных у рыбок данио». Клетка развития. 21 (1): 48–64. Дои:10.1016 / j.devcel.2011.06.007. PMID 21763608.
- ^ Рашиди Х., Соттиль V (апрель 2009 г.). «Куриный эмбрион: модель для современных биомедицинских исследований». BioEssays. 31 (4): 459–65. Дои:10.1002 / bies.200800168. PMID 19274658.
- ^ Берингер Р., Герценштейн М., Винтерстен К., Надь М. (2014). Манипулирование эмбрионом мыши. Лабораторное руководство (Четвертое изд.). Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор.
- ^ Сент-Джонстон Д. (март 2002 г.). «Искусство и дизайн генетических экранов: Drosophila melanogaster». Обзоры природы. Генетика. 3 (3): 176–88. Дои:10.1038 / nrg751. PMID 11972155.
- ^ Загадка Д.Л., Блюменталь Т., Мейер Б.Дж., Присс-младший (1997). C.elegans II. Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор.
- ^ Эттенсон CA, Sweet HC (2000). Создание рисунка раннего эмбриона морского ежа. Curr. Вершина. Dev. Биол. Актуальные темы биологии развития. 50. Академическая пресса. стр.1–44. Дои:10.1016 / S0070-2153 (00) 50002-7. ISBN 9780121531508. PMID 10948448.
- ^ Lemaire P (июнь 2011 г.). «Эволюционный перекресток в биологии развития: оболочки». Разработка. 138 (11): 2143–52. Дои:10.1242 / dev.048975. PMID 21558365.
- ^ Наку Э., Танака Э.М. (2011). «Регенерация конечностей: новое развитие?». Ежегодный обзор клеточной биологии и биологии развития. 27: 409–40. Дои:10.1146 / annurev-cellbio-092910-154115. PMID 21801016.
- ^ Адер М, Танака Э.М. (декабрь 2014 г.). «Моделирование человеческого развития в 3D-культуре». Текущее мнение в области клеточной биологии. 31: 23–8. Дои:10.1016 / j.ceb.2014.06.013. PMID 25033469.
- ^ Вайгель Д., Глейзбрук Дж. (2002). Арабидопсис. Лабораторное руководство. Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор.
дальнейшее чтение
- Гилберт С.Ф. (2013). Биология развития. Сандерленд, Массачусетс: Sinauer Associates Inc.
- Slack JM (2013). Основная биология развития. Оксфорд: Wiley-Blackwell.
- Вольперт Л., Щекотка С. (2011). Принципы развития. Оксфорд и Нью-Йорк: Издательство Оксфордского университета.