Нервная система - Nervous system
| Нервная система | |
|---|---|
 Нервная система человека | |
| Подробности | |
| Идентификаторы | |
| латинский | systema nervosum |
| MeSH | D009420 |
| FMA | 7157 |
| Анатомическая терминология | |
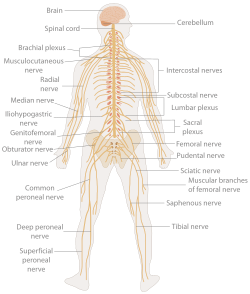
В биология, то нервная система это очень сложный часть животное который координирует его действия и сенсорный информацию путем передачи сигналы в разные части его тела и обратно. Нервная система обнаруживает изменения окружающей среды, которые влияют на организм, а затем работает в тандеме с эндокринная система реагировать на такие события.[1] Нервная ткань впервые возник в червеобразные организмы около 550-600 миллионов лет назад. У позвоночных он состоит из двух основных частей: Центральная нервная система (ЦНС) и периферическая нервная система (ПНС). ЦНС состоит из мозг и спинной мозг. PNS состоит в основном из нервы, которые представляют собой замкнутые пучки длинных волокон или аксоны, которые соединяют ЦНС со всеми остальными частями тела. Нервы, передающие сигналы от мозга, называются мотор или же эфферент нервы, а те нервы, которые передают информацию от тела к ЦНС, называются сенсорный или же афферентный. Позвоночные нервы выполняют обе функции и называются смешанный нервы. PNS разделена на три отдельные подсистемы: соматический, автономный, и кишечный нервная система. Соматические нервы опосредуют произвольные движения. Вегетативная нервная система далее подразделяется на сочувствующий и парасимпатический нервная система. Симпатическая нервная система активируется в чрезвычайных ситуациях для мобилизации энергии, в то время как парасимпатическая нервная система активируется, когда организмы находятся в расслабленном состоянии. Кишечная нервная система контролирует желудочно-кишечный система. И вегетативная, и кишечная нервные системы функционируют непроизвольно. Нервы, выходящие из черепа, называются черепные нервы в то время как выходящие из спинного мозга называются позвоночные нервы.
На клеточном уровне нервная система определяется наличием особого типа клеток, называемых клетками. нейрон, также известная как «нервная клетка». У нейронов есть особые структуры, которые позволяют им быстро и точно посылать сигналы другим клеткам. Они посылают эти сигналы в виде электрохимических волн, распространяющихся по тонким волокнам, называемым аксоны, которые вызывают химические вещества, называемые нейротрансмиттеры быть выпущенным на стыках, называемых синапсы. Клетка, которая получает синаптический сигнал от нейрона, может быть возбуждена, подавлена или иным образом модулирована. Связи между нейронами могут образовывать нервные пути, нейронные цепи, и больше сети которые формируют восприятие мира организмом и определяют его поведение. Наряду с нейронами нервная система содержит другие специализированные клетки, называемые глиальные клетки (или просто глия), которые обеспечивают структурную и метаболическую поддержку.
Нервные системы встречаются у большинства многоклеточных животных, но сильно различаются по сложности.[2] Единственные многоклеточные животные, у которых вообще нет нервной системы, - это губки, плакозои, и мезозои, у которых очень простые планы кузова. Нервная система радиально симметричный организмы гребневики (гребешки) и книдарийцы (который включает в себя анемоны, гидры, кораллы и медуза ) состоят из диффузного нервная сеть. У всех других видов животных, за исключением нескольких типов червей, есть нервная система, содержащая мозг, центральный шнур (или два соединяющихся шнура). параллельно ), и нервы, исходящие от головного мозга и центрального канатика. Размер нервной системы колеблется от нескольких сотен клеток у простейших червей до примерно 300 миллиардов клеток у африканских слонов.[3]
Центральная нервная система отправляет сигналы от одной клетки к другой или от одной части тела к другим и получает обратную связь. Неисправность нервной системы может возникнуть в результате генетических дефектов, физического повреждения в результате травмы или токсичности, инфекции или просто ощущение. Медицинская специальность неврология изучает нарушения нервной системы и ищет способы их предотвращения или лечения. В периферической нервной системе наиболее распространенной проблемой является нарушение нервной проводимости, которое может быть вызвано разными причинами, в том числе: диабетическая невропатия и демиелинизирующие расстройства, такие как рассеянный склероз и боковой амиотрофический склероз. Неврология это область науки, которая фокусируется на изучении нервной системы.
Структура
Нервная система получила свое название от нервов, которые представляют собой цилиндрические пучки волокон ( аксоны из нейроны ), которые исходят из мозга и спинной мозг, и несколько раз ветвятся, чтобы иннервировать каждую часть тела.[4] Нервы достаточно большие, чтобы их распознали древние египтяне, греки и римляне.[5] но их внутреннее строение не было понято, пока не стало возможным исследовать их с помощью микроскопа.[6] Автор Майкл Николецис писал:[7]
«Трудно поверить, что примерно до 1900 года не было известно, что нейроны являются основными единицами мозга (Сантьяго Рамон-и-Кахаль ). Не менее удивительным является тот факт, что концепция передачи химических веществ в мозге не была известна примерно до 1930 г. (Генри Халлетт Дейл и Отто Леви ). Мы начали понимать основное электрическое явление, которое нейроны используют для связи между собой, потенциал действия, в 1950-х годах (Алан Ллойд Ходжкин, Эндрю Хаксли и Джон Экклс ). Это было в 1960-х годах, когда мы узнали, как основные нейронные сети кодируют стимулы и, следовательно, возможны основные концепции (Дэвид Хьюбел и Торстен Визель ). Молекулярная революция охватила университеты США в 1980-х годах. Именно в 1990-е годы стали широко известны молекулярные механизмы поведенческих явлений (Эрик Ричард Кандел )."
Исследование под микроскопом показывает, что нервы состоят в основном из аксонов, а также из различных мембран, которые обвивают их и разделяют на пучки. Нейроны, дающие начало нервам, не лежат полностью внутри самих нервов - их клеточные тела находятся в головном мозге, спинной мозг, или периферийный ганглии.[4]
Все животные, более развитые, чем губки, имеют нервную систему. Однако даже губки, одноклеточные животные и неживотные, такие как слизистые плесени, имеют межклеточные сигнальные механизмы, которые являются предшественниками нейронов.[8] У радиально-симметричных животных, таких как медузы и гидры, нервная система состоит из нервная сеть, диффузная сеть изолированных клеток.[9] В двухсторонний у животных, которые составляют подавляющее большинство существующих видов, нервная система имеет общую структуру, которая возникла в Эдиакарский период, более 550 миллионов лет назад.[10][11]
Клетки
Нервная система состоит из двух основных категорий или типов клеток: нейроны и глиальные клетки.
Нейроны
| Нейрон |
|---|

Нервная система определяется наличием особого типа клеток - нейрон (иногда называемый «нейрон» или «нервная клетка»).[4] Нейроны можно отличить от других клеток разными способами, но их наиболее фундаментальное свойство состоит в том, что они общаются с другими клетками через синапсы, которые представляют собой соединения мембраны с мембраной, содержащие молекулярные механизмы, обеспечивающие быструю передачу сигналов, электрических или химических.[4] Многие типы нейронов обладают аксон протоплазматический выступ, который может распространяться на отдаленные части тела и создавать тысячи синаптических контактов;[12] аксоны обычно проходят по всему телу в пучках, называемых нервами.
Даже в нервной системе одного вида, такого как человек, существуют сотни различных типов нейронов с широким разнообразием морфологии и функций.[12] К ним относятся сенсорные нейроны которые преобразуют физические стимулы, такие как свет и звук, в нейронные сигналы, и двигательные нейроны которые преобразуют нервные сигналы в активацию мышц или желез; однако у многих видов подавляющее большинство нейронов участвуют в формировании централизованных структур (головного мозга и ганглиев), и они получают все свои входные данные от других нейронов и отправляют свои выходные данные другим нейронам.[4]
Глиальные клетки
Глиальные клетки (названные от греческого слова «клей») - это ненейронные клетки, которые обеспечивают поддержку и питание, поддерживать гомеостаз, форма миелин, и участвуют в передаче сигналов в нервной системе.[13] в человеческий мозг, считается, что общее количество глии примерно равно количеству нейронов, хотя пропорции различаются в разных областях мозга.[14] Среди наиболее важных функций глиальных клеток - поддержка нейронов и удержание их на месте; снабжать нейроны питательными веществами; электрически изолировать нейроны; разрушать патогены и удалить мертвые нейроны; и предоставить подсказки, направляющие аксоны нейронов к их мишеням.[13] Очень важный тип глиальной клетки (олигодендроциты в центральной нервной системе, и Клетки Шванна в периферической нервной системе) образует слои жирового вещества, называемого миелин который оборачивается вокруг аксонов и обеспечивает электрическую изоляцию, которая позволяет им передавать потенциалы действия намного быстрее и эффективнее. Недавние открытия показывают, что глиальные клетки, такие как микроглия и астроциты, служат важными резидентными иммунными клетками в центральной нервной системе.
Анатомия позвоночных

Нервная система позвоночные (включая людей) делится на Центральная нервная система (ЦНС) и периферическая нервная система (ПНС).[15]
(ЦНС) является основным отделом и состоит из мозг и спинной мозг.[15] В спинномозговой канал содержит спинной мозг, а полость черепа содержит мозг. ЦНС закрыта и защищена мозговые оболочки, трехслойная система мембран, включающая жесткий кожаный внешний слой, называемый твёрдая мозговая оболочка. Головной мозг также защищен черепом, а спинной мозг - позвонки.
Периферическая нервная система (ПНС) - это собирательный термин для структур нервной системы, которые не находятся в ЦНС.[16] Считается, что подавляющее большинство пучков аксонов, называемых нервами, принадлежит ПНС, даже если клеточные тела нейронов, которым они принадлежат, находятся в головном или спинном мозге. ПНС делится на соматический и висцеральный части. Соматическая часть состоит из нервов, которые иннервируют кожу, суставы и мышцы. Тела соматических сенсорных нейронов лежат в ганглии задних корешков спинного мозга. Висцеральная часть, также известная как вегетативная нервная система, содержит нейроны, которые иннервируют внутренние органы, кровеносные сосуды и железы. Сама вегетативная нервная система состоит из двух частей: Симпатическая нервная система и парасимпатическая нервная система. Некоторые авторы также включают сенсорные нейроны, чьи клеточные тела лежат на периферии (для таких органов чувств, как слух), как часть ПНС; другие же их опускают.[17]
Нервную систему позвоночных также можно разделить на области, называемые серое вещество и белое вещество.[18] Серое вещество (которое является только серым в консервированной ткани и лучше описывается как розовое или светло-коричневое в живой ткани) содержит высокую долю клеточных тел нейронов. Белое вещество состоит в основном из миелинизированный аксонов, и берет свой цвет от миелина. Белое вещество включает в себя все нервы и большую часть внутренних частей головного и спинного мозга. Серое вещество находится в скоплениях нейронов головного и спинного мозга, а также в корковых слоях, выстилающих их поверхности. Существует анатомическое соглашение о том, что кластер нейронов в головном или спинном мозге называется ядро, тогда как скопление нейронов на периферии называется ганглий.[19] Однако есть несколько исключений из этого правила, в частности, в том числе часть переднего мозга, называемая базальный ганглий.[20]
Сравнительная анатомия и эволюция
Нейронные предшественники в губках
Губки не имеют ячеек, связанных друг с другом синаптические соединения, то есть без нейронов и, следовательно, без нервной системы. Однако у них есть гомологи многих генов, которые играют ключевую роль в синаптической функции. Недавние исследования показали, что клетки губки экспрессируют группу белков, которые группируются вместе, образуя структуру, напоминающую постсинаптическая плотность (приемная часть синапса).[8] Однако функция этой структуры в настоящее время неясна. Хотя клетки губки не демонстрируют синаптической передачи, они взаимодействуют друг с другом посредством волн кальция и других импульсов, которые опосредуют некоторые простые действия, такие как сокращение всего тела.[21]
Radiata
Медуза, гребешки, и родственные животные имеют диффузные нервные сети, а не центральную нервную систему. У большинства медуз нервная сеть более или менее равномерно распределена по телу; в гребешках он сконцентрирован около рта. Нервные сети состоят из сенсорных нейронов, которые улавливают химические, тактильные и визуальные сигналы; мотонейроны, которые могут активировать сокращения стенки тела; и промежуточные нейроны, которые обнаруживают паттерны активности сенсорных нейронов и в ответ посылают сигналы группам моторных нейронов. В некоторых случаях группы промежуточных нейронов группируются в дискретные ганглии.[9]
Развитие нервной системы у радиата относительно неструктурирован. В отличие от билатерии, radiata имеют только два первичных клеточных слоя, энтодерма и эктодерма. Нейроны генерируются из особого набора эктодермальных клеток-предшественников, которые также служат предшественниками для любого другого типа эктодермальных клеток.[22]
Bilateria

Подавляющее большинство существующих животных билатерии, что означает животных, у которых левая и правая стороны являются приблизительно зеркальным отображением друг друга. Считается, что все bilateria произошли от общего червеобразного предка, который появился в эдиакарский период, 550–600 миллионов лет назад.[10] Основная форма билатерального тела представляет собой трубку с полой кишкой, проходящей ото рта к анусу, и нервный шнур с расширением («ганглием») для каждого сегмента тела с особенно большим ганглием спереди, называемым «мозгом». ".

Даже млекопитающие, в том числе люди, демонстрируют сегментированный билатерианский план тела на уровне нервной системы. Спинной мозг содержит серию сегментарных ганглиев, каждый из которых дает начало двигательным и чувствительным нервам, которые иннервируют часть поверхности тела и подлежащую мускулатуру. На конечностях схема иннервации сложна, но на туловище она дает серию узких полос. Три верхних сегмента принадлежат головному мозгу, давая начало переднему, среднему и заднему мозгу.[23]
На основании событий, происходящих на очень ранних стадиях эмбрионального развития, Bilaterians можно разделить на две группы (суперфила ) называется протостомы и дейтеростомы.[24] Deuterostomes включают позвоночных, а также иглокожие, полухордовые (в основном желудевые черви) и Ксенотурбеллиданы.[25] Протостомы, более разнообразная группа, включают: членистоногие, моллюски, и многочисленные виды червей. Между этими двумя группами существует фундаментальное различие в расположении нервной системы внутри тела: протостомы имеют нервный шнур на вентральной (обычно нижней) стороне тела, тогда как у дейтеростомов нервный шнур находится на дорсальной (обычно верхней) стороне тела. ) сторона. Фактически, многие аспекты тела инвертированы между двумя группами, включая паттерны экспрессии нескольких генов, которые демонстрируют градиенты от дорсального к вентральному. Большинство анатомов в настоящее время считают, что тела протостомов и дейтеростомов "перевернуты" относительно друг друга, и эта гипотеза была впервые предложена Жоффруа Сен-Илер для насекомых по сравнению с позвоночными. Так, например, у насекомых есть нервные связки, которые проходят вдоль средней линии вентральной части тела, в то время как у всех позвоночных есть спинной мозг, проходящий вдоль средней линии спины.[26]
черви
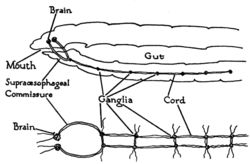
черви являются простейшими двустворчатыми животными и наиболее наглядно раскрывают основную структуру двуногой нервной системы. В качестве примера, дождевые черви иметь двойной нервные шнуры проходит по длине тела и сливается у хвоста и рта. Эти нервные тяжи связаны между собой поперечный нервы, как ступеньки лестницы. Эти поперечные нервы помогают координировать две стороны животного. Два ганглии во главе ("нервное кольцо ") конечная функция, аналогичная простой мозг. Фоторецепторы на глазных пятнах животного предоставляют сенсорную информацию о свете и темноте.[27]
Нервная система одного очень маленького круглого червя, нематода Caenorhabditis elegans, был полностью отображен в коннектом включая его синапсы. Каждый нейрон и его клеточная линия был зарегистрирован, и большинство, если не все, нейронные связи известны. У этого вида нервная система сексуально диморфный; нервные системы обоих полов, мужчин и женщин гермафродиты, имеют разное количество нейронов и групп нейронов, которые выполняют специфичные для пола функции. В C. elegans, у мужчин ровно 383 нейрона, а у гермафродитов - ровно 302 нейрона.[28]
Членистоногие

Членистоногие, Такие как насекомые и ракообразные имеют нервную систему, состоящую из ряда ганглии, соединенный брюшной нервный тяж состоит из двух параллельных связок, идущих по длине живот.[29] Обычно каждый сегмент тела имеет один ганглий с каждой стороны, хотя некоторые ганглии сливаются, образуя мозг и другие крупные ганглии. Головной сегмент содержит мозг, также известный как надэзофагеальный ганглий. в нервная система насекомых, мозг анатомически делится на протоцеребрум, дейтоцеребрум, и тритоцеребрум. Сразу за мозгом находится подэзофагеальный ганглий, который состоит из трех пар сросшихся ганглиев. Он контролирует ротовые органы, то слюнные железы и некоторые мышцы. Многие членистоногие имеют хорошо развитые сенсорный органы, в том числе сложные глаза для видения и усики за обоняние и феромон ощущение. Сенсорная информация от этих органов обрабатывается мозгом.
У насекомых многие нейроны имеют клеточные тела, которые расположены на краю мозга и являются электрически пассивными - клеточные тела служат только для обеспечения метаболической поддержки и не участвуют в передаче сигналов. Протоплазматическое волокно проходит от тела клетки и обильно разветвляется, при этом одни части передают сигналы, а другие принимают сигналы. Таким образом, большая часть мозг насекомого тела пассивных клеток расположены по периферии, в то время как обработка нервных сигналов происходит в клубке протоплазматических волокон, называемых нейропиль, в интерьере.[30]
«Идентифицированные» нейроны
Нейрон называется идентифицированный если у него есть свойства, которые отличают его от любого другого нейрона того же животного - такие свойства, как местоположение, нейротрансмиттер, паттерн экспрессии генов и связь - и если каждый отдельный организм, принадлежащий к тому же виду, имеет один и только один нейрон с таким же набором свойств.[31] В нервных системах позвоночных очень мало нейронов «идентифицируются» в этом смысле - у людей, как полагают, их нет, - но в более простых нервных системах некоторые или все нейроны могут быть, таким образом, уникальными. В аскариде C. elegans, нервная система которого наиболее подробно описана среди всех животных, каждый нейрон в организме уникально идентифицируется, имеет одинаковое местоположение и одинаковые связи в каждом отдельном черве. Одним из заметных следствий этого факта является то, что форма C. elegans нервная система полностью определена геномом, без зависимости от опыта.[28]
Мозг многих моллюсков и насекомых также содержит значительное количество идентифицированных нейронов.[31] У позвоночных наиболее известными идентифицированными нейронами являются гигантские Клетки Маутнера рыбы.[32] У каждой рыбы есть две клетки Маутнера в нижней части ствола мозга, одна слева, а другая - справа. Каждая клетка Маутнера имеет аксон, который пересекает, иннервируя нейроны на том же уровне мозга, а затем движется вниз по спинному мозгу, создавая многочисленные связи на своем пути. Синапсы, генерируемые маутнеровской клеткой, настолько мощны, что единственный потенциал действия вызывает серьезную поведенческую реакцию: в течение миллисекунд рыба изгибает свое тело в форму. C-образная форма, затем выпрямляется, тем самым быстро продвигаясь вперед. Функционально это быстрая реакция на побег, которая наиболее легко запускается сильной звуковой волной или волной давления, падающей на орган боковой линии рыбы. Клетки Маутнера - не единственные идентифицированные нейроны у рыб - существует еще около 20 типов, включая пары «аналогов клеток Маутнера» в каждом сегментарном ядре спинного мозга. Хотя клетка Маутнера способна вызывать ответную реакцию по отдельности, в контексте обычного поведения клетки других типов обычно вносят вклад в формирование амплитуды и направления ответа.
Клетки Маутнера были описаны как командные нейроны. Командный нейрон - это особый тип идентифицированного нейрона, определяемый как нейрон, который способен индивидуально управлять определенным поведением.[33] Такие нейроны чаще всего появляются в системах быстрого бегства различных видов - гигантский аксон кальмара и кальмар гигантский синапс, используемые для новаторских экспериментов в нейрофизиологии из-за их огромных размеров, оба участвуют в схеме быстрого побега кальмара. Однако концепция командного нейрона стала противоречивой из-за исследований, показывающих, что некоторые нейроны, которые первоначально казались соответствующими описанию, действительно были способны вызывать реакцию только в ограниченном наборе обстоятельств.[34]
Функция
На самом базовом уровне функция нервной системы заключается в посылке сигналов от одной клетки к другим или от одной части тела к другим. Есть несколько способов, которыми ячейка может посылать сигналы другим ячейкам. Один из них - выпуск химикатов, гормоны во внутреннее кровообращение, чтобы они могли распространяться в отдаленные места. В отличие от этого «широковещательного» режима передачи сигналов, нервная система обеспечивает сигналы «точка-точка» - нейроны проецируют свои аксоны в определенные целевые области и создают синаптические связи с определенными целевыми клетками.[35] Таким образом, нейронная передача сигналов имеет гораздо более высокий уровень специфичности, чем передача гормональных сигналов. Это также намного быстрее: самые быстрые нервные сигналы передаются со скоростью, превышающей 100 метров в секунду.
На более интегративном уровне основная функция нервной системы - контролировать тело.[4] Он делает это путем извлечения информации из окружающей среды с помощью сенсорных рецепторов, отправки сигналов, кодирующих эту информацию, в центральную нервную систему, обработки информации для определения соответствующей реакции и отправки выходных сигналов мышцам или железам для активации реакции. Эволюция сложной нервной системы позволила различным видам животных обрести развитые способности восприятия, такие как зрение, сложные социальные взаимодействия, быстрая координация систем органов и интегрированная обработка параллельных сигналов. У людей развитая нервная система делает возможным язык, абстрактное представление концепций, передачу культуры и многие другие особенности человеческого общества, которые не существовали бы без человеческого мозга.
Нейроны и синапсы

Большинство нейронов посылают сигналы через свои аксоны, хотя некоторые типы способны к обмену данными между дендритами. (Фактически, типы нейронов, называемые амакриновые клетки не имеют аксонов и общаются только через свои дендриты.) Нейронные сигналы распространяются вдоль аксона в виде электрохимических волн, называемых потенциалы действия, которые производят сигналы от ячейки к ячейке в точках, где терминалы аксонов делать синаптический контакт с другими клетками.[36]
Синапсы могут быть электрическими или химическими. Электрические синапсы устанавливать прямые электрические связи между нейронами,[37] но химические синапсы гораздо более распространены и гораздо более разнообразны по функциям.[38] В химическом синапсе клетка, которая посылает сигналы, называется пресинаптической, а клетка, которая принимает сигналы, называется постсинаптической. И пресинаптическая, и постсинаптическая области заполнены молекулярными механизмами, которые осуществляют процесс передачи сигналов. Пресинаптическая область содержит большое количество крошечных сферических сосудов, называемых синаптические везикулы, упакованный нейротрансмиттер химикаты.[36] Когда пресинаптический терминал электрически стимулируется, активируется множество молекул, встроенных в мембрану, и заставляет содержимое везикул высвобождаться в узкое пространство между пресинаптической и постсинаптической мембранами, называемое синаптическая щель. Затем нейротрансмиттер связывается с рецепторы встроены в постсинаптическую мембрану, заставляя их перейти в активированное состояние.[38] В зависимости от типа рецептора результирующий эффект на постсинаптическую клетку может быть более сложным возбуждающим, тормозящим или модулирующим. Например, выброс нейромедиатора ацетилхолин при синаптическом контакте между двигательный нейрон и мышечная клетка вызывает быстрое сокращение мышечной клетки.[39] Весь процесс синаптической передачи занимает лишь долю миллисекунды, хотя воздействие на постсинаптическую клетку может длиться намного дольше (даже бесконечно, в тех случаях, когда синаптический сигнал приводит к формированию след памяти ).[12]
| Структура типового химический синапс |
|---|

Существуют буквально сотни различных типов синапсов. На самом деле известно более сотни нейромедиаторов, и многие из них имеют рецепторы нескольких типов.[40] Многие синапсы используют более одного нейромедиатора - обычно синапсы используют один быстродействующий низкомолекулярный нейромедиатор, такой как глутамат или же ГАМК вместе с одним или несколькими пептид нейротрансмиттеры, которые играют медленно действующие модулирующие роли. Молекулярные нейробиологи обычно делят рецепторы на две большие группы: химически закрытые ионные каналы и системы вторичных сообщений. Когда химически управляемый ионный канал активируется, он образует проход, который позволяет определенным типам ионов проходить через мембрану. В зависимости от типа иона действие на клетку-мишень может быть возбуждающим или тормозящим. Когда активируется вторая система обмена сообщениями, она запускает каскад молекулярных взаимодействий внутри клетки-мишени, которые в конечном итоге могут вызывать широкий спектр сложных эффектов, таких как повышение или снижение чувствительности клетки к стимулам или даже изменение транскрипция гена.
Согласно правилу, называемому Принцип Дейла, у которого есть только несколько известных исключений, нейрон выделяет одни и те же нейротрансмиттеры во всех своих синапсах.[41] Однако это не означает, что нейрон оказывает одинаковый эффект на все свои мишени, потому что эффект синапса зависит не от нейромедиатора, а от рецепторов, которые он активирует.[38] Поскольку разные мишени могут (и часто используют) разные типы рецепторов, нейрон может оказывать возбуждающее действие на один набор клеток-мишеней, ингибирующее действие на другие и сложные модулирующие эффекты на другие. Тем не менее бывает, что два наиболее широко используемых нейромедиатора, глутамат и ГАМК, каждый имеет в значительной степени согласованные эффекты. Глутамат имеет несколько широко распространенных типов рецепторов, но все они являются возбуждающими или модулирующими. Точно так же у ГАМК есть несколько широко распространенных типов рецепторов, но все они ингибирующие.[42] Из-за этой последовательности глутаматергические клетки часто называют «возбуждающими нейронами», а ГАМКергические клетки - «тормозящими нейронами». Строго говоря, это злоупотребление терминологией - возбуждающими и тормозящими являются рецепторы, а не нейроны, - но это часто наблюдается даже в научных публикациях.
Одно очень важное подмножество синапсов способно формировать следы памяти посредством длительных зависимых от активности изменений синаптической силы.[43] Самая известная форма нейронной памяти - это процесс, называемый долгосрочное потенцирование (сокращенно LTP), который работает в синапсах, которые используют нейротрансмиттер глутамат действует на особый тип рецептора, известный как Рецептор NMDA.[44] Рецептор NMDA обладает «ассоциативным» свойством: если обе клетки, участвующие в синапсе, активируются примерно в одно и то же время, открывается канал, который позволяет кальцию течь в клетку-мишень.[45] Поступление кальция запускает второй каскад мессенджеров, который в конечном итоге приводит к увеличению количества рецепторов глутамата в клетке-мишени, тем самым увеличивая эффективную силу синапса. Это изменение силы может длиться несколько недель или дольше. С момента открытия LTP в 1973 году было обнаружено много других типов следов синаптической памяти, включая увеличение или уменьшение синаптической силы, которые вызываются различными условиями и сохраняются в течение различных периодов времени.[44] В система вознаграждений, который, например, усиливает желаемое поведение, зависит от вариантной формы LTP, которая обусловлена дополнительным входом, поступающим из пути передачи сигналов вознаграждения, который использует дофамин как нейротрансмиттер.[46] Все эти формы синаптической модифицируемости, взятые вместе, приводят к нейронная пластичность то есть способность нервной системы приспосабливаться к изменениям окружающей среды.
Нейронные схемы и системы
Основная нейронная функция посылки сигналов другим клеткам включает способность нейронов обмениваться сигналами друг с другом. Сети образованные взаимосвязанными группами нейронов, способны выполнять широкий спектр функций, включая обнаружение признаков, генерацию паттернов и синхронизацию,[47] и существует множество возможных способов обработки информации. Уоррен МакКаллох и Уолтер Питтс показал в 1943 г., что даже искусственные нейронные сети сформированные из сильно упрощенной математической абстракции нейрона, способны универсальное вычисление.[48]

Исторически сложилось так, что в течение многих лет преобладающим взглядом на функцию нервной системы была функция, ассоциирующая со стимулом и реакцией.[49] Согласно этой концепции, нейронная обработка начинается со стимулов, которые активируют сенсорные нейроны, производя сигналы, которые распространяются через цепочки связей в спинном и головном мозге, что в конечном итоге приводит к активации моторных нейронов и тем самым к сокращению мышц, то есть к явным ответам. Декарт считал, что все поведение животных и большинство поведения людей можно объяснить в терминах цепей стимул-реакция, хотя он также считал, что высшие когнитивные функции, такие как язык, нельзя объяснить механистически.[50] Чарльз Шеррингтон в своей влиятельной книге 1906 г. Интегративное действие нервной системы,[49] разработал концепцию механизмов стимула-реакции более подробно, и Бихевиоризм, школа мысли, которая доминировала Психология в середине 20-го века пытались объяснить каждый аспект человеческого поведения в терминах «стимул-реакция».[51]
Однако экспериментальные исследования электрофизиология, начиная с начала 20 века и достигнув высокой продуктивности к 1940-м годам, показали, что нервная система содержит множество механизмов для поддержания возбудимость клеток и создание паттернов активности внутренне, не требуя внешнего стимула.[52] Было обнаружено, что нейроны способны производить регулярные последовательности потенциалов действия или последовательности всплесков даже в полной изоляции.[53] Когда внутренне активные нейроны соединяются друг с другом в сложные цепи, возможности для генерации сложных временных паттернов становятся намного более обширными.[47] Современная концепция рассматривает функцию нервной системы частично с точки зрения цепочек «стимул-ответ», а частично с точки зрения внутренне генерируемых паттернов активности - оба типа активности взаимодействуют друг с другом, создавая полный репертуар поведения.[54]
Рефлексы и другие схемы стимул-реакция

Самый простой тип нейронной цепи - это рефлекторная дуга, который начинается с сенсорный входом и заканчивается моторным выходом, проходящим через последовательность нейронов, соединенных в серии.[55] Это может быть отражено в «рефлексе отхода», когда рука дергается назад после прикосновения к горячей плите. Схема начинается с Рецепторы чувств в коже, которые активируются опасными уровнями тепла: особый тип молекулярной структуры, встроенной в мембрану, заставляет тепло изменять электрическое поле через мембрану. Если изменение электрического потенциала достаточно велико, чтобы преодолеть заданный порог, оно вызывает потенциал действия, который передается по аксону рецепторной клетки в спинной мозг. Там аксон устанавливает возбуждающие синаптические контакты с другими клетками, некоторые из которых проецируются (посылают аксональный сигнал) в ту же область спинного мозга, другие - в головной мозг. Одна цель - это набор позвоночника интернейроны который проецируется на двигательные нейроны, управляющие мышцами рук. Интернейроны возбуждают мотонейроны, и, если возбуждение достаточно сильное, некоторые из мотонейронов генерируют потенциалы действия, которые перемещаются по их аксонам до точки, где они устанавливают возбуждающие синаптические контакты с мышечными клетками. Возбуждающие сигналы вызывают сокращение мышечных клеток, в результате чего углы суставов в руке изменяются, оттягивая руку.
В действительности эта простая схема подвержена многочисленным сложностям.[55] Хотя для самых простых рефлексы есть короткие нейронные пути от сенсорного нейрона к двигательному нейрону, есть также другие соседние нейроны, которые участвуют в цепи и модулируют реакцию. Кроме того, есть проекции от головного мозга к спинному мозгу, которые способны усиливать или подавлять рефлекс.
Хотя простейшие рефлексы могут быть опосредованы цепями, полностью лежащими в спинном мозге, более сложные ответы зависят от обработки сигналов в головном мозге.[56] Например, когда объект на периферии поля зрения движется, и человек смотрит на него, инициируются многие этапы обработки сигнала. Первоначальная сенсорная реакция сетчатки глаза и конечная двигательная реакция глазодвигательных ядер ствола мозга не так уж и отличаются от таковых при простом рефлексе, но промежуточные стадии совершенно разные. Вместо одно- или двухступенчатой цепочки обработки зрительные сигналы проходят, возможно, дюжину стадий интеграции, включая таламус, кору головного мозга, базальные ганглии, верхний бугорок, мозжечок и несколько ядер ствола мозга. Эти области выполняют функции обработки сигналов, которые включают обнаружение функции, перцептивный анализ, отзыв памяти, принимать решение, и двигательное планирование.[57]
Обнаружение функции это способность извлекать биологически значимую информацию из комбинаций сенсорных сигналов.[58] в зрительная система, например, сенсорные рецепторы в сетчатка глаза только индивидуально способны обнаруживать «световые точки» во внешнем мире.[59] Зрительные нейроны второго уровня получают входные данные от групп первичных рецепторов, нейроны более высокого уровня получают входные данные от групп нейронов второго уровня и т. Д., Образуя иерархию этапов обработки. На каждом этапе важная информация извлекается из ансамбля сигналов, а неважная информация отбрасывается. К концу процесса входные сигналы, представляющие «световые точки», были преобразованы в нейронное представление объектов окружающего мира и их свойств. Самая сложная сенсорная обработка происходит внутри головного мозга, но извлечение сложных функций также происходит в спинном мозге и в периферических органах чувств, таких как сетчатка.
Генерация внутреннего шаблона
Хотя механизмы стимул-реакция легче всего понять, нервная система также способна управлять телом способами, не требующими внешнего раздражителя, посредством генерируемых внутри ритмов активности. Из-за разнообразия чувствительных к напряжению ионных каналов, которые могут быть встроены в мембрану нейрона, многие типы нейронов способны, даже изолированно, генерировать ритмические последовательности потенциалов действия или ритмические чередования между высокоскоростным взрывом и покоем. . Когда нейроны, которые по своей природе ритмичны, связаны друг с другом возбуждающими или тормозящими синапсами, результирующие сети способны к широкому разнообразию динамического поведения, включая аттрактор динамика, периодичность и даже хаос. Сеть нейронов, которая использует свою внутреннюю структуру для генерации структурированного во времени вывода, не требуя соответствующего стимула, структурированного во времени, называется сетью. центральный генератор шаблонов.
Генерация внутреннего шаблона работает в широком диапазоне временных масштабов, от миллисекунд до часов и более. Один из наиболее важных типов темпорального паттерна - это циркадная ритмичность - то есть ритмичность с периодом примерно 24 часа. У всех исследованных животных наблюдаются циркадные колебания нейронной активности, которые контролируют циркадные изменения в поведении, например, цикл сна и бодрствования. Экспериментальные исследования 1990-х годов показали, что циркадные ритмы генерируются «генетическими часами», состоящими из особого набора генов, уровень экспрессии которых повышается и понижается в течение дня. Такие разные животные, как насекомые и позвоночные, имеют сходную генетическую систему часов. На циркадные часы влияет свет, но они продолжают работать, даже когда уровень освещенности поддерживается постоянным и нет других внешних сигналов времени суток. Гены часов экспрессируются во многих частях нервной системы, а также во многих периферических органах, но у млекопитающих все эти «тканевые часы» поддерживаются синхронно с помощью сигналов, исходящих от главного хронометриста в крошечной части мозга, называемой то супрахиазматическое ядро.
Зеркальные нейроны
А зеркальный нейрон это нейрон, который пожары и когда животное действует, и когда животное наблюдает за тем же действием, выполняемым другим.[60][61][62] Таким образом, нейрон «отражает» поведение другого, как если бы наблюдатель действовал сам. Такие нейроны непосредственно наблюдались в примат разновидность.[63] Было показано, что птицы обладают имитационным резонансным поведением, а неврологические данные свидетельствуют о наличии некоторой формы зеркальной системы.[63][64] У людей активность мозга, соответствующая активности зеркальных нейронов, была обнаружена в премоторная кора, то дополнительная моторная зона, то первичная соматосенсорная кора и нижняя теменная кора.[65] Функция зеркальной системы является предметом многих предположений. Многие исследователи в области когнитивной нейробиологии и когнитивной психологии считают, что эта система обеспечивает физиологический механизм взаимодействия восприятия и действия (см. общая теория кодирования ).[62] Они утверждают, что зеркальные нейроны могут быть важны для понимания действий других людей и для обучения новым навыкам путем имитации. Некоторые исследователи также предполагают, что зеркальные системы могут имитировать наблюдаемые действия и, таким образом, способствовать теория разума навыки,[66][67] в то время как другие связывают зеркальные нейроны с язык способности.[68] Однако на сегодняшний день не было предложено общепринятых нейронных или вычислительных моделей, чтобы описать, как активность зеркальных нейронов поддерживает когнитивные функции, такие как имитация.[69] Есть нейробиологи, которые предупреждают, что заявления о роли зеркальных нейронов не подтверждаются адекватными исследованиями.[70][71]
Разработка
У позвоночных ориентиров эмбрионального нейронное развитие включить рождение и дифференциация из нейроны из стволовая клетка предшественники, миграция незрелых нейронов от мест их рождения в эмбрионе до их конечных положений, рост аксоны от нейронов и руководство подвижного конус роста через эмбрион к постсинаптическим партнерам, поколение синапсы между этими аксонами и их постсинаптическими партнерами, и, наконец, пожизненное изменения в синапсах, которые, как считается, лежат в основе обучения и памяти.[72]
Все двустворчатые животные на ранней стадии развития образуют гаструла, который поляризован, с одним концом, называемым полюс животного а другой растительный полюс. Гаструла имеет форму диска с тремя слоями клеток, внутренний слой называется энтодерма, который дает начало подкладке большинства внутренних органов, средний слой, называемый мезодерма, который дает рост костей и мускулов, а также внешний слой, называемый эктодерма, дающий начало коже и нервной системе.[73]
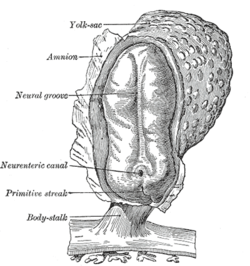 Человеческий эмбрион с нервной бороздкой |  Четыре стадии развития нервной трубки у эмбриона человека |
У позвоночных первым признаком нервной системы является появление тонкой полосы клеток по центру спины, называемой нервная пластинка. Внутренняя часть нервной пластинки (по средней линии) предназначена стать Центральная нервная система (ЦНС), внешняя часть периферическая нервная система (ПНС). По мере развития появляется складка, называемая нервная борозда появляется по средней линии. Эта складка углубляется, а затем закрывается наверху. На этом этапе будущая ЦНС выглядит как цилиндрическая структура, называемая нервная трубка, тогда как будущая ПНС выглядит как две полосы ткани, называемые нервный гребень, проходит над нервной трубкой в продольном направлении. Последовательность стадий от нервной пластинки до нервной трубки и нервного гребня известна как нейруляция.
В начале 20 века серия знаменитых экспериментов Ханса Спеманна и Хильды Мангольд показала, что образование нервной ткани «индуцируется» сигналами от группы мезодермальных клеток, называемых регион организатора.[72] Однако на протяжении десятилетий природа нервная индукция побеждал все попытки понять это, пока, наконец, в 1990-х годах проблема не была решена с помощью генетических подходов. Для индукции нервной ткани требуется ингибирование гена так называемого костный морфогенетический белок, или BMP. В частности, белок BMP4 похоже, причастен. Два белка под названием Noggin и Chordin оба секретируются мезодермой и способны ингибировать BMP4 и тем самым вызывать превращение эктодермы в нервную ткань. Похоже, что аналогичный молекулярный механизм задействован для очень разных типов животных, включая членистоногих, а также позвоночных. Однако у некоторых животных существует другой тип молекул, называемый Фактор роста фибробластов или FGF также может играть важную роль в индукции.
Индукция нервных тканей вызывает образование нервных клеток-предшественников, называемых нейробласты.[74] У дрозофилы нейробласты делятся асимметрично, так что один продукт представляет собой «материнскую клетку ганглия» (GMC), а другой - нейробласт. GMC делится один раз, давая начало паре нейронов или паре глиальных клеток. В целом нейробласт способен генерировать неопределенное количество нейронов или глии.
Как показано в исследовании 2008 года, один общий для всех фактор двусторонний организмов (включая людей) - это семейство секретных сигнальные молекулы называется нейротрофины которые регулируют рост и выживание нейроны.[75] Zhu et al. идентифицировали DNT1, первый нейротрофин, обнаруженный в мухи. DNT1 имеет структурное сходство со всеми известными нейротрофинами и является ключевым фактором в судьбе нейронов в Дрозофила. Поскольку нейротрофины в настоящее время идентифицированы как у позвоночных, так и у беспозвоночных, эти данные свидетельствуют о том, что нейротрофины присутствовали у предков, общих для двусторонних организмов, и могут представлять общий механизм формирования нервной системы.
Патология

Центральная нервная система защищена основными физическими и химическими барьерами. Физически головной и спинной мозг окружены жесткими менингеальный мембраны и заключены в костях череп и позвоночник, которые вместе образуют прочный физический щит. Химически головной и спинной мозг изолированы гематоэнцефалический барьер, который предотвращает попадание большинства химических веществ из кровотока внутрь ЦНС. Эти защиты делают ЦНС менее уязвимой во многих отношениях, чем ПНС; обратная сторона, однако, заключается в том, что повреждение ЦНС имеет тенденцию иметь более серьезные последствия.
Хотя нервы, как правило, лежат глубоко под кожей, за исключением нескольких мест, таких как локтевой нерв вблизи локтевого сустава они все еще относительно подвержены физическим повреждениям, которые могут вызвать боль, потерю чувствительности или потерю мышечного контроля. Повреждение нервов также может быть вызвано отеком или ушибами в местах, где нерв проходит через плотный костный канал, как это происходит в синдром запястного канала. Если нерв полностью перерезан, он часто возродить, но для длинных нервов этот процесс может занять месяцы. Помимо физического урона, периферическая невропатия может быть вызвано многими другими медицинскими проблемами, включая генетические состояния, метаболические состояния, такие как сахарный диабет воспалительные состояния, такие как Синдром Гийена-Барре, авитаминоз, инфекционные заболевания, такие как проказа или же опоясывающий лишай, или отравление токсинами, такими как тяжелые металлы. Многие случаи не имеют причины, которую можно установить, и называются идиопатический. Также возможно, что нервы временно потеряют функцию, что приведет к онемению как жесткости - частые причины включают механическое давление, падение температуры или химическое взаимодействие с местный анестетик наркотики, такие как лидокаин.
Физическое повреждение спинного мозга может привести к потеря чувствительности или движения. Если травма позвоночника не вызывает ничего хуже, чем отек, симптомы могут быть временными, но если нервные волокна в позвоночнике фактически разрушены, потеря функции обычно необратима. Экспериментальные исследования показали, что спинномозговые нервные волокна пытаются вырасти таким же образом, как и нервные волокна, но в спинном мозге разрушение ткани обычно приводит к образованию рубцовой ткани, через которую отрастающие нервы не могут проникнуть.
Смотрите также
Рекомендации
- ^ Тортора, Дж. Дж., Дерриксон, Б. (2016). Основы анатомии и физиологии (15-е изд.). Дж. Вили. ISBN 978-1-119-34373-8.CS1 maint: несколько имен: список авторов (связь)
- ^ "Нервная система". Колумбийская энциклопедия. Издательство Колумбийского университета.
- ^ Herculano-Houzel S, Avelino-de-Souza K и др. (2014). «Слоновий мозг в цифрах». Фронт нейроанат. 8: 46. Дои:10.3389 / fnana.2014.00046. ЧВК 4053853. PMID 24971054.
- ^ а б c d е ж Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 2: Нервные клетки и поведение». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Палец S (2001). «Глава 1: Мозг в древности». Истоки нейробиологии: история исследований функций мозга. Oxford Univ. Нажмите. ISBN 978-0-19-514694-3.
- ^ Палец, стр. 43–50.
- ^ Николецис Майкл М. (2010) Поведенческая и нервная пластичность. ISBN 978-1-4537-8945-2
- ^ а б Сакарья О., Армстронг К.А., Адамска М. и др. (2007). Vosshall L (ред.). «Постсинаптический каркас у истоков животного мира». PLOS ONE. 2 (6): e506. Bibcode:2007PLoSO ... 2..506S. Дои:10.1371 / journal.pone.0000506. ЧВК 1876816. PMID 17551586.
- ^ а б Рупперт EE, Fox RS, Barnes RD (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.111–124. ISBN 0-03-025982-7.
- ^ а б Балавойн Г (2003). «Сегментированная урбилатерия: проверяемый сценарий». Int Comp Биология. 43 (1): 137–147. Дои:10.1093 / icb / 43.1.137. PMID 21680418.
- ^ Ортега-Эрнандес, Хавьер (29 февраля 2016 г.), Окаменелость нервной системы возрастом 500 миллионов лет проливает свет на эволюцию животных, The Conversation US, Inc., получено 6 марта 2016
- ^ а б c Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 4: Цитология нейронов». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ а б Аллен Нью-Джерси, Баррес Б.А. (2009). «Неврология: Глия - больше, чем клей для мозга». Природа. 457 (7230): 675–677. Bibcode:2009Натура.457..675А. Дои:10.1038 / 457675a. PMID 19194443. S2CID 205044137.
- ^ Азеведо Ф.А., Карвалью Л.Р., Гринберг Л.Т. и др. (2009). «Равное количество нейрональных и ненейрональных клеток делает человеческий мозг изометрически увеличенным мозгом приматов». J. Comp. Neurol. 513 (5): 532–541. Дои:10.1002 / cne.21974. PMID 19226510. S2CID 5200449.
- ^ а б Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 17: Анатомическая организация центральной нервной системы». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Standring, Сьюзан (главный редактор) (2005). Анатомия Грея (39-е изд.). Эльзевьер Черчилль Ливингстон. стр.233–234. ISBN 978-0-443-07168-3.
- ^ Хаббард Д.И. (1974). Периферическая нервная система. Пленум Пресс. п. vii. ISBN 978-0-306-30764-5.
- ^ Purves D, Augustine GJ, Fitzpatrick D, Hall WC, LaMantia AS, McNamara JO, White LE (2008). Неврология. 4-е изд. Sinauer Associates. С. 15–16.
- ^ "ганглий " в Медицинский словарь Дорланда
- ^ Афифи А.К. (июль 1994 г.). «Базальные ганглии: функциональная анатомия и физиология. Часть 1». J. Child Neurol. 9 (3): 249–260. Дои:10.1177/088307389400900306. PMID 7930403. S2CID 13045090.
- ^ Джейкобс Д.К., Наканиши Н., Юань Д., Камара А., Николс С.А., Хартенштейн В. и др. (2007). «Эволюция сенсорных структур у базальных многоклеточных животных». Интегр Комп Биол. 47 (5): 712–723. Дои:10.1093 / icb / icm094. PMID 21669752.
- ^ Санес Д.Х., Рех Т.А., Харрис В.А. (2006). Развитие нервной системы. Академическая пресса. стр.3 –4. ISBN 978-0-12-618621-5.
- ^ Гайсен А (2003). «Происхождение и эволюция нервной системы». Int. J. Dev. Биол. 47 (7–8): 555–562. CiteSeerX 10.1.1.511.5106. PMID 14756331.
- ^ Эрвин Д.Х., Дэвидсон Э.Х. (июль 2002 г.). «Последний общий предок двуногих». Разработка. 129 (13): 3021–3032. PMID 12070079.
- ^ Бурлат С.Дж., Юлиусдоттир Т., Лоу С.Дж. и др. (Ноябрь 2006 г.). «Филогения Deuterostome выявляет монофилетические хордовые и новый тип Xenoturbellida». Природа. 444 (7115): 85–88. Bibcode:2006Натура 444 ... 85Б. Дои:10.1038 / природа05241. PMID 17051155. S2CID 4366885.
- ^ Lichtneckert R, Reichert H (май 2005 г.). "Взгляд на мозг urbilaterian: консервативные механизмы формирования генетического паттерна в развитии мозга насекомых и позвоночных". Наследственность. 94 (5): 465–477. Дои:10.1038 / sj.hdy.6800664. PMID 15770230.
- ^ ADEY WR (февраль 1951 г.). «Нервная система дождевого червя Megascolex». J. Comp. Neurol. 94 (1): 57–103. Дои:10.1002 / cne.900940104. PMID 14814220. S2CID 30827888.
- ^ а б «Книга червя: Спецификация нервной системы».
- ^ Чапман РФ (1998). «Глава 20: Нервная система». Насекомые: строение и функции. Издательство Кембриджского университета. стр.533–568. ISBN 978-0-521-57890-5.
- ^ Чепмен, стр. 546
- ^ а б Хойл Г., Виерсма, Калифорния (1977). Выявленные нейроны и поведение членистоногих. Пленум Пресс. ISBN 978-0-306-31001-0.
- ^ Штейн ПСЖ (1999). Нейроны, сети и моторное поведение. MIT Press. С. 38–44. ISBN 978-0-262-69227-4.
- ^ Штейн, стр. 112
- ^ Симмонс П.Дж., Янг Д. (1999). Нервные клетки и поведение животных. Издательство Кембриджского университета. п.43. ISBN 978-0-521-62726-9.
- ^ Грей ПО (2006). Психология (5-е изд.). Макмиллан. п.170. ISBN 978-0-7167-7690-1.
- ^ а б Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 9: Распространенная сигнализация: потенциал действия». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Хормузди С.Г., Филиппов М.А., Митропулу Г. и др. (2004). «Электрические синапсы: динамическая сигнальная система, которая формирует активность нейронных сетей». Биохим. Биофиз. Acta. 1662 (1–2): 113–137. Дои:10.1016 / j.bbamem.2003.10.023. PMID 15033583.
- ^ а б c Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 10: Обзор синаптической передачи». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 11: Передача сигналов в нервно-мышечном синапсе». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 15: Нейротрансмиттеры». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Страта П., Харви Р. (1999). «Принцип Дейла». Brain Res. Бык. 50 (5–6): 349–350. Дои:10.1016 / S0361-9230 (99) 00100-8. PMID 10643431. S2CID 29406273.
- ^ Есть ряд исключительных ситуаций, в которых было обнаружено, что ГАМК оказывает возбуждающее действие, в основном на раннем этапе развития. Для обзора см. Марти А., Льяно I. (июнь 2005 г.). «Возбуждающие эффекты ГАМК в установленных сетях мозга». Тенденции Neurosci. 28 (6): 284–289. Дои:10.1016 / j.tins.2005.04.003. PMID 15927683. S2CID 40022079.
- ^ Парадизо М.А., Медведь М.Ф., Коннорс Б.В. (2007). Нейробиология: исследование мозга. Липпинкотт Уильямс и Уилкинс. п.718. ISBN 978-0-7817-6003-4.
- ^ а б Кук SF, Bliss TV (2006). «Пластичность центральной нервной системы человека». Мозг. 129 (Pt 7): 1659–1673. Дои:10.1093 / мозг / awl082. PMID 16672292.
- ^ Bliss TV, Collingridge GL (январь 1993 г.). «Синаптическая модель памяти: долговременная потенциация в гиппокампе». Природа. 361 (6407): 31–39. Bibcode:1993Натура 361 ... 31Б. Дои:10.1038 / 361031a0. PMID 8421494. S2CID 4326182.
- ^ Кауэр Я.А., Маленка RC (ноябрь 2007 г.). «Синаптическая пластичность и зависимость». Nat. Преподобный Neurosci. 8 (11): 844–858. Дои:10.1038 / номер 2234. PMID 17948030. S2CID 38811195.
- ^ а б Даян П., Эбботт Л.Ф. (2005). Теоретическая нейробиология: вычислительное и математическое моделирование нейронных систем. MIT Press. ISBN 978-0-262-54185-5.
- ^ Маккаллох WS, Питтс W (1943). «Логический исчисление идей, присущих нервной деятельности». Бык. Математика. Биофизы. 5 (4): 115–133. Дои:10.1007 / BF02478259.
- ^ а б Шеррингтон CS (1906). Интегративное действие нервной системы. Скрибнер.
- ^ Декарт Р. (1989). Страсти души. Восс С. Хакетт. ISBN 978-0-87220-035-7.
- ^ Баум WM (2005). Понимание бихевиоризма: поведение, культура и эволюция. Блэквелл. ISBN 978-1-4051-1262-8.
- ^ Piccolino M (ноябрь 2002 г.). «Пятьдесят лет эпохи Ходжкина-Хаксли». Тенденции Neurosci. 25 (11): 552–553. Дои:10.1016 / S0166-2236 (02) 02276-2. PMID 12392928. S2CID 35465936.
- ^ Джонстон Д., Ву С.М. (1995). Основы клеточной нейрофизиологии. MIT Press. ISBN 978-0-262-10053-3.
- ^ Симмонс П.Дж., Янг Д. (1999). «Гл 1. Введение». Нервные клетки и поведение животных. Cambridge Univ. Нажмите. ISBN 978-0-521-62726-9.
- ^ а б Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 36: Спинальные рефлексы». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 38: Добровольное движение». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 39: Контроль взгляда». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 21: Кодирование сенсорной информации». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 25: Построение визуального образа». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Риццолатти, Джакомо; Крейгеро, Лейла (2004). «Зеркало-нейронная система» (PDF). Ежегодный обзор нейробиологии. 27: 169–192. Дои:10.1146 / annurev.neuro.27.070203.144230. PMID 15217330.
- ^ Кейзерс, Кристиан (2010). «Зеркальные нейроны» (PDF). Текущая биология. 19 (21): R971–973. Дои:10.1016 / j.cub.2009.08.026. PMID 19922849. S2CID 12668046. Архивировано из оригинал (PDF) 19 января 2013 г.
- ^ а б Кейзерс, Кристиан (23 июня 2011 г.). Эмпатический мозг. Kindle.
- ^ а б Риццолатти, Джакомо; Фадига, Лучано (1999). «Резонансное поведение и зеркальные нейроны». Italiennes de Biologie. 137: 85–100. PMID 10349488.
- ^ Акинс, Хана; Кляйн, Эдвард (2002). «Имитационное обучение японского перепела с использованием процедуры двунаправленного контроля». Обучение и поведение животных. 30 (3): 275–281. Дои:10.3758 / bf03192836. PMID 12391793.
- ^ Моленбергс П., Каннингтон Р., Мэттингли Дж. (Июль 2009 г.). «Участвует ли система зеркальных нейронов в имитации? Краткий обзор и метаанализ». Неврология и биоповеденческие обзоры. 33 (1): 975–980. Дои:10.1016 / j.neubiorev.2009.03.010. PMID 19580913. S2CID 25620637.
- ^ Кейзерс, Кристиан; Газзола, Валерия (2006). «Прогресс в исследованиях мозга» (PDF). Bcn-nic.nl. Архивировано из оригинал (PDF) 30 июня 2007 г.
- ^ Михаил Арбиб, Гипотеза зеркальной системы. Связь языка с теорией разума В архиве 29 марта 2009 г. Wayback Machine, 2005, получено 17 февраля 2006 г.
- ^ Теорет, Гюго; Паскуаль-Леоне, Альваро (2002). «Приобретение языка: делай, как слышишь». Текущая биология. 12 (21): R736 – R737. Дои:10.1016 / S0960-9822 (02) 01251-4. PMID 12419204. S2CID 12867585.
- ^ Динштейн I, Томас С., Берманн М., Хигер Д.Д. (2008). «Зеркало природы». Curr Biol. 18 (1): R13 – R18. Дои:10.1016 / j.cub.2007.11.004. ЧВК 2517574. PMID 18177704.
- ^ Хикок, Г. (21 июля 2009 г.). «Восемь проблем для понимания действия теории зеркального нейрона у обезьян и людей». Журнал когнитивной неврологии. 21 (7): 1229–1243. Дои:10.1162 / jocn.2009.21189. ЧВК 2773693. PMID 19199415.
- ^ Привет, Сесилия (2009). "Откуда берутся зеркальные нейроны?" (PDF). Неврология и биоповеденческие обзоры. Архивировано из оригинал (PDF) 26 апреля 2012 г.. Получено 14 января 2015.
- ^ а б Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 52: индукция и формирование паттернов нервной системы». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Санес DH, Рех TH, Харрис WA (2006). "Глава 1, Нейронная индукция". Развитие нервной системы. Elsevier Academic Press. ISBN 978-0-12-618621-5.
- ^ Кандел Э. Р., Шварц Дж. Х., Джессел Т. М., ред. (2000). «Глава 53: Формирование и выживание нервных клеток». Принципы нейронологии. McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ^ Чжу Б., Пеннак Дж., Маккуилтон П., Фореро М. Г., Мизугучи К., Сатклифф Б., Гу Си Джей, Фентон Дж. К., Идальго А. (ноябрь 2008 г.). Бейт М (ред.). «Нейротрофины дрозофилы раскрывают общий механизм формирования нервной системы». ПЛОС Биол. 6 (11): e284. Дои:10.1371 / journal.pbio.0060284. ЧВК 2586362. PMID 19018662. Архивировано из оригинал 5 февраля 2009 г.
дальнейшее чтение
- Нервная система Уильям Э. Скэггс, Scholarpedia
внешняя ссылка
| Библиотечные ресурсы о Нервная система |
 Нервная система в Викиучебнике (человек)
Нервная система в Викиучебнике (человек)- Нервная система в Викиучебнике (кроме людей)
- Домашняя страница проекта "Человеческий мозг"