РНК-интерференция - RNA interference
РНК-интерференция (РНКи) - это биологический процесс, в котором РНК молекулы подавлять экспрессию генов или перевод, нейтрализуя целевые мРНК молекулы.[1] Исторически RNAi была известна под другими названиями, включая совместное подавление, посттранскрипционное молчание генов (PTGS) и подавление. Детальное изучение каждого из этих, казалось бы, различных процессов выяснило, что идентичность этих явлений на самом деле была РНКи. Эндрю Файер и Крейг С. Мелло разделила 2006 Нобелевская премия по физиологии и медицине за их работу по интерференции РНК в нематода червь Caenorhabditis elegans, который они опубликовали в 1998 году. С момента открытия РНКи и ее регуляторных потенциалов стало очевидно, что РНКи обладают огромным потенциалом в подавлении желаемых генов. РНКи теперь известны как точные, эффективные, стабильные и лучше, чем антисмысловая терапия для подавления генов.[2] Тем не мение, антисмысловая РНК продуцируемый внутриклеточно экспрессирующим вектором, можно разработать и найти применение в качестве новых терапевтических агентов.[3]
Два типа малых рибонуклеиновая кислота (РНК) молекулы - микроРНК (миРНК) и малая интерферирующая РНК (миРНК ) - центральное место в РНК-интерференции. РНК являются непосредственными продуктами генов, и эти небольшие РНК могут направлять ферментные комплексы на разрушение. информационная РНК (мРНК) и, таким образом, снижают их активность за счет предотвращения трансляции за счет посттранскрипционного молчания генов. Более того, транскрипция может быть ингибирована с помощью механизма пре-транскрипционного сайленсинга РНК-интерференции, посредством которого ферментный комплекс катализирует метилирование ДНК в геномных положениях, комплементарных комплексной миРНК или миРНК. РНК-интерференция играет важную роль в защите клеток от паразитарных болезней. нуклеотид последовательности - вирусы и транспозоны. Это также влияет разработка.
Путь РНКи встречается во многих эукариоты, включая животных, и инициируется ферментом Дайсер, который долго рассекает двухцепочечная РНК (дцРНК) молекулы на короткие двухцепочечные фрагменты по ~ 21 нуклеотид миРНК. Каждый миРНК разматывается на две одноцепочечные РНК (оцРНК), цепь-пассажира и цепь-проводник. Пассажирская прядь разрушается, и направляющая прядь включается в РНК-индуцированный комплекс сайленсинга (RISC). Наиболее хорошо изученным результатом является посттранскрипционное молчание генов, которое происходит, когда направляющая цепь соединяется с комплементарной последовательностью в молекуле информационной РНК и вызывает расщепление посредством Аргонавт 2 (Ago2), каталитический компонент RISC. У некоторых организмов этот процесс распространяется системно, несмотря на изначально ограниченные молярные концентрации миРНК.
RNAi - ценный исследовательский инструмент как в культура клеток И в живые организмы, потому что синтетическая дцРНК, введенная в клетки, может селективно и надежно индуцировать супрессию конкретных представляющих интерес генов. РНКи можно использовать для крупномасштабных экранов, которые систематически отключают каждый ген в клетке, что может помочь идентифицировать компоненты, необходимые для конкретного клеточного процесса или события, такого как деление клеток. Путь также используется как практический инструмент в биотехнология, лекарство и инсектициды.[4]
Сотовый механизм

РНКи является РНК-зависимой подавление гена процесс, который контролируется комплексом РНК-индуцированного сайленсинга (RISC) и инициируется короткими двухцепочечными молекулами РНК в цитоплазме клетки, где они взаимодействуют с каталитическим компонентом RISC аргонавт.[6] Когда дцРНК является экзогенной (возникшей в результате заражения вирусом с геномом РНК или лабораторных манипуляций), РНК импортируется непосредственно в цитоплазма и расщеплен на короткие фрагменты Дайсером. Инициирующая дцРНК также может быть эндогенной (происходить из клетки), как в пре-микроРНК, экспрессируемых из РНК-кодирующие гены в геноме. Первичные транскрипты таких генов сначала обрабатываются для формирования характерных стебель-петля структура пре-миРНК в ядро, затем экспортируется в цитоплазму. Таким образом, два пути дцРНК, экзогенный и эндогенный, сходятся в RISC.[7]
Экзогенная дцРНК инициирует РНКи путем активации рибонуклеаза белок Dicer[8] который связывает и расщепляет двухцепочечные РНК (дцРНК) в растениях или короткие шпильчатые РНК (кшРНК) у человека с образованием двухцепочечных фрагментов из 20-25 пар оснований с 2-нуклеотидным выступом на 3'-конце.[9] Биоинформатика исследования геномов множества организмов показывают, что такая длина максимизирует специфичность гена-мишени и сводит к минимуму неспецифические эффекты.[10] Эти короткие двухцепочечные фрагменты называются малыми интерферирующими РНК (миРНК ). Эти миРНК затем разделяются на отдельные нити и интегрируются в активный RISC с помощью RISC-Loading Complex (RLC). RLC включает Dicer-2 и R2D2 и имеет решающее значение для объединения Ago2 и RISC.[11] TATA-связывающий белок-связанный фактор 11 (TAF11) собирает RLC, способствуя тетрамеризации Dcr-2-R2D2, что увеличивает сродство связывания с siRNA в 10 раз. Ассоциация с TAF11 преобразует комплекс R2-D2-инициатор (RDI) в RLC.[12] R2D2 несет тандемные двухцепочечные РНК-связывающие домены для распознавания термодинамически стабильного конца миРНК дуплексы, тогда как Дайсер-2 другая менее устойчивая конечность. Загрузка асимметрична: домен MID Ago2 распознает термодинамически стабильный конец siRNA. Следовательно, «пассажирская» (смысловая) цепь, 5'-конец которой отбрасывается MID, выбрасывается, в то время как сохраненная «направляющая» (антисмысловая) цепь взаимодействует с AGO с образованием RISC.[11]
После интеграции в RISC, миРНК пары оснований к своей мРНК-мишени и расщепляют ее, тем самым предотвращая ее использование в качестве перевод шаблон.[13] В отличие от миРНК, нагруженный miRNA комплекс RISC сканирует цитоплазматические мРНК на предмет потенциальной комплементарности. Вместо деструктивного расщепления (с помощью Ago2) miRNAs скорее нацелены на области 3'-нетранслируемой области (UTR) мРНК, где они обычно связываются с несовершенной комплементарностью, тем самым блокируя доступ рибосом для трансляции.[14]
Экзогенная дцРНК обнаруживается и связывается эффекторным белком, известным как RDE-4 в C. elegans и R2D2 в Дрозофила, что стимулирует активность дайсера.[15] Механизм, обеспечивающий такую специфичность длины, неизвестен, и этот белок связывает только длинные дцРНК.[15]
В C. elegans этот инициирующий ответ усиливается за счет синтеза популяции «вторичных» миРНК во время которого производимый дайсером инициирующий или «первичный» миРНК используются как шаблоны.[16] Эти "второстепенные" миРНК структурно отличаются от производимых дайсером миРНК и, кажется, произведен РНК-зависимая РНК-полимераза (RdRP).[17][18]
МикроРНК

МикроРНК (миРНК) являются геномно закодированный некодирующие РНК которые помогают регулировать экспрессия гена особенно во время разработка.[19] Феномен РНК-интерференции, в широком смысле, включает в себя эндогенно индуцированные эффекты сайленсинга генов с помощью miRNA, а также сайленсинг, запускаемый чужеродной дцРНК. Зрелые миРНК структурно похожи на миРНК продуцируются из экзогенной дцРНК, но до достижения зрелости миРНК должны сначала пройти обширную посттранскрипционная модификация. МикроРНК экспрессируется из гораздо более длинного гена, кодирующего РНК, как первичная стенограмма известный как при-миРНК который обрабатывается, в ядро клетки, к 70-нуклеотиду стебель-петля структура называется пре-миРНК посредством микропроцессорный комплекс. Этот комплекс состоит из РНКаза III фермент под названием Дроша и дцРНК-связывающий белок DGCR8. Часть dsRNA этой пре-miRNA связывается и расщепляется Dicer с образованием зрелой молекулы miRNA, которая может быть интегрирована в комплекс RISC; таким образом, miRNA и миРНК используют одно и то же последующее клеточное оборудование.[20] Во-первых, в EBV была описана кодируемая вирусом miRNA.[21] После этого у вирусов было описано все большее количество микроРНК. VIRmiRNA - это полный каталог вирусных микроРНК, их мишеней и антивирусных miRNA. [22] (см. также ресурс VIRmiRNA: http://crdd.osdd.net/servers/virmirna/).
миРНК полученные из длинных предшественников дцРНК, отличаются от миРНК тем, что миРНК, особенно у животных, обычно имеют неполное спаривание оснований с мишенью и ингибируют трансляцию многих различных мРНК со сходными последовательностями. В отличие, миРНК обычно идеально сочетаются с парами оснований и вызывают расщепление мРНК только в одной конкретной мишени.[23] В Дрозофила и C. elegans, миРНК и миРНК обрабатываются различными белками аргонавта и ферментами-дицерами.[24][25]
Три первичных нетранслируемых области и микроРНК
Три основных непереведенных региона (3'UTRs) из информационные РНК (мРНК) часто содержат регуляторные последовательности, которые посттранскрипционно вызывают интерференцию РНК. Такие 3'-UTR часто содержат оба сайта связывания для микроРНК (miRNA), а также для регуляторных белков. Связываясь со специфическими сайтами в 3'-UTR, miRNA могут снижать экспрессию генов различных мРНК, либо ингибируя трансляцию, либо напрямую вызывая деградацию транскрипта. 3'-UTR также может иметь области сайленсеров, которые связывают репрессорные белки, которые ингибируют экспрессию мРНК.
3'-UTR часто содержит элементы ответа микроРНК (MRE). MRE представляют собой последовательности, с которыми связываются miRNA. Это преобладающие мотивы в 3'-UTR. Среди всех регуляторных мотивов в 3'-UTR (например, включая области сайленсеров) MRE составляют около половины мотивов.
По состоянию на 2014 г. miRBase интернет сайт,[26] архив miRNA последовательности и аннотации, перечисляющие 28 645 записей о 233 биологических видах. Из них 1881 miRNA находились в аннотированных локусах miRNA человека. miRNAs предположительно имеют в среднем около четырехсот мишеней мРНК (влияя на экспрессию нескольких сотен генов).[27] Friedman et al.[27] подсчитали, что> 45000 сайтов-мишеней miRNA в 3'UTR мРНК человека консервативны выше фоновых уровней, и более 60% генов, кодирующих белок человека, находились под селективным давлением для поддержания спаривания с miRNA.
Прямые эксперименты показывают, что одна миРНК может снизить стабильность сотен уникальных мРНК.[28] Другие эксперименты показывают, что одиночная miRNA может подавлять продукцию сотен белков, но эта репрессия часто бывает относительно мягкой (менее чем в 2 раза).[29][30]
Эффекты нарушения регуляции экспрессии генов miRNA кажутся важными при раке.[31] Например, при раке желудочно-кишечного тракта девять миРНК были идентифицированы как эпигенетически изменен и эффективен в подавлении ферментов репарации ДНК.[32]
Эффекты дисрегуляции экспрессии генов miRNA также, по-видимому, важны при нейропсихиатрических расстройствах, таких как шизофрения, биполярное расстройство, большая депрессия, болезнь Паркинсона, болезнь Альцгеймера и расстройства аутистического спектра.[33][34][35]
Активация и катализ RISC
Экзогенная дцРНК обнаруживается и связывается эффекторным белком, известным как RDE-4 в C. elegans и R2D2 в Дрозофила, что стимулирует активность дайсера.[15] Этот белок связывает только длинные дцРНК, но механизм, обеспечивающий специфичность этой длины, неизвестен.[15] Этот связывающий РНК белок затем облегчает перенос расщепленных миРНК к комплексу RISC.[36]
В C. elegans этот инициирующий ответ усиливается за счет синтеза популяции «вторичных» миРНК во время которого производимая дайсером инициирующая или «первичная» миРНК используются как шаблоны.[16] Эти "второстепенные" миРНК структурно отличаются от производимых дайсером миРНК и, кажется, произведен РНК-зависимая РНК-полимераза (RdRP).[17][18]


Активные компоненты комплекса РНК-индуцированного сайленсинга (RISC): эндонуклеазы называемые белками аргонавта, которые расщепляют целевую цепь мРНК дополнительный к их границам миРНК.[6] Поскольку фрагменты, произведенные дайсером, являются двухцепочечными, каждый из них теоретически может давать функциональную миРНК. Однако только одна из двух нитей, известная как направляющая прядь, связывается с белком аргонавта и направляет молчание генов. Другой антинаправляющая прядь или же пассажирский проезд ухудшается во время активации RISC.[37] Хотя сначала считалось, что АТФ -зависимый геликаза разделили эти две нити,[38] процесс оказался АТФ-независимым и осуществляется непосредственно белковыми компонентами RISC.[39][40] Однако in vitro кинетический анализ РНКи в присутствии и в отсутствие АТФ показал, что АТФ может потребоваться для раскручивания и удаления расщепленной цепи мРНК из комплекса RISC после катализа.[41] Направляющая прядь обычно та, чья 5 'конец менее стабильно соединяется со своим дополнением,[42] но на отбор цепи не влияет направление, в котором дисер расщепляет дцРНК перед включением RISC.[43] Вместо этого белок R2D2 может служить фактором дифференциации, связывая более стабильный 5'-конец цепи-пассажира.[44]
Структурная основа связывания РНК с белком аргонавта была исследована Рентгеновская кристаллография привязки домен белка аргонавта, связанного с РНК. Здесь фосфорилированный 5 'конец цепи РНК входит в консервированный базовый поверхность карман и устанавливает контакты через двухвалентный катион (атом с двумя положительными зарядами), например магний и по ароматный штабелирование (процесс, который позволяет более чем одному атому обмениваться электроном, передавая его вперед и назад) между 5'-нуклеотидом в миРНК и консервированный тирозин остаток. Считается, что этот сайт формирует сайт зародышеобразования для связывания миРНК к его мишени мРНК.[45] Анализ ингибирующего эффекта несовпадений на 5'- или 3'-конце направляющей цепи показал, что 5'-конец направляющей цепи, вероятно, отвечает за сопоставление и связывание целевой мРНК, в то время как 3'-конец отвечает за физическое размещение мРНК-мишени в области RISC, благоприятной для расщепления.[41]
Неизвестно, как активированный комплекс RISC находит комплементарные мРНК внутри клетки. Хотя было предложено связать процесс расщепления с перевод, трансляция мРНК-мишени не важна для РНКи-опосредованной деградации.[46] Действительно, РНКи могут быть более эффективными против мишеней мРНК, которые не транслируются.[47] Белки аргонавтов локализуются в определенных областях цитоплазмы, называемых П-тела (также цитоплазматические тельца или тельца GW), которые представляют собой области с высокими скоростями распада мРНК;[48] Активность miRNA также сгруппирована в P-тельцах.[49] Нарушение Р-телец снижает эффективность РНК-интерференции, предполагая, что они являются критическим сайтом в процессе РНКи.[50]
Транскрипционное молчание
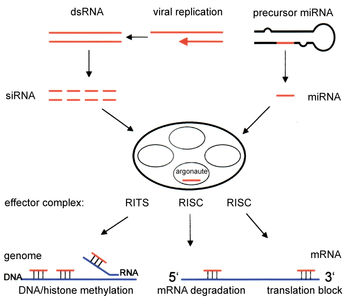
Компоненты пути РНКи используются у многих эукариот для поддержания организации и структуры их геномы. Модификация гистоны и связанная индукция гетерохроматин образование служит для подавления генов пре-транскрипционно;[52] этот процесс называется РНК-индуцированное подавление транскрипции (RITS) и осуществляется комплексом белков, называемым комплексом RITS. В делящиеся дрожжи этот комплекс содержит аргонавт, хромодомен белок Chp1 и белок Tas3 неизвестной функции.[53] Как следствие, для индукции и распространения гетерохроматиновых областей необходимы белки argonaute и RdRP.[54] Действительно, делеция этих генов у делящихся дрожжей С. Помбе нарушает метилирование гистонов и центромера формирование[55] вызывая медленный или остановленный анафаза в течение деление клеток.[56] В некоторых случаях наблюдались сходные процессы, связанные с модификацией гистонов, с активацией транскрипции генов.[57]
Механизм, с помощью которого комплекс RITS индуцирует образование и организацию гетерохроматина, не совсем понятен. Большинство исследований сосредоточено на область спаривания в делящихся дрожжах, которые не могут быть репрезентативными для активности в других геномных регионах / организмах. Поддерживая существующие участки гетерохроматина, RITS образует комплекс с миРНК дополнительный с локальными генами и стабильно связывает локальные метилированные гистоны, действуя котранскрипционно, разрушая любые возникающие транскрипты пре-мРНК, которые инициируются РНК-полимераза. Формирование такой гетерохроматиновой области, хотя и не ее поддержание, зависит от дайсера, предположительно потому, что дайсер необходим для генерации начального набора миРНК которые нацелены на последующие стенограммы.[58] Было высказано предположение, что поддержание гетерохроматина функционирует как самоусиливающаяся петля обратной связи, поскольку новые siRNAs формируются из случайных возникающих транскриптов с помощью RdRP для включения в локальные комплексы RITS.[59] Актуальность наблюдений из областей спаривающегося типа и центромер делящихся дрожжей для млекопитающие не ясно, поскольку поддержание гетерохроматина в клетках млекопитающих может быть независимым от компонентов пути РНКи.[60]
Перекрестные помехи с редактированием РНК
Тип Редактирование РНК что наиболее часто встречается у новообращенных высших эукариот аденозин нуклеотиды в инозин в дцРНК через фермент аденозиндезаминаза (АДАР).[61] Первоначально в 2000 г. было высказано предположение, что пути редактирования РНКи и A → I РНК могут конкурировать за общий субстрат дцРНК.[62] Некоторые пре-миРНК подвергаются редактированию РНК A → I[63][64] и этот механизм может регулировать процессинг и экспрессию зрелых miRNA.[64] Кроме того, по крайней мере один ADAR млекопитающего может изолировать миРНК из компонентов пути РНКи.[65] Дальнейшая поддержка этой модели исходит из исследований ADAR-null. C. elegans штаммы, указывающие на то, что редактирование РНК A → I может противодействовать РНКи сайленсингу эндогенных генов и трансгенов.[66]

Различия между организмами
Организмы различаются по своей способности захватывать чужеродную дцРНК и использовать ее в пути РНКи. Эффекты РНК-интерференции могут быть как системными, так и наследственными у растений и растений. C. elegans, хотя и не в Дрозофила или млекопитающие. Считается, что в растениях РНКи размножаются путем передачи миРНК между ячейками через плазмодесматы (каналы в клеточных стенках, обеспечивающие связь и транспорт).[38] Наследственность происходит от метилирование промоторов, нацеленных на РНКи; новый паттерн метилирования копируется в каждом новом поколении клетки.[68] Общее различие между растениями и животными заключается в нацеливании на эндогенно продуцируемые miRNAs; у растений miRNAs обычно полностью или почти идеально комплементарны своим генам-мишеням и вызывают прямое расщепление мРНК с помощью RISC, тогда как miRNA животных имеют тенденцию быть более дивергентными по последовательности и индуцируют репрессию трансляции.[67] Этот трансляционный эффект может быть вызван ингибированием взаимодействий трансляции. факторы инициирования с РНК-мессенджерами полиадениновый хвост.[69]
Некоторые эукариотические простейшие Такие как Leishmania major и Trypanosoma cruzi полностью лишены пути РНКи.[70][71] Большинство или все компоненты отсутствуют в некоторых грибы, в первую очередь модельный организм Saccharomyces cerevisiae.[72] Присутствие РНКи у других видов почкующихся дрожжей, таких как Saccharomyces castellii и грибковые микроорганизмы албиканс, дополнительно демонстрирует, что индукция двух родственных РНКи белков из S. castellii способствует РНКи в С. cerevisiae.[73] Этот определенный аскомицеты и базидиомицеты отсутствие путей РНК-интерференции указывает на то, что белки, необходимые для подавления РНК, были потеряны независимо от многих грибковых родословная, возможно, из-за эволюции нового пути с аналогичной функцией или из-за отсутствия избирательного преимущества в некоторых ниши.[74]
Связанные прокариотические системы
На экспрессию генов у прокариот влияет система на основе РНК, в некоторых отношениях сходная с РНКи. Здесь гены, кодирующие РНК, контролируют изобилие или трансляцию мРНК, производя комплементарную РНК, которая отжигается с мРНК. Однако эти регуляторные РНК обычно не считаются аналогами миРНК, потому что в этом не участвует фермент дайсера.[75] Было высказано предположение, что CRISPR вмешательство системы прокариот аналогичны системам интерференции РНК эукариот, хотя ни один из белковых компонентов не является ортологичный.[76]
Биологические функции
Этот раздел должен быть обновлено. (Май 2020 г.) |
Иммунитет
РНК-интерференция - жизненно важная часть иммунная реакция к вирусам и прочим чужеродным генетический материал, особенно в растениях, где он также может препятствовать самораспространению транспозонов.[77] Такие растения как Arabidopsis thaliana экспресс множественный dicer гомологи которые по-разному реагируют, когда растение подвергается воздействию различных вирусов.[78] Еще до того, как путь РНКи был полностью понят, было известно, что индуцированное молчание генов в растениях может распространяться по всему растению с системным эффектом и может передаваться из стада в отпрыск растения через прививка.[79] Этот феномен был с тех пор признан особенностью адаптивной иммунной системы растений и позволяет всему растению реагировать на вирус после первоначального локального столкновения.[80] В ответ многие вирусы растений развили сложные механизмы для подавления ответа РНКи.[81] К ним относятся вирусные белки, которые связывают короткие двухцепочечные фрагменты РНК с одноцепочечными выступающими концами, такие как те, которые продуцируются дайсером.[82] В геномах некоторых растений также экспрессируются эндогенные миРНК в ответ на заражение определенными типами бактерии.[83] Эти эффекты могут быть частью общей реакции на патогены, которая подавляет любой метаболический процесс в организме хозяина, который способствует процессу инфекции.[84]
Хотя животные обычно экспрессируют меньше вариантов фермента дайсера, чем растения, РНКи у некоторых животных вызывает противовирусный ответ. И у молодых, и у взрослых Дрозофила, РНК-интерференция важна в противовирусном врожденный иммунитет и активен против патогенов, таких как Вирус Drosophila X.[85][86] Аналогичная роль в иммунитете может действовать в C. elegans, поскольку белки аргонавта активируются в ответ на вирусы, а черви, избыточно экспрессирующие компоненты пути РНКи, устойчивы к вирусной инфекции.[87][88]
Роль РНК-интерференции в врожденном иммунитете млекопитающих плохо изучена, и имеется относительно мало данных. Однако существование вирусов, кодирующих гены, способные подавлять РНКи-ответ в клетках млекопитающих, может свидетельствовать в пользу РНКи-зависимого иммунного ответа млекопитающих.[89][90] хотя эта гипотеза была оспорена как плохо обоснованная.[91]Представлены доказательства существования функционального пути противовирусной РНКи в клетках млекопитающих.[92][93]
Существуют и другие функции РНКи в вирусах млекопитающих, такие как миРНК, экспрессируемые вирус герпеса что может действовать как гетерохроматин Организационные триггеры опосредуют вирусную латентность.[94]
Подавление генов
Эндогенно экспрессируемые миРНК, включая обе интронный и межгенный miRNAs, наиболее важны в репрессии трансляции[67] и в регулировании развития, особенно по срокам морфогенез и поддержание недифференцированный или неполностью дифференцированные типы клеток, такие как стволовые клетки.[95] Роль эндогенно экспрессируемой miRNA в подавлении экспрессия гена впервые был описан в C. elegans в 1993 г.[96] У растений эта функция была обнаружена, когда «микроРНК JAW» Арабидопсис было показано, что он участвует в регуляции нескольких генов, контролирующих форму растений.[97] У растений большинство генов, регулируемых миРНК, являются факторы транскрипции;[98] таким образом, активность микроРНК особенно обширна и регулирует все генные сети во время развития путем модуляции экспрессии ключевых регуляторных генов, включая факторы транскрипции, а также F-бокс белки.[99] У многих организмов, включая человека, миРНК связаны с образованием опухоли и нарушение регуляции клеточный цикл. Здесь miRNA могут функционировать как онкогены и опухолевые супрессоры.[100]
Эволюция
На основе на основе экономии филогенетический анализ, самый последний общий предок из всех эукариоты скорее всего, уже обладали ранним путем интерференции РНК; отсутствие этого пути у некоторых эукариот считается производной характеристикой.[101] Эта предковая система РНКи, вероятно, содержала по крайней мере один дайсер-подобный белок, один аргонавт, один Белок PIWI, и РНК-зависимая РНК-полимераза это могло также сыграть другие клеточные роли. Масштабный сравнительная геномика исследование также показывает, что эукариотические группа короны уже обладали этими компонентами, которые, возможно, затем имели более тесные функциональные ассоциации с общими системами деградации РНК, такими как экзосома.[102] Это исследование также предполагает, что семейство РНК-связывающих белков аргонавтов, общих для эукариот, большинства архей и, по крайней мере, некоторых бактерий (таких как Aquifex aeolicus ), гомологичен и первоначально развился из компонентов инициирование перевода система.
Обычно считается, что предковой функцией системы РНКи была иммунная защита от экзогенных генетических элементов, таких как транспозоны и вирусные геномы.[101][103] Родственные функции, такие как модификация гистонов, возможно, уже присутствовали у предков современных эукариот, хотя другие функции, такие как регуляция развития с помощью miRNA, как полагают, развились позже.[101]
Гены РНК-интерференции как компоненты противовирусной врожденной иммунной системы у многих эукариот участвуют в эволюционная гонка вооружений с вирусными генами. Некоторые вирусы развили механизмы для подавления ответа РНКи в своих клетках-хозяевах, особенно для вирусов растений.[81] Исследования темпов эволюции в Дрозофила показали, что гены пути РНКи подвержены сильному направленный выбор и являются одними из самых быстрыхразвивающийся гены в Дрозофила геном.[104]
Приложения
Джин нокдаун
Путь интерференции РНК часто используется в экспериментальная биология изучить функцию генов в культура клеток и in vivo в модельные организмы.[6] Двухцепочечная РНК синтезируется с последовательностью, комплементарной интересующему гену, и вводится в клетку или организм, где она распознается как экзогенный генетический материал и активирует путь РНКи. Используя этот механизм, исследователи могут вызвать резкое снижение экспрессии целевого гена. Изучение эффектов этого снижения может показать физиологическую роль продукта гена. Поскольку РНКи не может полностью исключить экспрессию гена, этот метод иногда называют "сбить ", чтобы отличить это от"нокаутировать «процедуры, в которых полностью исключается экспрессия гена.[105] В недавнем исследовании, подтверждение эффективности подавления РНКи с использованием данных массива генов, показало, что частота неудач составляет 18,5% в 429 независимых экспериментах.[106]
Обширные усилия в вычислительная биология были направлены на разработку успешных реагентов дцРНК, которые максимизируют нокдаун гена, но минимизируют «нецелевые» эффекты. Эффекты вне мишени возникают, когда введенная РНК имеет базовую последовательность, которая может спариваться и, таким образом, снижать экспрессию нескольких генов. Такие проблемы возникают чаще, когда дцРНК содержит повторяющиеся последовательности. Было подсчитано, изучая геномы людей, C. elegans и С. Помбе что около 10% возможных миРНК имеют существенные нецелевые эффекты.[10] Было разработано множество программных инструментов, реализующих алгоритмы для дизайна общего[107][108] специфично для млекопитающих,[109] и вирус-специфичный[110] миРНК которые автоматически проверяются на возможную перекрестную реактивность.
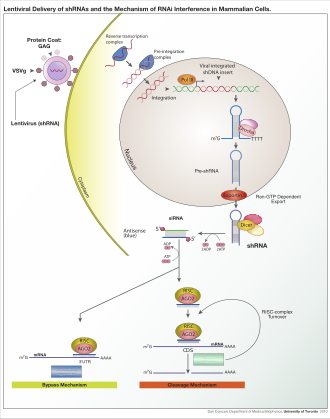
В зависимости от организма и экспериментальной системы, экзогенная РНК может быть длинной цепью, предназначенной для расщепления дицером, или короткой РНК, предназначенной для использования в качестве миРНК субстраты. В большинстве клеток млекопитающих используются более короткие РНК, потому что длинные двухцепочечные молекулы РНК вызывают у млекопитающих интерферон ответ, форма врожденный иммунитет который неспецифически реагирует на чужеродный генетический материал.[111] Мышь ооциты и клетки ранних мышей эмбрионы лишены этой реакции на экзогенную дцРНК и поэтому являются общей модельной системой для изучения эффектов нокдауна генов у млекопитающих.[112] Также были разработаны специализированные лабораторные методы для повышения полезности РНКи в системах млекопитающих, избегая прямого введения миРНК, например, стабильной трансфекция с плазмида кодирование соответствующей последовательности, из которой миРНК можно переписать,[113] или более сложным лентивирусный векторные системы, позволяющие индуцибельную активацию или дезактивацию транскрипции, известные как условная РНКи.[114][115]
Функциональная геномика

Наиболее функциональная геномика применения РНКи у животных использовали C. elegans[116] и Дрозофила,[117] поскольку это общие модельные организмы в которых РНКи наиболее эффективны. C. elegans особенно полезен для исследования РНКи по двум причинам: во-первых, эффекты молчания генов обычно наследуются, а во-вторых, потому что доставка дцРНК чрезвычайно проста. Через механизм, детали которого плохо изучены, бактерии, такие как Кишечная палочка которые несут желаемую дцРНК, могут быть скармлены червям и будут передавать полезную нагрузку своей РНК червю через кишечный тракт. Эта «доставка путем кормления» столь же эффективна для индукции сайленсинга генов, как и более дорогие и трудоемкие методы доставки, такие как вымачивание червей в растворе дцРНК и инъекция дцРНК в гонады.[118] Хотя доставка является более сложной для большинства других организмов, предпринимаются также усилия по проведению крупномасштабного геномного скрининга в культуре клеток с клетками млекопитающих.[119]
Подходы к созданию полногеномных библиотек РНКи могут потребовать большей сложности, чем создание единого миРНК для определенного набора экспериментальных условий. Искусственные нейронные сети часто используются для дизайна миРНК библиотеки[120] и предсказать их вероятную эффективность при нокдауне гена.[121] Массовый геномный скрининг широко рассматривается как многообещающий метод аннотация генома и послужило толчком к разработке высокопроизводительных методов скрининга, основанных на микрочипы.[122][123] Однако полезность этих экранов и способность методов, разработанных на модельных организмах, распространяться даже на близкородственные виды, были поставлены под сомнение, например, из C. elegans родственным паразитическим нематодам.[124][125]
Функциональная геномика с использованием РНКи является особенно привлекательной техникой для геномного картирования и аннотации растений, поскольку многие растения являются полиплоид, что создает серьезные проблемы для более традиционных методов генной инженерии. Например, РНКи успешно использовалась для исследований функциональной геномики в хлеб пшеничный (который гексаплоид)[126] а также более распространенные системы моделей завода Арабидопсис и кукуруза.[127]
Лекарство
История использования РНКи в медицине

Первый экземпляр РНК молчание у животных было задокументировано в 1996 году, когда Гуо и Кемфуэс заметили это, введя смысл и антисмысловая РНК к мРНК par-1 в Caenorhabditis elegans вызвали ухудшение качества сообщения par-1.[128] Считалось, что эта деградация запускается одноцепочечной РНК (оцРНК), но два года спустя, в 1998 году, Файер и Мелло обнаружили, что эта способность подавлять экспрессию гена par-1 фактически запускается двухцепочечной РНК (дцРНК). ).[128] В конечном итоге они поделятся Нобелевская премия по физиологии и медицине за это открытие.[129] Сразу после новаторского открытия Файра и Мелло Эльбашир и др. обнаружен, используя синтетически сделанный малая интерферирующая РНК (siRNA) можно было нацелить на подавление молчания определенных последовательностей в гене, а не на подавление всего гена.[130] Только год спустя McCaffrey и его коллеги продемонстрировали, что это специфичное для последовательности молчание имеет терапевтическое применение, нацеленное на последовательность из Гепатит С вирус в трансгенный мышей.[131] С тех пор несколько исследователей пытались расширить терапевтическое применение РНКи, уделяя особое внимание генам-мишеням, вызывающим различные типы рак.[132][133] К 2006 г. первые приложения, достигшие клинические испытания находились на лечении дегенерация желтого пятна и респираторно-синцитиальный вирус.[134] Четыре года спустя было начато первое клиническое испытание фазы I на людях с использованием система доставки наночастиц к цели солидные опухоли.[135] Хотя в настоящее время в большинстве исследований изучается применение РНКи в лечении рака, список возможных применений обширен. РНКи потенциально могут быть использованы для лечения вирусы,[136] бактериальные болезни,[137] паразиты,[138] неадаптивный генетический мутации,[139] контролировать потребление наркотиков,[140] облегчить боль,[141] и даже модулировать спать.[142]
Терапевтические приложения
Вирусная инфекция
Противовирусное лечение - одно из первых предложенных медицинских приложений на основе РНКи, и были разработаны два разных типа. Первый тип направлен на вирусные РНК. Многие исследования показали, что нацеливание на вирусные РНК может подавлять репликацию множества вирусов, включая ВИЧ,[143] ВПЧ,[144] гепатит А,[145] гепатит Б,[146] вирус гриппа,[147][148][149][150] респираторно-синцитиальный вирус (RSV),[150] Коронавирус SARS (SARS-CoV),[150] аденовирус[150] и вирус кори.[151] Другая стратегия состоит в том, чтобы заблокировать начальные проникновения вируса путем нацеливания на гены клетки-хозяина.[152] Например, подавление хемокиновых рецепторов (CXCR4 и CCR5 ) на клетках-хозяевах может предотвратить проникновение вируса ВИЧ.[153]
Рак
Хотя традиционные химиотерапия может эффективно убивать раковые клетки, отсутствие специфичности для различения нормальных клеток и раковых клеток в этих методах лечения обычно вызывает серьезные побочные эффекты. Многочисленные исследования продемонстрировали, что РНКи могут обеспечить более специфический подход к подавлению роста опухоли путем воздействия на гены, связанные с раком (т. Е. онкоген ).[154] Также было высказано предположение, что РНКи могут повышать чувствительность раковых клеток к химиотерапевтические агенты, обеспечивающий комбинаторный терапевтический подход с химиотерапией.[155] Еще одно возможное лечение на основе РНКи - ингибирование клеточной инвазии и миграция.[156]
Неврологические заболевания
Стратегии РНКи также показывают потенциал для лечения нейродегенеративные заболевания. Исследования на клетках и мышах показали, что специфическое нацеливание Бета-амилоид -продуцирующие гены (например, BACE1 и APP) с помощью РНКи могут значительно снизить количество пептида Aβ, которое коррелирует с причиной Болезнь Альцгеймера.[157][158][159] Кроме того, эти подходы, основанные на подавлении звука, также обеспечивают многообещающие результаты при лечении болезнь Паркинсона и Полиглутаминовая болезнь.[160][161][162]
Трудности терапевтического применения
Чтобы реализовать клинический потенциал РНКи, миРНК необходимо эффективно транспортировать к клеткам тканей-мишеней. Однако существуют различные барьеры, которые необходимо устранить, прежде чем его можно будет использовать в клинических условиях. Например, «голый» миРНК подвержен ряду препятствий, которые снижают его терапевтическую эффективность.[163] Кроме того, как только миРНК попадает в кровоток, голая РНК может расщепляться нуклеазами сыворотки крови и стимулировать врожденную иммунную систему.[163] Из-за своего размера и сильно полианионной (содержащей отрицательные заряды на нескольких сайтах) природы немодифицированные молекулы миРНК не могут легко проникать в клетки через клеточную мембрану. Следовательно, искусственный или миРНК, инкапсулированная наночастицами должны быть использованы. Однако транспортировка siRNA через клеточную мембрану по-прежнему имеет свои уникальные проблемы. Если миРНК переносится через клеточную мембрану, может возникнуть непреднамеренная токсичность, если терапевтические дозы не оптимизированы, и миРНК могут проявлять нецелевые эффекты (например, непреднамеренное подавление активности генов с частичная комплементарность последовательностей ).[164] Даже после попадания в клетки требуется повторное дозирование, поскольку их эффекты ослабляются при каждом делении клеток. Как описано ранее, части вектора, транспортирующие дцРНК, также могут иметь регуляторные эффекты. Поэтому следует учитывать и контролировать неспецифические побочные эффекты.[165]
Лечение рака
По сравнению с химиотерапией или другими противораковыми препаратами, препарат siRNA имеет много преимуществ.[166] SiRNA действует на посттранскрипционной стадии экспрессии генов, поэтому она не модифицирует и не изменяет ДНК, оказывая вредное воздействие.[166] SiRNA также можно использовать для получения определенного ответа определенным способом, например, путем снижения уровня подавления экспрессии генов.[166] В одной раковой клетке siRNA может вызывать резкое подавление экспрессии генов всего с несколькими копиями.[166] Это происходит путем подавления генов, способствующих развитию рака, с помощью РНКи, а также нацеливания на последовательность мРНК.[166]
РНКи-препараты лечат рак, подавляя определенные гены, способствующие развитию рака.[166] Это достигается путем дополнения раковых генов РНКи, например, сохранения последовательностей мРНК в соответствии с препаратом РНКи.[166] В идеале РНКи следует вводить и / или химически модифицировать, чтобы РНКи могли более эффективно достигать раковых клеток.[166] Поглощение и регуляция РНКи контролируется почками.[166]
Стимуляция иммунного ответа
Иммунная система человека делится на две отдельные ветви: врожденная иммунная система и адаптивная иммунная система.[167] Врожденная иммунная система является первой защитой от инфекции и реагирует на патогены обычным образом.[167] С другой стороны, адаптивная иммунная система, система, которая возникла позже, чем врожденная, состоит в основном из высокоспециализированных В- и Т-клеток, которые обучены реагировать на определенные части патогенных молекул.[167]
Проблема между старыми и новыми патогенами помогла создать систему защищенных клеток и частиц, которую называют безопасным каркасом.[167] Эта структура дала людям армейские системы, которые ищут и уничтожают частицы-захватчики, такие как патогены, микроскопические организмы, паразиты и инфекции.[167] Безопасная структура для млекопитающих была разработана для включения миРНК в качестве инструмента для индикации вирусного заражения, что позволило миРНК создать интенсивный врожденный иммунный ответ.[167]
siRNA контролируется врожденной иммунной системой, которую можно разделить на острые воспалительные реакции и противовирусные реакции.[167] Воспалительный ответ создается с помощью сигналов от небольших сигнальных молекул или цитокинов.[167] К ним относятся интерлейкин-1 (IL-1), интерлейкин-6 (IL-6), интерлейкин-12 (IL-12) и фактор некроза опухоли α (TNF-α).[167] Врожденная иммунная система генерирует воспаление и противовирусные реакции, которые вызывают рецепторы распознавания паттернов высвобождения (PRR).[167] Эти рецепторы помогают определить, какие патогены являются вирусами, грибами или бактериями.[167] Более того, важность siRNA и врожденной иммунной системы состоит в том, чтобы включать больше PRR, чтобы помочь распознавать различные структуры РНК.[167] Это увеличивает вероятность того, что миРНК вызовет иммуностимулирующий ответ в случае воздействия патогена.[167]
Перспективы как лечебная методика
Этот раздел должен быть обновлено. (Май 2020 г.) |
Клинические исследования фаз I и II терапии миРНК, проведенные в период с 2015 по 2017 год, продемонстрировали эффективные и надежные методы лечения. нокдаун генов в печень, с некоторыми признаками клинического улучшения и без неприемлемой токсичности.[164] В настоящее время проводятся два исследования фазы III по лечению семейных нейродегенеративных и сердечных синдромов, вызванных мутациями в транстиретин (TTR).[164] Многочисленные публикации показали, что системы доставки in vivo очень перспективны и разнообразны по характеристикам, что позволяет использовать их в самых разных областях. Система доставки наночастиц является наиболее многообещающей, но этот метод создает дополнительные проблемы в увеличить масштаб производственного процесса, например, необходимость в строго контролируемых процессах смешивания для достижения стабильного качества лекарственного препарата.[163]
В таблице ниже показаны различные препараты, использующие РНК-интерференцию, а также их фазы и статус в клинических испытаниях по состоянию на 2013 год.[163]
| Препарат, средство, медикамент | Цель | Система доставки | Болезнь | Фаза | Положение дел | Компания | Идентификатор |
| ALN – VSP02 | KSP и VEGF | LNP | Солидные опухоли | я | Завершенный | Alnylam Pharmaceuticals | NCT01158079 |
| миРНК-EphA2-DOPC | EphA2 | LNP | Продвинутые формы рака | я | Рекрутинг | Онкологический центр доктора медицины Андерсона | NCT01591356 |
| Atu027 | ПКН3 | LNP | Солидные опухоли | я | Завершенный | Безмолвная терапия | NCT00938574 |
| ТКМ – 080301 | PLK1 | LNP | Рак | я | Рекрутинг | Tekmira Pharmaceutical | NCT01262235 |
| ТКМ – 100201 | VP24, VP35, L-полимераза вируса Эбола Заира | LNP | Эбола-вирусная инфекция | я | Рекрутинг | Tekmira Pharmaceutical | NCT01518881 |
| ALN – RSV01 | Нуклеокапсид RSV | Голая миРНК | Респираторно-синцитиальные вирусные инфекции | II | Завершенный | Alnylam Pharmaceuticals | NCT00658086 |
| PRO-040201 | ApoB | LNP | Гиперхолестеринемия | я | Прекращено | Tekmira Pharmaceutical | NCT00927459 |
| ALN – PCS02 | PCSK9 | LNP | Гиперхолестеринемия | я | Завершенный | Alnylam Pharmaceuticals | NCT01437059 |
| ALN – TTR02 | TTR | LNP | Транстиретин-опосредованный амилоидоз | II | Рекрутинг | Alnylam Pharmaceuticals | NCT01617967 |
| КАЛАА-01 | RRM2 | Циклодекстрин НП | Солидные опухоли | я | Активный | Calando Pharmaceuticals | NCT00689065 |
| TD101 | K6a (мутация N171K) | Голая миРНК | Врожденная пахионихия | я | Завершенный | Pachyonychia Congenita Project | NCT00716014 |
| AGN211745 | VEGFR1 | Голая миРНК | Возрастная дегенерация желтого пятна, неоваскуляризация хориоидеи | II | Прекращено | Аллерган | NCT00395057 |
| QPI-1007 | CASP2 | Голая миРНК | Атрофия зрительного нерва, неартеритическая передняя ишемическая оптическая нейропатия | я | Завершенный | Quark Pharmaceuticals | NCT01064505 |
| I5NP | p53 | Голая миРНК | Поражение почек, острая почечная недостаточность | я | Завершенный | Quark Pharmaceuticals | NCT00554359 |
| Задержка функции трансплантата, осложнения трансплантации почки | I, II | Рекрутинг | Quark Pharmaceuticals | NCT00802347 | |||
| ПФ-655 (ПФ-04523655) | RTP801 (собственная цель) | Голая миРНК | Неоваскуляризация хориоидеи, диабетическая ретинопатия, диабетический макулярный отек | II | Активный | Quark Pharmaceuticals | NCT01445899 |
| siG12D LODER | KRAS | LODER полимер | Панкреатический рак | II | Рекрутинг | Silenseed | NCT01676259 |
| Бевасираниб | VEGF | Голая миРНК | Диабетический отек желтого пятна, дегенерация желтого пятна | II | Завершенный | Опко Здоровье | NCT00306904 |
| SYL1001 | TRPV1 | Голая миРНК | Глазная боль, синдром сухого глаза | I, II | Рекрутинг | Sylentis | NCT01776658 |
| SYL040012 | ADRB2 | Голая миРНК | Глазная гипертензия, открытоугольная глаукома | II | Рекрутинг | Sylentis | NCT01739244 |
| CEQ508 | CTNNB1 | КшРНК, несущая Escherichia coli | Семейный аденоматозный полипоз | I, II | Рекрутинг | Марина Биотех | Неизвестный |
| RXi-109 | CTGF | Самодоставляющееся соединение РНКи | Профилактика рубцовых рубцов | я | Рекрутинг | RXi Pharmaceuticals | NCT01780077 |
| ALN – TTRsc | TTR | конъюгат миРНК-GalNAc | Транстиретин-опосредованный амилоидоз | я | Рекрутинг | Alnylam Pharmaceuticals | NCT01814839 |
| ARC-520 | Консервативные области HBV | ЦОД | HBV | я | Рекрутинг | Arrowhead Research | NCT01872065 |
Биотехнологии
РНК-интерференция использовалась для приложений в биотехнология и приближается к коммерциализации в других областях. RNAi привела к изобретению новых сельскохозяйственных культур, таких как безникотиновый табак, кофе без кофеина, растения, обогащенные питательными веществами, и гипоаллергенные культуры. Генно-инженерный Арктические яблоки получил одобрение FDA в 2015 году.[168] Яблоки были произведены путем подавления РНКи гена PPO (полифенолоксидазы), в результате чего были получены сорта яблок, которые не потемнели после нарезки. Яблоки с глухим PPO не могут превращать хлорогеновую кислоту в стандартный хиноновый продукт.[2]
Есть несколько возможностей применения РНКи в растениеводстве для ее улучшения, таких как устойчивость к стрессу и повышение уровня питательности. RNAi докажет свой потенциал ингибирования фотодыхания для повышения продуктивности растений C3. Эта технология «нокдауна» может быть полезна для стимулирования раннего цветения, отсроченного созревания, замедленного старения, нарушения покоя, снятия стресса растений, преодоления самопроизвольной стерильности и т. Д.[2]
Еда
РНКи использовались для генетической инженерии растений для производства более низких уровней естественных токсинов растений. Такие методы используют преимущества стабильного и наследуемого фенотипа РНКи в растительных стадах. Хлопок семена богаты диетический белок но естественно содержат токсичные терпеноид товар госсипол, что делает их непригодными для употребления в пищу. RNAi использовался для производства запасов хлопка, семена которых содержат пониженные уровни дельта-кадиненсинтаза, ключевой фермент в производстве госсипола, не влияющий на производство фермента в других частях растения, где госсипол сам по себе важен для предотвращения повреждения от вредителей растений.[169] Аналогичные усилия были направлены на сокращение цианогенный натуральный продукт линамарин в маниока растения.[170]
Нет растительных продуктов, в которых используется РНКи генная инженерия еще не вышли из экспериментальной стадии. Усилия разработчиков успешно снизили уровни аллергены в помидор растения[171] и обогащение растений, таких как помидоры, диетическими антиоксиданты.[172] Предыдущие коммерческие продукты, включая Флавр Савр помидор и два сорта из кольцо -устойчивый папайя, изначально были разработаны с использованием антисмысловой технологии, но, вероятно, использовали путь RNAi.[173][174] РНКи подавление альфа-амилаза также использовались для уменьшения Aspergillus flavus грибковый рост кукурузы, который в противном случае загрязнил бы зерна опасными афлатоксины.[175] Подавление синтазы слезоточивого фактора в лук произвели лук без слез, а РНКи использовались в генах BP1 в рапс для улучшения фотосинтеза.[176] Гены SBEIIa и SBEIIb в пшенице были нацелены на пшеницу для получения более высоких уровней амилоза с целью улучшения работы кишечника.[177]
Другие культуры
Еще одна попытка уменьшила количество предшественников вероятного канцерогены в табак растения.[178] Другие свойства растений, которые были разработаны в лаборатории, включают производство ненаркотик натуральные продукты опийный мак[179] и устойчивость к распространенным вирусам растений.[180]
Инсектицид
РНКи разрабатывается как инсектицид, используя несколько подходов, включая генную инженерию и актуальное применение.[4] Клетки средней кишки некоторых насекомых захватывают молекулы дцРНК в процессе, называемом РНКи окружающей среды.[181] У некоторых насекомых эффект носит системный характер, поскольку сигнал распространяется по всему телу насекомого (это называется системной РНКи).[182]
Животные, подвергшиеся воздействию РНКи в дозах, в миллионы раз превышающих ожидаемые уровни воздействия на человека, не проявляют никаких побочных эффектов.[183]
РНКи по-разному влияет на разные виды Чешуекрылые (бабочки и мотыльки).[184] Возможно, потому что их слюна кишечный сок лучше расщепляет РНК, хлопковая совка, то совка совка и Азиатский рисовый мотылек до сих пор не доказана их чувствительность к РНКи при кормлении.[4]
Недавние данные свидетельствуют о том, что устойчивость к РНКи может иметь широкий спектр, а это означает, что устойчивость к одной последовательности может придавать устойчивость к другим последовательностям дцРНК. В одной лабораторной популяции западного кукурузного корневого червя устойчивость возникла из-за недостаточного поглощения дцРНК DvSnf7 через кишечник.[185] Когда другие последовательности дцРНК были протестированы против DvSnf7, другие последовательности перестали быть эффективными, что предполагает, что управление устойчивостью будет более трудным, чем простое отключение последовательностей дцРНК. Сочетание нескольких стратегий, таких как создание белка Cry, полученного из бактерии, называемой Bacillus thuringiensis (Bt), и РНКи в одном растении задерживают начало устойчивости.[4][186]
Трансгенные растения
Трансгенные культуры были сделаны для экспрессии дцРНК, тщательно подобранной, чтобы заставить замолчать важные гены у вредителей-мишеней. Эти дцРНК предназначены для воздействия только на насекомых, которые экспрессируют определенные генные последовательности. Как Доказательство принципа, в 2009 году исследование показало, что РНК могут убить любой из четырех видов плодовых мух, не нанося вреда трем другим.[4]
В 2012 Сингента купил бельгийскую фирму RNAi Devgen за 522 миллиона долларов и Monsanto заплатил 29,2 миллиона долларов за исключительные права на интеллектуальная собственность из Alnylam Pharmaceuticals. В Международный центр картофеля в Лима, Перу ищет гены, нацеленные на сладкий картофельный долгоносик, жук, чьи личинки уничтожают сладкий картофель глобально. Другие исследователи пытаются заставить замолчать гены муравьев, гусениц и пыльцевых жуков. Monsanto, вероятно, будет первой на рынке с трансгенными семенами кукурузы, которые экспрессируют дцРНК на основе гена Snf7 из западный кукурузный корень, а жук чей личинки только в Соединенных Штатах ежегодно наносят ущерб в размере одного миллиарда долларов. В статье 2012 года было показано, что подавление Snf7 останавливает рост личинок, убивая их в считанные дни. В 2013 году та же команда показала, что РНК влияет на очень немногие другие виды.[4]
Актуальные
Альтернативно дцРНК может поставляться без генной инженерии. Один из подходов - добавить их в орошение воды. Молекулы поглощаются растениями. сосудистый системы и ядовитых насекомых, питающихся ими. Другой подход включает распыление дцРНК, как обычный пестицид. Это позволит быстрее адаптироваться к сопротивлению. Такие подходы потребуют недорогих источников дцРНК, которые в настоящее время не существуют.[4]
Скрининг в масштабе генома
Исследования РНКи в масштабе генома опираются на высокопроизводительный скрининг (HTS) технология. Технология RNAi HTS позволяет проводить скрининг потери функции по всему геному и широко используется для идентификации генов, связанных с конкретными фенотипами. Эта технология была провозглашена потенциальной второй волной геномики после первой волны геномики экспрессия гена микрочип и однонуклеотидный полиморфизм платформы открытия.[187]Одним из основных преимуществ скрининга РНКи на уровне генома является его способность одновременно опрашивать тысячи генов. Благодаря способности генерировать большой объем данных за эксперимент, скрининг РНКи в масштабе генома привел к взрывному росту скорости генерации данных. Использование таких больших наборов данных является фундаментальной проблемой, требующей подходящих методов статистики / биоинформатики. Основной процесс скрининга РНКи на основе клеток включает выбор библиотеки РНКи, надежных и стабильных типов клеток, трансфекцию агентами РНКи, обработку / инкубацию, обнаружение сигналов, анализ и идентификацию важных генов или терапевтических мишеней.[188]
История

Процесс РНКи упоминался как «совместное подавление» и «подавление», когда наблюдался до того, как был известен механизм, связанный с РНК. Открытию РНКи сначала предшествовали наблюдения за ингибированием транскрипции антисмысловой РНК экспрессируется в трансгенный растения[190] и, что более конкретно, сообщениями о неожиданных результатах экспериментов, проведенных учеными-растениеводами Соединенные Штаты и Нидерланды в начале 1990-х гг.[191] В попытке изменить цветок цвета в петунии, исследователи ввели дополнительные копии гена, кодирующего халкон-синтаза, ключевой фермент для цветов пигментация в растения петунии с обычно розовыми или фиолетовыми цветками. Ожидалось, что сверхэкспрессия гена приведет к появлению более темных цветов, но вместо этого у некоторых цветов будет менее заметный пурпурный пигмент, иногда с пестрым рисунком, что указывает на то, что активность халконсинтазы была существенно снижена или стала подавляться в зависимости от контекста. Позже это будет объяснено как результат встраивания трансгена рядом с промоторами в противоположном направлении в различных положениях по всему геному некоторых трансформантов, что приводит к экспрессии антисмысловых транскриптов и подавлению гена, когда эти промоторы активны.Еще одно раннее наблюдение РНКи было получено в результате исследования грибок Neurospora crassa,[192] хотя это не было сразу признано родственным. Дальнейшее исследование феномена у растений показало, что подавление происходило из-за посттранскрипционного ингибирования экспрессии генов за счет увеличения скорости деградации мРНК.[193] Это явление было названо совместное подавление экспрессии генов, но молекулярный механизм остался неизвестным.[194]
Вскоре после этого посадили вирусологи работая над повышением устойчивости растений к вирусным заболеваниям, наблюдали подобное неожиданное явление. Хотя было известно, что растения, экспрессирующие вирусспецифические белки, демонстрируют повышенную толерантность или устойчивость к вирусной инфекции, не ожидалось, что растения, несущие только короткие, некодирующие области последовательностей вирусной РНК, будут демонстрировать аналогичные уровни защиты. Исследователи полагали, что вирусная РНК, продуцируемая трансгенами, также может подавлять репликацию вируса.[195] Обратный эксперимент, в котором короткие последовательности генов растений были введены в вирусы, показал, что целевой ген подавлялся в инфицированном растении.[196] Это явление было названо «сайленсингом генов, индуцированным вирусом» (VIGS), а совокупность таких явлений получила общее название посттранскрипционное молчание генов.[197]
После этих первоначальных наблюдений на растениях лаборатории начали искать это явление у других организмов.[198][199] Крейг С. Мелло и Эндрю Файер 1998 год Природа В статье сообщается о мощном эффекте подавления гена после инъекции двухцепочечной РНК в C. elegans.[200] При исследовании регуляции выработки мышечного белка они обнаружили, что ни мРНК, ни антисмысловая РНК инъекции оказали влияние на продукцию белка, но двухцепочечная РНК успешно подавила ген-мишень. В результате этой работы они создали термин РНКи. Это открытие представляло собой первое определение возбудителя этого явления. Огонь и Мелло были награждены премией 2006 года. Нобелевская премия по физиологии и медицине.[6]
Смотрите также
Рекомендации
- ^ видео природы, видео природы (16 декабря 2011 г.). "РНК-интерференция (РНКи): от Nature Video". YouTube. Получено 21 марта 2020.
- ^ а б c Саураб С., Видьярти А.С., Прасад Д. (март 2014 г.). «РНК-интерференция: от концепции к реальности в улучшении урожая». Planta. 239 (3): 543–64. Дои:10.1007 / s00425-013-2019-5. PMID 24402564.
- ^ Вайс Б., Давидкова Г., Чжоу Л.В. (март 1999 г.). «Генная терапия антисмысловой РНК для изучения и модуляции биологических процессов». Клеточные и молекулярные науки о жизни. 55 (3): 334–58. Дои:10.1007 / с000180050296. PMID 10228554. S2CID 9448271.
- ^ а б c d е ж грамм Купфершмидт, К. (2013). «Смертельная доза РНК». Наука. 341 (6147): 732–3. Bibcode:2013Наука ... 341..732K. Дои:10.1126 / science.341.6147.732. PMID 23950525.
- ^ Макрэ И.Дж., Чжоу К., Ли Ф., Репик А., Брукс А.Н., Канде В.З., Адамс П.Д., Дудна Д.А. (январь 2006 г.). «Структурная основа процессинга двухцепочечной РНК с помощью Dicer». Наука. 311 (5758): 195–8. Bibcode:2006Научный ... 311..195М. Дои:10.1126 / science.1121638. PMID 16410517. S2CID 23785494.
- ^ а б c d Данехольт Б. «Дополнительная информация: вмешательство РНК». Нобелевская премия по физиологии и медицине 2006 г.. Архивировано из оригинал 20 января 2007 г.. Получено 25 января 2007.
- ^ Багасра О., Приллиман К.Р. (август 2004 г.). «РНК-интерференция: молекулярная иммунная система». Журнал молекулярной гистологии. 35 (6): 545–53. CiteSeerX 10.1.1.456.1701. Дои:10.1007 / s10735-004-2192-8. PMID 15614608. S2CID 2966105.
- ^ Бернштейн Э., Кауди А.А., Хаммонд С.М., Хэннон Г.Дж. (январь 2001 г.). «Роль бидентатной рибонуклеазы на этапе инициации РНК-интерференции». Природа. 409 (6818): 363–6. Bibcode:2001Натура.409..363Б. Дои:10.1038/35053110. PMID 11201747. S2CID 4371481.

- ^ Сиоми Х., Сиоми МС (январь 2009 г.). «На пути к чтению кода РНК-интерференции». Природа. 457 (7228): 396–404. Bibcode:2009Натура.457..396S. Дои:10.1038 / природа07754. PMID 19158785. S2CID 205215974.
Заморе П.Д., Тушл Т., Шарп П.А., Бартель Д.П. (март 2000 г.). «РНКи: двухцепочечная РНК направляет АТФ-зависимое расщепление мРНК с интервалом от 21 до 23 нуклеотидов». Клетка. 101 (1): 25–33. Дои:10.1016 / S0092-8674 (00) 80620-0. PMID 10778853.
Вермёлен А., Белен Л., Рейнольдс А., Вольфсон А., Маршалл В.С., Карпилов Дж., Хворова А. (май 2005 г.). «Вклад структуры дцРНК в специфичность и эффективность Dicer». РНК. 11 (5): 674–82. Дои:10.1261 / rna.7272305. ЧВК 1370754. PMID 15811921.
Кастанотто Д., Росси Дж. Дж. (Январь 2009 г.). «Обещания и подводные камни терапии, основанной на РНК-интерференции». Природа. 457 (7228): 426–33. Bibcode:2009Натура 457..426C. Дои:10.1038 / природа07758. ЧВК 2702667. PMID 19158789. - ^ а б Цю С., Адема С.М., Лейн Т (2005). «Вычислительное исследование нецелевых эффектов РНК-интерференции». Исследования нуклеиновых кислот. 33 (6): 1834–47. Дои:10.1093 / нар / gki324. ЧВК 1072799. PMID 15800213.
- ^ а б Наканиши К. (сентябрь 2016 г.). «Анатомия RISC: как малые РНК и шапероны активируют белки Argonaute?». Междисциплинарные обзоры Wiley: РНК. 7 (5): 637–60. Дои:10.1002 / wrna.1356. ЧВК 5084781. PMID 27184117.
- ^ Лян Ц., Ван И, Мурота И, Лю Х, Смит Д., Сиоми М.С., Лю Ц. (сентябрь 2015 г.). «TAF11 собирает загрузочный комплекс RISC для повышения эффективности РНКи». Молекулярная клетка. 59 (5): 807–18. Дои:10.1016 / j.molcel.2015.07.006. ЧВК 4560963. PMID 26257286.
- ^ Алквист П. (2002). «РНК-зависимые РНК-полимеразы, вирусы и подавление РНК». Наука. 296 (5571): 1270–3. Bibcode:2002Наука ... 296.1270A. Дои:10.1126 / science.1069132. PMID 12016304. S2CID 42526536.
- ^ Робертс, TC (2015). «Механизм микроРНК». МикроРНК: фундаментальные науки. Достижения экспериментальной медицины и биологии. 887. С. 15–30. Дои:10.1007/978-3-319-22380-3_2. ISBN 978-3-319-22379-7. PMID 26662984.
- ^ а б c d Паркер Г., Эккерт Д., Бас Б. (2006). «RDE-4 предпочтительно связывает длинную дцРНК, и ее димеризация необходима для расщепления дцРНК до миРНК». РНК. 12 (5): 807–18. Дои:10.1261 / rna.2338706. ЧВК 1440910. PMID 16603715.
- ^ а б Баулкомб, округ Колумбия (январь 2007 г.). «Молекулярная биология. Усиленное молчание». Наука. 315 (5809): 199–200. Дои:10.1126 / science.1138030. PMID 17218517. S2CID 46285020.
- ^ а б Пак Дж., Огонь A (январь 2007 г.). «Различные популяции первичных и вторичных эффекторов во время РНКи у C. elegans». Наука. 315 (5809): 241–4. Bibcode:2007Научный ... 315..241П. Дои:10.1126 / наука.1132839. PMID 17124291. S2CID 46620298.
- ^ а б Зайен Т., Штайнер Ф.А., Тийссен К.Л., Plasterk RH (январь 2007 г.). «Вторичные миРНК возникают в результате синтеза непраймированной РНК и образуют отдельный класс». Наука. 315 (5809): 244–7. Bibcode:2007Научный ... 315..244С. Дои:10.1126 / science.1136699. PMID 17158288. S2CID 9483460.
- ^ Ван QL, Ли Чж (май 2007 г.). «Функции микроРНК в растениях». Границы биологических наук. 12: 3975–82. Дои:10.2741/2364. ЧВК 2851543. PMID 17485351. S2CID 23014413.
Чжао Ю., Шривастава Д. (апрель 2007 г.). «Взгляд на развитие функции микроРНК». Тенденции в биохимических науках. 32 (4): 189–97. Дои:10.1016 / j.tibs.2007.02.006. PMID 17350266. - ^ Грегори Р.И., Чендримада Т.П., Шихаттар Р. (2006). «Биогенез микроРНК: выделение и характеристика микропроцессорного комплекса». Протоколы микроРНК. Методы молекулярной биологии. 342. С. 33–47. Дои:10.1385/1-59745-123-1:33. ISBN 978-1-59745-123-9. PMID 16957365.
- ^ Пфеффер С., Заволан М., Грэссер Ф.А., Чиен М., Руссо Дж. Дж., Джу Дж., Джон Б., Энрайт А. Дж., Маркс Д., Сандер С., Тушл Т. (апрель 2004 г.). «Идентификация микроРНК, кодируемых вирусами». Наука. 304 (5671): 734–6. Bibcode:2004Наука ... 304..734P. Дои:10.1126 / science.1096781. PMID 15118162. S2CID 25287167.
- ^ Куреши А., Такур Н., Монга И., Такур А., Кумар М. (1 января 2014 г.). «VIRmiRNA: исчерпывающий ресурс для экспериментально подтвержденных вирусных miRNA и их мишеней». База данных. 2014: bau103. Дои:10.1093 / база данных / bau103. ЧВК 4224276. PMID 25380780.
- ^ Пиллаи Р.С., Бхаттачарья С.Н., Филипович В. (2007). «Подавление синтеза белка миРНК: сколько механизмов?». Тенденции Cell Biol. 17 (3): 118–26. Дои:10.1016 / j.tcb.2006.12.007. PMID 17197185.
- ^ Окамура К., Исидзука А., Сиоми Х., Сиоми М. (2004). «Различная роль белков Argonaute в путях расщепления малой РНК-направленной РНК». Genes Dev. 18 (14): 1655–66. Дои:10.1101 / gad.1210204. ЧВК 478188. PMID 15231716.
- ^ Ли Й, Накахара К., Фам Дж, Ким К., Хе З, Сонтхаймер Э, Картью Р. (2004). «Различная роль Dicer-1 и Dicer-2 Drosophila в путях подавления siRNA / miRNA». Клетка. 117 (1): 69–81. Дои:10.1016 / S0092-8674 (04) 00261-2. PMID 15066283.
- ^ miRBase.org
- ^ а б Фридман Р.К., Фарх К.К., Бурдж CB, Бартель Д.П. (2009). «Большинство мРНК млекопитающих являются консервативными мишенями для микроРНК». Genome Res. 19 (1): 92–105. Дои:10.1101 / гр.082701.108. ЧВК 2612969. PMID 18955434.
- ^ Лим LP, Лау NC, Гарретт-Энгеле П., Гримсон А., Шелтер Дж. М., Касл Дж., Бартель Д. П., Линсли П. С., Джонсон Дж. М. (февраль 2005 г.). «Анализ микроматрицы показывает, что некоторые микроРНК подавляют большое количество целевых мРНК». Природа. 433 (7027): 769–73. Bibcode:2005Натура.433..769L. Дои:10.1038 / природа03315. PMID 15685193. S2CID 4430576.
- ^ Зельбах М., Шванхойссер Б., Тирфельдер Н., Фанг З., Ханин Р., Раевский Н. (сентябрь 2008 г.). «Широко распространенные изменения в синтезе белков, вызванные микроРНК». Природа. 455 (7209): 58–63. Bibcode:2008Натура.455 ... 58С. Дои:10.1038 / природа07228. PMID 18668040. S2CID 4429008.
- ^ Бэк Д., Виллен Дж., Шин С., Камарго Ф. Д., Гиги С. П., Бартель Д. П. (сентябрь 2008 г.). «Влияние микроРНК на выход белка». Природа. 455 (7209): 64–71. Bibcode:2008Натура.455 ... 64Б. Дои:10.1038 / природа07242. ЧВК 2745094. PMID 18668037.
- ^ Palmero EI, de Campos SG, Campos M, de Souza NC, Guerreiro ID, Carvalho AL, Marques MM (июль 2011 г.). «Механизмы и роль нарушения регуляции микроРНК в возникновении и прогрессировании рака». Генетика и молекулярная биология. 34 (3): 363–70. Дои:10.1590 / S1415-47572011000300001. ЧВК 3168173. PMID 21931505.
- ^ Бернштейн C, Бернштейн H (май 2015 г.). «Эпигенетическое снижение репарации ДНК при прогрессировании рака желудочно-кишечного тракта». Всемирный журнал онкологии желудочно-кишечного тракта. 7 (5): 30–46. Дои:10.4251 / wjgo.v7.i5.30. ЧВК 4434036. PMID 25987950.
- ^ Maffioletti E, Tardito D, Gennarelli M, Bocchio-Chiavetto L (2014). «Микро-шпионы от мозга к периферии: новые ключи к разгадке в исследованиях микроРНК при нервно-психических расстройствах». Границы клеточной неврологии. 8: 75. Дои:10.3389 / fncel.2014.00075. ЧВК 3949217. PMID 24653674.
- ^ Меллиос Н., Сур М. (2012). «Растущая роль микроРНК при шизофрении и расстройствах аутистического спектра». Границы в психиатрии. 3: 39. Дои:10.3389 / fpsyt.2012.00039. ЧВК 3336189. PMID 22539927.
- ^ Гиган М., Кэрнс MJ (август 2015 г.). «МикроРНК и посттранскрипционная дисрегуляция в психиатрии». Биологическая психиатрия. 78 (4): 231–9. Дои:10.1016 / j.biopsych.2014.12.009. PMID 25636176.
- ^ Лю Кью, Рэнд Т.А., Калидас С., Ду Ф, Ким ХЭ, Смит Д.П., Ван Х (сентябрь 2003 г.). «R2D2, мост между инициирующей и эффекторной стадиями пути РНКи дрозофилы». Наука. 301 (5641): 1921–5. Bibcode:2003Наука ... 301.1921Л. Дои:10.1126 / science.1088710. PMID 14512631. S2CID 41436233.
- ^ Грегори Р.И., Чендримада Т.П., Куч Н., Шихаттар Р. (ноябрь 2005 г.). «Человеческий RISC сочетает биогенез микроРНК и посттранскрипционное молчание генов». Клетка. 123 (4): 631–40. Дои:10.1016 / j.cell.2005.10.022. PMID 16271387.
- ^ а б Лодиш Х, Берк А., Мацудаира П., Кайзер Калифорния, Кригер М., Скотт М. П., Зипуркси С. Л., Дарнелл Дж. (2004). Молекулярная клеточная биология (5-е изд.). WH Freeman: Нью-Йорк, штат Нью-Йорк. ISBN 978-0-7167-4366-8.
- ^ Матранга С., Томари Ю., Шин С., Бартель Д.П., Заморе П.П. (ноябрь 2005 г.). «Расщепление цепи-пассажира облегчает сборку siRNA в Ago2-содержащие ферментные комплексы РНКи». Клетка. 123 (4): 607–20. Дои:10.1016 / j.cell.2005.08.044. PMID 16271386.
- ^ Леушнер П.Дж., Амерес С.Л., Куэнг С., Мартинес Дж. (Март 2006 г.). «Расщепление пассажирской цепи миРНК во время сборки RISC в клетках человека». Отчеты EMBO. 7 (3): 314–20. Дои:10.1038 / sj.embor.7400637. ЧВК 1456892. PMID 16439995.
- ^ а б Хейли Б., Замор, PD (июль 2004 г.). «Кинетический анализ ферментного комплекса РНКи». Структурная и молекулярная биология природы. 11 (7): 599–606. Дои:10.1038 / nsmb780. PMID 15170178. S2CID 12400060.
- ^ Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (октябрь 2003 г.). «Асимметрия в сборке ферментного комплекса РНКи». Клетка. 115 (2): 199–208. Дои:10.1016 / S0092-8674 (03) 00759-1. PMID 14567917.
- ^ Preall JB, He Z, Gorra JM, Sontheimer EJ (март 2006 г.). «Отбор короткой интерферирующей цепи РНК не зависит от полярности процессинга дцРНК во время РНКи у дрозофилы». Текущая биология. 16 (5): 530–5. Дои:10.1016 / j.cub.2006.01.061. PMID 16527750.
- ^ Томари Ю., Матранга С., Хейли Б., Мартинес Н., Заморе П.Д. (ноябрь 2004 г.). «Белковый сенсор асимметрии миРНК». Наука. 306 (5700): 1377–80. Bibcode:2004Научный ... 306.1377T. Дои:10.1126 / science.1102755. PMID 15550672. S2CID 31558409.
- ^ Ма Дж. Б., Юань Ю. Р., Мейстер Дж., Пей И., Тушл Т., Патель Д. Д. (март 2005 г.). «Структурная основа для 5'-конца специфического распознавания направляющей РНК белком A. fulgidus Piwi». Природа. 434 (7033): 666–70. Bibcode:2005Натура.434..666М. Дои:10.1038 / природа03514. ЧВК 4694588. PMID 15800629.
- ^ Сен G, Wehrman T, Blau H (2005). «Трансляция мРНК не является предпосылкой для опосредованного малыми интерферирующими РНК расщепления мРНК». Дифференциация. 73 (6): 287–93. Дои:10.1111 / j.1432-0436.2005.00029.x. PMID 16138829. S2CID 41117614.
- ^ Гу С, Росси Дж (2005). «Отключение РНКи от активной трансляции в клетках млекопитающих». РНК. 11 (1): 38–44. Дои:10.1261 / rna.7158605. ЧВК 1370689. PMID 15574516.
- ^ Сен Дж., Блау Х (2005). «Argonaute 2 / RISC находится в сайтах распада мРНК млекопитающих, известных как цитоплазматические тельца». Nat Cell Biol. 7 (6): 633–6. Дои:10.1038 / ncb1265. PMID 15908945. S2CID 6085169.
- ^ Лиан С., Джакиму А., Эстатиой Т., Хамель Дж., Фритцлер М., Чан Э (2006). «Тельца GW, микроРНК и клеточный цикл». Клеточный цикл. 5 (3): 242–5. Дои:10.4161 / cc.5.3.2410. PMID 16418578.
- ^ Jakymiw A, Lian S, Eystathioy T, Li S, Satoh M, Hamel J, Fritzler M, Chan E (2005). «Нарушение Р-тел нарушает интерференцию РНК млекопитающих». Nat Cell Biol. 7 (12): 1267–74. Дои:10.1038 / ncb1334. PMID 16284622. S2CID 36630239.
- ^ Хаммонд S, Бернштейн E, Пляж D, Hannon G (2000). «РНК-направленная нуклеаза опосредует пост-транскрипционное молчание генов в клетках дрозофилы». Природа. 404 (6775): 293–6. Bibcode:2000Натура.404..293H. Дои:10.1038/35005107. PMID 10749213. S2CID 9091863.
- ^ Холмквист GP, Эшли Т. (2006). «Хромосомная организация и модификация хроматина: влияние на функцию и эволюцию генома». Цитогенетические и геномные исследования. 114 (2): 96–125. Дои:10.1159/000093326. PMID 16825762. S2CID 29910065.
- ^ Вердел А., Джиа С., Гербер С., Сугияма Т., Гайги С., Гревал С.И., Моазед Д. (январь 2004 г.). «РНКи-опосредованное нацеливание на гетерохроматин комплексом RITS». Наука. 303 (5658): 672–6. Bibcode:2004Наука ... 303..672В. Дои:10.1126 / science.1093686. ЧВК 3244756. PMID 14704433.
- ^ Ирвин Д.В., Заратиеги М., Толия Н.Х., Гото Д.Б., Читвуд Д.Х., Вон М.В., Джошуа-Тор Л., Мартиенсен РА (август 2006 г.). «Нарезка Argonaute требуется для гетерохроматического подавления и распространения». Наука. 313 (5790): 1134–7. Bibcode:2006Научный ... 313.1134I. Дои:10.1126 / science.1128813. PMID 16931764. S2CID 42997104.
- ^ Volpe TA, Kidner C, Hall IM, Teng G, Grewal SI, Martienssen RA (сентябрь 2002 г.). «Регулирование гетерохроматического сайленсинга и метилирования гистона H3 лизина-9 с помощью РНКи». Наука. 297 (5588): 1833–7. Bibcode:2002Научный ... 297.1833V. Дои:10.1126 / science.1074973. PMID 12193640. S2CID 2613813.
- ^ Volpe T, Schramke V, Hamilton GL, White SA, Teng G, Martienssen RA, Allshire RC (2003). «РНК-интерференция необходима для нормальной функции центромеры у делящихся дрожжей». Хромосомные исследования. 11 (2): 137–46. Дои:10.1023 / А: 1022815931524. PMID 12733640. S2CID 23813417.
- ^ Li LC, Okino ST, Zhao H, Pookot D, Place RF, Urakami S, Enokida H, Dahiya R (ноябрь 2006 г.). «Малые дцРНК вызывают активацию транскрипции в клетках человека». Труды Национальной академии наук Соединенных Штатов Америки. 103 (46): 17337–42. Bibcode:2006PNAS..10317337L. Дои:10.1073 / pnas.0607015103. ЧВК 1859931. PMID 17085592.
- ^ Нома К., Сугияма Т., Кам Х., Вердел А., Зофалл М., Джиа С., Моазед Д., Гревал С.И. (ноябрь 2004 г.). "RITS действует в цис-системе, способствуя опосредованному РНК-интерференцией транскрипционному и посттранскрипционному подавлению". Природа Генетика. 36 (11): 1174–80. Дои:10,1038 / ng1452. PMID 15475954.
- ^ Сугияма Т., Кам Х., Вердел А., Моазед Д., Гревал С.И. (январь 2005 г.). «РНК-зависимая РНК-полимераза является важным компонентом самоусиливающейся петли, связывающей сборку гетерохроматина с производством миРНК». Труды Национальной академии наук Соединенных Штатов Америки. 102 (1): 152–7. Bibcode:2005ПНАС..102..152С. Дои:10.1073 / pnas.0407641102. ЧВК 544066. PMID 15615848.
- ^ Ван Ф., Кояма Н., Нисида Х., Харагути Т., Рейт В., Цукамото Т. (июнь 2006 г.). «Сборка и поддержание гетерохроматина, инициированное повторами трансгена, не зависит от пути интерференции РНК в клетках млекопитающих». Молекулярная и клеточная биология. 26 (11): 4028–40. Дои:10.1128 / MCB.02189-05. ЧВК 1489094. PMID 16705157.
- ^ Бас БЛ (2002). «Редактирование РНК аденозиндезаминазами, действующими на РНК». Ежегодный обзор биохимии. 71: 817–46. Дои:10.1146 / annurev.biochem.71.110601.135501. ЧВК 1823043. PMID 12045112.
- ^ Bass BL (апрель 2000 г.). «Двухцепочечная РНК как матрица для сайленсинга генов». Клетка. 101 (3): 235–8. Дои:10.1016 / S0092-8674 (02) 71133-1. PMID 10847677.
- ^ Лучано DJ, Мирский H, Vendetti NJ, Maas S (август 2004 г.). «Редактирование РНК предшественника миРНК». РНК. 10 (8): 1174–7. Дои:10.1261 / rna.7350304. ЧВК 1370607. PMID 15272117.
- ^ а б Ян В., Чендримада Т.П., Ван Ц., Хигучи М., Зеебург П.Х., Шихаттар Р., Нишикура К. (январь 2006 г.). «Модуляция процессинга и экспрессии микроРНК посредством редактирования РНК дезаминазами ADAR». Структурная и молекулярная биология природы. 13 (1): 13–21. Дои:10.1038 / nsmb1041. ЧВК 2950615. PMID 16369484.
- ^ Ян В., Ван Кью, Хауэлл К.Л., Ли Дж. Т., Чо Д.С., Мюррей Дж. М., Нишикура К. (февраль 2005 г.). «ADAR1 РНК дезаминаза ограничивает эффективность короткой интерферирующей РНК в клетках млекопитающих». Журнал биологической химии. 280 (5): 3946–53. Дои:10.1074 / jbc.M407876200. ЧВК 2947832. PMID 15556947.
- ^ Нисикура К. (декабрь 2006 г.). «Редактор встречает глушитель: перекрестные помехи между редактированием РНК и вмешательством РНК». Обзоры природы Молекулярная клеточная биология. 7 (12): 919–31. Дои:10.1038 / nrm2061. ЧВК 2953463. PMID 17139332.
- ^ а б c Саумет А., Леселье СН (2006). «Антивирусное подавление РНК: мы похожи на растения?». Ретровирология. 3 (1): 3. Дои:10.1186/1742-4690-3-3. ЧВК 1363733. PMID 16409629.
- ^ Джонс Л., Рэтклифф Ф., Баулкомб, округ Колумбия (май 2001 г.). «РНК-направленное подавление транскрипционного гена у растений может быть унаследовано независимо от триггера РНК и требует Met1 для поддержания». Текущая биология. 11 (10): 747–57. Дои:10.1016 / S0960-9822 (01) 00226-3. PMID 11378384. S2CID 16789197.
- ^ Хамфрис Д.Т., Вестман Б.Дж., Мартин Д.И., Прейсс Т. (ноябрь 2005 г.). «МикроРНК контролируют инициацию трансляции, ингибируя эукариотический фактор инициации 4E / cap и функцию поли (A) хвоста». Труды Национальной академии наук Соединенных Штатов Америки. 102 (47): 16961–6. Bibcode:2005PNAS..10216961H. Дои:10.1073 / pnas.0506482102. ЧВК 1287990. PMID 16287976.
- ^ ДаРоча В.Д., Оцу К., Тейшейра С.М., Донельсон Дж. Э. (февраль 2004 г.). «Тесты цитоплазматической РНК-интерференции (РНКи) и конструирование индуцируемой тетрациклином системы промотора Т7 в Trypanosoma cruzi». Молекулярная и биохимическая паразитология. 133 (2): 175–86. Дои:10.1016 / j.molbiopara.2003.10.005. PMID 14698430.
- ^ Робинсон К.А., Беверли С.М. (май 2003 г.). «Повышение эффективности трансфекции и тестирование подходов к РНК-интерференции (РНКи) у простейших паразитов Leishmania». Молекулярная и биохимическая паразитология. 128 (2): 217–28. Дои:10.1016 / S0166-6851 (03) 00079-3. PMID 12742588.
- ^ Аравинд Л., Ватанабе Х, Липман Д. Д., Кунин Е. В. (октябрь 2000 г.). «Потеря и расхождение функционально связанных генов у эукариот». Труды Национальной академии наук Соединенных Штатов Америки. 97 (21): 11319–24. Bibcode:2000PNAS ... 9711319A. Дои:10.1073 / pnas.200346997. ЧВК 17198. PMID 11016957.
- ^ Дринненберг И.А., Вайнберг Д.Е., Се К.Т., Косилка Дж. П., Вулф К. Х., Финк Г. Р., Бартель Д. П. (октябрь 2009 г.). «РНКи в почкующихся дрожжах». Наука. 326 (5952): 544–550. Bibcode:2009Наука ... 326..544D. Дои:10.1126 / science.1176945. ЧВК 3786161. PMID 19745116.
- ^ Накаяшики Х., Кадотани Н., Маяма С. (июль 2006 г.). «Эволюция и диверсификация белков, подавляющих РНК, в грибах» (PDF). Журнал молекулярной эволюции. 63 (1): 127–35. Bibcode:2006JMolE..63..127N. Дои:10.1007 / s00239-005-0257-2. PMID 16786437. S2CID 22639035.
- ^ Морита Т., Мотидзуки Ю., Айба Х (март 2006 г.). «Репрессия трансляции достаточна для подавления гена небольшими некодирующими РНК бактерий в отсутствие разрушения мРНК». Труды Национальной академии наук Соединенных Штатов Америки. 103 (13): 4858–63. Bibcode:2006ПНАС..103.4858М. Дои:10.1073 / pnas.0509638103. ЧВК 1458760. PMID 16549791.
- ^ Макарова К.С., Гришин Н.В., Шабалина С.А., Вольф Ю.И., Кунин Е.В. (март 2006 г.). «Предполагаемая иммунная система прокариот, основанная на РНК-интерференции: компьютерный анализ предсказанного ферментативного механизма, функциональные аналогии с эукариотической РНКи и гипотетические механизмы действия». Биология Директ. 1: 7. Дои:10.1186/1745-6150-1-7. ЧВК 1462988. PMID 16545108.
- ^ Страм Ю., Кузнцова Л. (июнь 2006 г.). «Ингибирование вирусов посредством РНК-интерференции». Гены вирусов. 32 (3): 299–306. Дои:10.1007 / s11262-005-6914-0. ЧВК 7088519. PMID 16732482.
- ^ Блевинс Т., Раджешваран Р., Шивапрасад П.В., Бекназариантс Д., Си-Аммор А., Парк Х.С., Васкес Ф., Робертсон Д., Мейнс Ф., Хон Т., Пуггин М.М. (2006). «Четыре дайсера растений опосредуют биогенез вирусной малой РНК и подавление индуцированного ДНК вируса». Исследования нуклеиновых кислот. 34 (21): 6233–46. Дои:10.1093 / нар / gkl886. ЧВК 1669714. PMID 17090584.
- ^ Palauqui JC, Elmayan T, Pollien JM, Vaucheret H (август 1997 г.). «Системное приобретенное молчание: посттранскрипционное молчание, специфичное для трансгена, передается путем прививки от молчаливого стада к потомству без молчания». Журнал EMBO. 16 (15): 4738–45. Дои:10.1093 / emboj / 16.15.4738. ЧВК 1170100. PMID 9303318.
- ^ Воиннет О (август 2001 г.). «РНК сайленсинг как иммунная система растений против вирусов». Тенденции в генетике. 17 (8): 449–59. Дои:10.1016 / S0168-9525 (01) 02367-8. PMID 11485817.
- ^ а б Люси А.П., Го Х.С., Ли В.Х., Дин С.В. (апрель 2000 г.). «Подавление посттранскрипционного сайленсинга генов растительным вирусным белком, локализованным в ядре». Журнал EMBO. 19 (7): 1672–80. Дои:10.1093 / emboj / 19.7.1672. ЧВК 310235. PMID 10747034.
- ^ Mérai Z, Kerényi Z, Kertész S, Magna M, Lakatos L, Silhavy D (июнь 2006 г.). «Связывание двухцепочечной РНК может быть общей вирусной стратегией растительной РНК для подавления молчания РНК». Журнал вирусологии. 80 (12): 5747–56. Дои:10.1128 / JVI.01963-05. ЧВК 1472586. PMID 16731914.
- ^ Катияр-Агарвал С., Морган Р., Дальбек Д., Борсани О., Виллегас А., Жу Дж. К., Стаскавич Б. Дж., Джин Х. (ноябрь 2006 г.). «Патоген-индуцируемая эндогенная миРНК в иммунитете растений». Труды Национальной академии наук Соединенных Штатов Америки. 103 (47): 18002–7. Bibcode:2006PNAS..10318002K. Дои:10.1073 / pnas.0608258103. ЧВК 1693862. PMID 17071740.
- ^ Фриц JH, Girardin SE, Philpott DJ (июнь 2006 г.). «Врожденная иммунная защита через вмешательство РНК». STKE науки. 2006 (339): pe27. Дои:10.1126 / stke.3392006pe27. PMID 16772641. S2CID 33972766.
- ^ Замбон Р.А., Вахария В.Н., Ву Л.П. (май 2006 г.). «РНКи представляет собой противовирусный иммунный ответ против вируса дцРНК у Drosophila melanogaster». Клеточная микробиология. 8 (5): 880–9. Дои:10.1111 / j.1462-5822.2006.00688.x. PMID 16611236. S2CID 32439482.
- ^ Ван XH, Алияри Р., Ли В.X., Ли Х.В., Ким К., Картью Р., Аткинсон П., Дин С.В. (апрель 2006 г.). «РНК-интерференция управляет врожденным иммунитетом против вирусов у взрослых дрозофил». Наука. 312 (5772): 452–4. Bibcode:2006Sci ... 312..452W. Дои:10.1126 / science.1125694. ЧВК 1509097. PMID 16556799.
- ^ Лу Р, Мадуро М., Ли Ф, Ли Х. У., Бройтман-Мадуро Дж., Ли У. X, Дин С.В. (август 2005 г.). «Репликация животных вирусов и РНКи-опосредованное противовирусное подавление у Caenorhabditis elegans». Природа. 436 (7053): 1040–1043. Bibcode:2005Натура 436.1040Л. Дои:10.1038 / природа03870. ЧВК 1388260. PMID 16107851.
- ^ Уилкинс С., Дишонг Р., Мур С.К., Уитт М.А., Чоу М., Мачака К. (август 2005 г.). «РНК-интерференция - это механизм противовирусной защиты у Caenorhabditis elegans». Природа. 436 (7053): 1044–7. Bibcode:2005 Натур.436.1044W. Дои:10.1038 / природа03957. PMID 16107852. S2CID 4431035.
- ^ Беркхут Б., Хааснот Дж. (Май 2006 г.). «Взаимодействие между вирусной инфекцией и механизмом клеточной РНК-интерференции». Письма FEBS. 580 (12): 2896–902. Дои:10.1016 / j.febslet.2006.02.070. ЧВК 7094296. PMID 16563388.
- ^ Шютц С., Сарнов П. (январь 2006 г.). «Взаимодействие вирусов с путем интерференции РНК млекопитающих». Вирусология. 344 (1): 151–7. Дои:10.1016 / j.virol.2005.09.034. PMID 16364746.
- ^ Каллен Б.Р. (июнь 2006 г.). «Участвует ли РНК-интерференция во внутреннем противовирусном иммунитете у млекопитающих?». Иммунология природы. 7 (6): 563–7. Дои:10.1038 / ni1352. PMID 16715068. S2CID 23467688.
- ^ Майяр П.В., Чаудо С., Марше А., Ли И, Джей Ф., Динг С.В., Воиннет О. (октябрь 2013 г.). «Противовирусная РНК-интерференция в клетках млекопитающих». Наука. 342 (6155): 235–8. Bibcode:2013Наука ... 342..235М. Дои:10.1126 / science.1241930. ЧВК 3853215. PMID 24115438.
- ^ Ли И, Лу Дж, Хан И, Фан Х, Дин С.В. (октябрь 2013 г.). «РНК-интерференция действует как механизм противовирусного иммунитета у млекопитающих». Наука. 342 (6155): 231–4. Bibcode:2013Наука ... 342..231Л. Дои:10.1126 / science.1241911. ЧВК 3875315. PMID 24115437.
- ^ Ли Х.В., Дин С.В. (октябрь 2005 г.). «Антивирусное подавление у животных». Письма FEBS. 579 (26): 5965–73. Дои:10.1016 / j.febslet.2005.08.034. ЧВК 1350842. PMID 16154568.
- ^ Кэррингтон Дж. К., Амброс V (июль 2003 г.). «Роль микроРНК в развитии растений и животных». Наука. 301 (5631): 336–8. Bibcode:2003Наука ... 301..336C. Дои:10.1126 / science.1085242. PMID 12869753. S2CID 43395657.
- ^ Ли Р.К., Фейнбаум Р.Л., Амброс V (декабрь 1993 г.). «Гетерохронный ген lin-4 C. elegans кодирует малые РНК с антисмысловой комплементарностью lin-14». Клетка. 75 (5): 843–54. Дои:10.1016 / 0092-8674 (93) 90529-У. PMID 8252621.
- ^ Палатник Дж. Ф., Аллен Э., Ву Х, Шоммер С., Шваб Р., Каррингтон Дж. К., Вейгель Д. (сентябрь 2003 г.). «Контроль морфогенеза листа с помощью микроРНК». Природа. 425 (6955): 257–63. Bibcode:2003Натура.425..257П. Дои:10.1038 / природа01958. PMID 12931144. S2CID 992057.
- ^ Чжан Б., Пан X, Кобб Г.П., Андерсон Т.А. (январь 2006 г.). «Растительная микроРНК: небольшая регуляторная молекула с большим влиянием». Биология развития. 289 (1): 3–16. Дои:10.1016 / j.ydbio.2005.10.036. PMID 16325172.
- ^ Джонс-Роудс MW, Бартель Д.П., Бартель Б (2006). «MicroRNAS и их регуляторная роль в растениях». Ежегодный обзор биологии растений. 57: 19–53. Дои:10.1146 / annurev.arplant.57.032905.105218. PMID 16669754. S2CID 13010154.
- ^ Чжан Б., Пан X, Кобб Г.П., Андерсон Т.А. (февраль 2007 г.). «микроРНК как онкогены и опухолевые супрессоры». Биология развития. 302 (1): 1–12. Дои:10.1016 / j.ydbio.2006.08.028. PMID 16989803.
- ^ а б c Cerutti H, Casas-Mollano JA (август 2006 г.). «О происхождении и функциях РНК-опосредованного сайленсинга: от протистов к человеку». Текущая генетика. 50 (2): 81–99. Дои:10.1007 / s00294-006-0078-х. ЧВК 2583075. PMID 16691418.
- ^ Анантараман В., Кунин Е.В., Аравинд Л. (апрель 2002 г.). «Сравнительная геномика и эволюция белков, участвующих в метаболизме РНК». Исследования нуклеиновых кислот. 30 (7): 1427–64. Дои:10.1093 / nar / 30.7.1427. ЧВК 101826. PMID 11917006.
- ^ Бучон Н., Ваури С. (февраль 2006 г.). «РНКи: защитное молчание РНК против вирусов и мобильных элементов». Наследственность. 96 (2): 195–202. Дои:10.1038 / sj.hdy.6800789. PMID 16369574.
- ^ Obbard DJ, Jiggins FM, Halligan DL, Little TJ (март 2006 г.). «Естественный отбор способствует чрезвычайно быстрой эволюции генов противовирусной РНКи». Текущая биология. 16 (6): 580–5. Дои:10.1016 / j.cub.2006.01.065. PMID 16546082.
- ^ Вурхув П.М., Агами Р. (январь 2003 г.). «Нокдаун встает». Тенденции в биотехнологии. 21 (1): 2–4. Дои:10.1016 / S0167-7799 (02) 00002-1. PMID 12480342.
- ^ Мункачи Г., Штупински З., Герман П., Бан Б., Пензвальто З, Сарвас Н., Дьёрфи Б. (сентябрь 2016 г.). «Подтверждение эффективности подавления РНКи с использованием данных массива генов показывает 18,5% отказов в 429 независимых экспериментах». Молекулярная терапия. Нуклеиновые кислоты. 5 (9): e366. Дои:10.1038 / mtna.2016.66. ЧВК 5056990. PMID 27673562.
- ^ Наито Ю., Ямада Т., Мацумия Т., Уи-Тей К., Сайго К., Моришита С. (июль 2005 г.). «dsCheck: высокочувствительное программное обеспечение для поиска вне мишени для двухцепочечной РНК-опосредованной РНК-интерференции». Исследования нуклеиновых кислот. 33 (Выпуск веб-сервера): W589–91. Дои:10.1093 / нар / gki419. ЧВК 1160180. PMID 15980542.
- ^ Henschel A, Buchholz F, Habermann B (июль 2004 г.). «DEQOR: веб-инструмент для разработки и контроля качества миРНК». Исследования нуклеиновых кислот. 32 (Выпуск веб-сервера): W113–20. Дои:10.1093 / нар / гх408. ЧВК 441546. PMID 15215362.
- ^ Наито Ю., Ямада Т., Уи-Тей К., Моришита С., Сайго К. (июль 2004 г.). «siDirect: высокоэффективное программное обеспечение для создания целевой siRNA для интерференции РНК млекопитающих». Исследования нуклеиновых кислот. 32 (Выпуск веб-сервера): W124–9. Дои:10.1093 / нар / гх442. ЧВК 441580. PMID 15215364.
- ^ Наито Ю., Уи-Тей К., Нисикава Т., Такебе Ю., Сайго К. (июль 2006 г.). «siVirus: веб-программа для разработки антивирусных siRNA для сильно расходящихся вирусных последовательностей». Исследования нуклеиновых кислот. 34 (Выпуск веб-сервера): W448–50. Дои:10.1093 / нар / gkl214. ЧВК 1538817. PMID 16845046.
- ^ Рейнольдс А., Андерсон Э.М., Вермёлен А., Федоров Ю., Робинсон К., Лик Д., Карпилов Дж., Маршалл В.С., Хворова А. (июнь 2006 г.). «Индукция интерферонового ответа миРНК зависит от типа клетки и длины дуплекса». РНК. 12 (6): 988–93. Дои:10.1261 / rna.2340906. ЧВК 1464853. PMID 16611941.
- ^ Штейн П., Цзэн Ф., Пан Х., Шульц Р.М. (октябрь 2005 г.). «Отсутствие неспецифических эффектов РНК-интерференции, вызванной длинной двухцепочечной РНК в ооцитах мышей». Биология развития. 286 (2): 464–71. Дои:10.1016 / j.ydbio.2005.08.015. PMID 16154556.
- ^ Бруммелькамп TR, Бернардс Р, Агами Р. (апрель 2002 г.). «Система стабильной экспрессии коротких интерферирующих РНК в клетках млекопитающих». Наука. 296 (5567): 550–3. Bibcode:2002Наука ... 296..550B. Дои:10.1126 / science.1068999. HDL:1874/15573. PMID 11910072. S2CID 18460980.
- ^ Тискорния Г., Тергаонкар В., Галими Ф., Верма И.М. (май 2004 г.). «Интерференция РНК, индуцируемая CRE-рекомбиназой, опосредованная лентивирусными векторами». Труды Национальной академии наук Соединенных Штатов Америки. 101 (19): 7347–51. Bibcode:2004ПНАС..101.7347Т. Дои:10.1073 / pnas.0402107101. ЧВК 409921. PMID 15123829.
- ^ Вентура А., Мейснер А., Диллон С.П., МакМанус М., Шарп П.А., Ван Парижс Л., Яениш Р., Джекс Т. (июль 2004 г.). «Cre-lox-регулируемая условная интерференция РНК трансгенов». Труды Национальной академии наук Соединенных Штатов Америки. 101 (28): 10380–5. Bibcode:2004ПНАС..10110380В. Дои:10.1073 / pnas.0403954101. ЧВК 478580. PMID 15240889.
- ^ Камат Р.С., Арингер Дж. (Август 2003 г.). «Полногеномный скрининг РНКи у Caenorhabditis elegans». Методы. 30 (4): 313–21. Дои:10.1016 / S1046-2023 (03) 00050-1. PMID 12828945.
- ^ Бутрос М., Кигер А.А., Армкнехт С., Керр К., Хилд М., Кох Б., Хаас С.А., Паро Р., Перримон Н. (февраль 2004 г.). «Полногеномный РНКи-анализ роста и жизнеспособности клеток дрозофилы». Наука. 303 (5659): 832–5. Bibcode:2004Наука ... 303..832B. Дои:10.1126 / science.1091266. PMID 14764878. S2CID 7289808.
- ^ Фортунато А., Фрейзер А.Г. (2005). «Раскройте генетические взаимодействия у Caenorhabditis elegans с помощью РНК-интерференции». Отчеты по бионауке. 25 (5–6): 299–307. Дои:10.1007 / s10540-005-2892-7. PMID 16307378. S2CID 6983519.
- ^ Каллен Л. М., Арндт Г. М. (июнь 2005 г.). «Полногеномный скрининг функции генов с использованием РНКи в клетках млекопитающих». Иммунология и клеточная биология. 83 (3): 217–23. Дои:10.1111 / j.1440-1711.2005.01332.x. PMID 15877598. S2CID 10443862.
- ^ Huesken D, Lange J, Mickanin C, Weiler J, Asselbergs F, Warner J, Meloon B, Engel S, Rosenberg A, Cohen D, Labow M, Reinhardt M, Natt F, Hall J (август 2005 г.). «Дизайн библиотеки миРНК для всего генома с использованием искусственной нейронной сети». Природа Биотехнологии. 23 (8): 995–1001. Дои:10.1038 / nbt1118. PMID 16025102. S2CID 11030533.
- ^ Ге Г., Вонг Г. В., Ло Б. (октябрь 2005 г.). «Прогнозирование эффективности нокдауна siRNA с использованием моделей искусственной нейронной сети». Сообщения о биохимических и биофизических исследованиях. 336 (2): 723–8. Дои:10.1016 / j.bbrc.2005.08.147. PMID 16153609.
- ^ Яниц М., Ванхеке Д., Лехрах Х (2006). «Высокопроизводительное вмешательство РНК в функциональной геномике». РНК к медицине. Справочник по экспериментальной фармакологии. 173. С. 97–104. Дои:10.1007/3-540-27262-3_5. ISBN 978-3-540-27261-8. PMID 16594612.
- ^ Ванхеке Д., Яниц М. (февраль 2005 г.). «Функциональная геномика с использованием высокопроизводительной РНК-интерференции». Открытие наркотиков сегодня. 10 (3): 205–12. Дои:10.1016 / S1359-6446 (04) 03352-5. HDL:11858 / 00-001M-0000-0010-86E7-8. PMID 15708535.
- ^ Гельдхоф П., Мюррей Л., Кутье А., Жиллард Дж. С., Маклаучлан Г., Нокс Д. П., Бриттон С. (июнь 2006 г.). «Проверка эффективности вмешательства РНК в Haemonchus contortus». Международный журнал паразитологии. 36 (7): 801–10. Дои:10.1016 / j.ijpara.2005.12.004. PMID 16469321.
- ^ Гельдхоф П., Виссер А., Кларк Д., Сондерс Г., Бриттон С., Жиллард Дж., Берриман М., Нокс Д. (май 2007 г.). «РНК-интерференция у паразитических гельминтов: текущая ситуация, возможные подводные камни и перспективы на будущее». Паразитология. 134 (Pt 5): 609–19. Дои:10.1017 / S0031182006002071. PMID 17201997.
- ^ Travella S, Klimm TE, Keller B (сентябрь 2006 г.). «Молчание генов на основе РНК-интерференции как эффективный инструмент функциональной геномики в гексаплоидной мягкой пшенице». Физиология растений. 142 (1): 6–20. Дои:10.1104 / стр. 106.084517. ЧВК 1557595. PMID 16861570.
- ^ Макгиннис К., Чендлер В., Конус К., Кэпплер Х., Кэпплер С., Кершен А., Пикаард С., Ричардс Э., Сидоренко Л., Смит Т., Спрингер Н., Вулан Т. (2005). «Индуцированная трансгеном интерференция РНК как инструмент функциональной геномики растений». РНК интерференция. Методы в энзимологии. 392. С. 1–24. Дои:10.1016 / S0076-6879 (04) 92001-0. ISBN 978-0-12-182797-7. PMID 15644172.
- ^ а б Сен GL, Blau HM (июль 2006 г.). «Краткая история RNAi: молчание генов». Журнал FASEB. 20 (9): 1293–9. Дои:10.1096 / fj.06-6014rev. PMID 16816104. S2CID 12917676.
- ^ Данехольт Б. (2 октября 2006 г.). «Нобелевская премия по физиологии и медицине 2006 г.». nobelprize.org. Получено 30 октября 2017.
- ^ Эльбашир С., Харборт Дж., Лендекель В. и др. (2001). «Дуплексы 21-нуклеотидной РНК опосредуют РНК-интерференцию в культивируемых клетках млекопитающих». Природа. 411 (6836): 494–498. Bibcode:2001Натура.411..494E. Дои:10.1038/35078107. PMID 11373684. S2CID 710341.
- ^ McCaffrey AP, Meuse L, Pham TT, Conklin DS, Hannon GJ, Кей М.А. (июль 2002 г.). «РНК-интерференция у взрослых мышей». Природа. 418 (6893): 38–9. Bibcode:2002Натура.418 ... 38М. Дои:10.1038 / 418038a. PMID 12097900. S2CID 4361399.
- ^ Devi GR (сентябрь 2006 г.). "подходы на основе siRNA в терапии рака". Генная терапия рака. 13 (9): 819–29. Дои:10.1038 / sj.cgt.7700931. PMID 16424918.
- ^ Уолл Н.Р., Ши И (октябрь 2003 г.). «Малая РНК: можно ли использовать РНК-интерференцию в терапии?». Ланцет. 362 (9393): 1401–3. Дои:10.1016 / с0140-6736 (03) 14637-5. PMID 14585643. S2CID 25034627.
- ^ Сах Д (2006). «Терапевтический потенциал РНК-интерференции при неврологических расстройствах». Life Sci. 79 (19): 1773–80. Дои:10.1016 / j.lfs.2006.06.011. PMID 16815477.
- ^ Дэвис М.Э., Цукерман Дж. Э., Чой СН, Селигсон Д., Толчер А., Алаби, Калифорния, Йен Й., Хайдель Д. Д., Рибас А. (апрель 2010 г.). «Доказательства наличия РНКи у людей при системном введении миРНК через наночастицы-мишени». Природа. 464 (7291): 1067–70. Bibcode:2010 Натур.464.1067D. Дои:10.1038 / природа08956. ЧВК 2855406. PMID 20305636.
- ^ Эскобар М.А., Чивероло Е.Л., Саммерфельт КР, Дандекар А.М. (ноябрь 2001 г.). «РНКи-опосредованное подавление онкогенов придает устойчивость к онкогенезу коронного галла». Труды Национальной академии наук Соединенных Штатов Америки. 98 (23): 13437–42. Bibcode:2001PNAS ... 9813437E. Дои:10.1073 / pnas.241276898. ЧВК 60889. PMID 11687652.
- ^ Перейра Т.К., Паскоал В.Д., Марчезини Р.Б., Майя И.Г., Магальяйнс Л.А., Занотти-Магальяйнс Е.М., Лопес-Сендес I (апрель 2008 г.). «Schistosoma mansoni: оценка лечения на основе РНКи, направленного на ген HGPRTase». Экспериментальная паразитология. 118 (4): 619–23. Дои:10.1016 / j.exppara.2007.11.017. PMID 18237732.
- ^ Рауль С., Аббас-Терки Т., Бенсадун Дж. К., Гийо С., Хаасе Дж., Шульц Дж., Хендерсон К. Э., Эбишер П. (апрель 2005 г.). «Опосредованное лентивирусами подавление SOD1 посредством РНК-интерференции замедляет начало и прогрессирование болезни в мышиной модели БАС». Природа Медицина. 11 (4): 423–8. Дои:10,1038 / нм1207. PMID 15768028. S2CID 25445264.
- ^ Пташник А., Наката Ю., Калота А., Эмерсон С. Г., Гевиртц А. М. (ноябрь 2004 г.). «Короткая интерферирующая РНК (миРНК), нацеленная на Lyn киназу, индуцирует апоптоз в первичных и устойчивых к лекарствам клетках лейкемии BCR-ABL1 (+)». Природа Медицина. 10 (11): 1187–9. Дои:10,1038 / нм1127. PMID 15502840. S2CID 21770360.
- ^ Kim SJ, Lee WI, Lee YS, Kim DH, Chang JW, Kim SW, Lee H (ноябрь 2009 г.). «Эффективное облегчение нейропатической боли за счет опосредованной аденоассоциированным вирусом экспрессии небольшой шпильочной РНК против GTP циклогидролазы 1». Молекулярная боль. 5: 1744–8069–5–67. Дои:10.1186/1744-8069-5-67. ЧВК 2785765. PMID 19922668.
- ^ Чен Л., МакКенна Дж. Т., Болортуя Ю., Уинстон С., Таккар М. М., Башир Р., Браун Р. Э., Маккарли Р. У. (ноябрь 2010 г.). «Нокдаун рецептора орексина типа 1 в голубом пятне крысы увеличивает быстрый сон в темное время суток». Европейский журнал нейробиологии. 32 (9): 1528–36. Дои:10.1111 / j.1460-9568.2010.07401.x. ЧВК 3058252. PMID 21089218.
- ^ Vargason JM, Szittya G, Burgyán J, Hall TM (декабрь 2003 г.). «Выборочное распознавание siRNA супрессором сайленсинга РНК». Клетка. 115 (7): 799–811. Дои:10.1016 / s0092-8674 (03) 00984-х. PMID 14697199.
- ^ Беркхут Б (апрель 2004 г.). «РНК-интерференция как противовирусный подход: нацеливание на ВИЧ-1». Современное мнение о молекулярной терапии. 6 (2): 141–5. PMID 15195925.
- ^ Цзян М., Милнер Дж. (Сентябрь 2002 г.). «Селективное подавление экспрессии вирусных генов в ВПЧ-положительных клетках карциномы шейки матки человека, обработанных миРНК, праймером РНК-интерференции». Онкоген. 21 (39): 6041–8. Дои:10.1038 / sj.onc.1205878. PMID 12203116.
- ^ Кусов Ю., Канда Т., Пальменберг А., Сгро Ю.Ю., Гаусс-Мюллер В. (июнь 2006 г.). «Подавление инфекции вируса гепатита А с помощью малых интерферирующих РНК». Журнал вирусологии. 80 (11): 5599–610. Дои:10.1128 / jvi.01773-05. ЧВК 1472172. PMID 16699041.
- ^ Цзя Ф., Чжан Ю.З., Лю С.М. (октябрь 2006 г.). «Система на основе ретровирусов, позволяющая стабильно заглушить гены вируса гепатита В с помощью РНК-интерференции». Письма о биотехнологии. 28 (20): 1679–85. Дои:10.1007 / s10529-006-9138-z. PMID 16900331. S2CID 34511611.
- ^ Ли Ю.К., Конг Л.Х., Ченг Б.З., Ли К.С. (декабрь 2005 г.). «Конструирование векторов экспрессии siRNA вируса гриппа и их ингибирующие эффекты на размножение вируса гриппа». Заболевания птиц. 49 (4): 562–73. Дои:10.1637 / 7365-041205R2.1. PMID 16405000. S2CID 86214047.
- ^ Кханна М., Саксена Л., Раджпут Р., Кумар Б., Прасад Р. (2015). «Замалчивание генов: терапевтический подход к борьбе с инфекциями вируса гриппа». Будущая микробиология. 10 (1): 131–40. Дои:10.2217 / fmb.14.94. PMID 25598342.
- ^ Раджпут Р., Кханна М., Кумар П., Кумар Б., Шарма С., Гупта Н., Саксена Л. (декабрь 2012 г.). «Малая интерферирующая РНК, нацеленная на транскрипт неструктурного гена 1, подавляет репликацию вируса гриппа А у экспериментальных мышей». Нуклеиновые кислоты. 22 (6): 414–22. Дои:10.1089 / нат.2012.0359. PMID 23062009.
- ^ а б c d Аша К., Кумар П., Саникас М., Месеко К.А., Кханна М., Кумар Б. (декабрь 2018 г.). «Достижения в терапии на основе нуклеиновых кислот против респираторных вирусных инфекций». Журнал клинической медицины. 8 (1): 6. Дои:10.3390 / см 8010006. ЧВК 6351902. PMID 30577479.
- ^ Ху Л., Ван З., Ху Ц., Лю Х, Яо Л., Ли В., Ци Ю. (2005). «Ингибирование размножения вируса кори в культуре клеток путем РНК-интерференции». Acta Virologica. 49 (4): 227–34. PMID 16402679.
- ^ Куреши А., Такур Н., Монга И., Такур А., Кумар М. (2014). «VIRmiRNA: исчерпывающий ресурс для экспериментально подтвержденных вирусных miRNA и их мишеней». База данных. 2014. Дои:10.1093 / база данных / bau103. ЧВК 4224276. PMID 25380780.
- ^ Кроу С (2003). «Подавление экспрессии хемокиновых рецепторов посредством РНК-интерференции позволяет ингибировать репликацию ВИЧ-1, Мартинес и др.». СПИД. 17 Дополнение 4: S103–5. PMID 15080188.
- ^ Fuchs U, Damm-Welk C, Borkhardt A (август 2004 г.). «Молчание генов, связанных с заболеванием, с помощью малых интерферирующих РНК». Современная молекулярная медицина. 4 (5): 507–17. Дои:10.2174/1566524043360492. PMID 15267222.
- ^ Чока Д.П., Аоки Ю., Киёсава К. (февраль 2003 г.). «РНК-интерференция - это функциональный путь с терапевтическим потенциалом в клеточных линиях миелоидного лейкоза человека». Генная терапия рака. 10 (2): 125–33. Дои:10.1038 / sj.cgt.7700544. PMID 12536201.
- ^ Лаптева Н., Ян А.Г., Сандерс Д.Е., Штрубе Р.В., Чен С.Ю. (январь 2005 г.). «Нокдаун CXCR4 малой интерферирующей РНК отменяет рост опухоли груди in vivo». Генная терапия рака. 12 (1): 84–9. Дои:10.1038 / sj.cgt.7700770. PMID 15472715.
- ^ Певица О, Марр Р.А., Рокенштейн Э., Крюс Л., Куфаль Н.Г., Гейдж Ф.Х., Верма И.М., Маслия Э. (октябрь 2005 г.). «Нацеливание на BACE1 с помощью siRNA улучшает невропатологию болезни Альцгеймера в трансгенной модели». Природа Неврология. 8 (10): 1343–9. Дои:10.1038 / nn1531. PMID 16136043. S2CID 6978101.
- ^ Родригес-Леброн Э., Гувион К.М., Мур С.А., Дэвидсон Б.Л., Полсон Х.Л. (сентябрь 2009 г.). «Аллель-специфическая РНКи смягчает фенотипическое прогрессирование в трансгенной модели болезни Альцгеймера». Молекулярная терапия. 17 (9): 1563–73. Дои:10.1038 / мт.2009.123. ЧВК 2835271. PMID 19532137.
- ^ Пьедрахита Д., Эрнандес И., Лопес-Тобон А., Федоров Д., Обара Б., Манджунат Б.С., Будро Р.Л., Дэвидсон Б., Лаферла Ф., Гальего-Гомес Й.С., Косик К.С., Кардона-Гомес, Гран При (октябрь 2010 г.). «Выключение CDK5 уменьшает нейрофибриллярные клубки у трансгенных мышей с болезнью Альцгеймера». Журнал неврологии. 30 (42): 13966–76. Дои:10.1523 / jneurosci.3637-10.2010. ЧВК 3003593. PMID 20962218.
- ^ Рауль С., Баркер С.Д., Эбишер П. (март 2006 г.). «Вирусное моделирование и коррекция нейродегенеративных заболеваний с помощью РНК-интерференции». Генная терапия. 13 (6): 487–95. Дои:10.1038 / sj.gt.3302690. PMID 16319945.
- ^ Harper SQ, Staber PD, He X, Eliason SL, Martins IH, Mao Q, Yang L, Kotin RM, Paulson HL, Davidson BL (апрель 2005 г.). «РНК-интерференция улучшает моторные и нейропатологические аномалии на мышиной модели с болезнью Гентингтона». Труды Национальной академии наук Соединенных Штатов Америки. 102 (16): 5820–5. Bibcode:2005ПНАС..102.5820Н. Дои:10.1073 / pnas.0501507102. ЧВК 556303. PMID 15811941.
- ^ Будро Р.Л., Родригес-Леброн Э., Дэвидсон Б.Л. (апрель 2011 г.). «РНКи-медицина для мозга: достижения и проблемы». Молекулярная генетика человека. 20 (R1): R21–7. Дои:10.1093 / hmg / ddr137. ЧВК 3095054. PMID 21459775.
- ^ а б c d Канасты Р., Доркин-младший, Вегас А., Андерсон Д. (ноябрь 2013 г.). «Материалы для доставки siRNA терапевтических средств». Материалы Природы. 12 (11): 967–77. Bibcode:2013НатМа..12..967K. Дои:10.1038 / nmat3765. PMID 24150415.
- ^ а б c Виттруп А., Либерман Дж. (Сентябрь 2015 г.). «Снятие болезни: отчет о достижениях в области терапии миРНК». Природа Обзоры Генетика. 16 (9): 543–52. Дои:10.1038 / nrg3978. ЧВК 4756474. PMID 26281785.
- ^ Де-Соуза Е.А., Камара Х., Салгейро В.Г., Моро Р.П., Книттель Т.Л., Тонон Дж. И др. (Май 2019). «РНК-интерференция может привести к неожиданным фенотипам у Caenorhabditis elegans». Исследования нуклеиновых кислот. 47 (8): 3957–3969. Дои:10.1093 / нар / gkz154. ЧВК 6486631. PMID 30838421.
- ^ а б c d е ж грамм час я Сюй Ц., Ван Дж. (1 февраля 2015 г.). «Системы доставки для разработки препаратов siRNA в терапии рака». Азиатский журнал фармацевтических наук. 10 (1): 1–12. Дои:10.1016 / j.ajps.2014.08.011.
- ^ а б c d е ж грамм час я j k л м Уайтхед К.А., Дальман Дж. Э., Лангер Р.С., Андерсон Д.Г. (2011). «Молчание или стимуляция доставки siRNA и иммунной системы». Ежегодный обзор химической и биомолекулярной инженерии. 2: 77–96. Дои:10.1146 / annurev-chembioeng-061010-114133. PMID 22432611. S2CID 28803811.
- ^ «FDA пришло к выводу, что арктические яблоки и врожденный картофель безопасны для употребления». Получено 29 сентября 2017.
- ^ Сунилкумар Г., Кэмпбелл Л.М., Пакхабер Л., Стипанович Р.Д., Ратор К.С. (ноябрь 2006 г.). «Разработка семян хлопчатника для использования в питании человека путем тканеспецифического снижения токсичного госсипола». Труды Национальной академии наук Соединенных Штатов Америки. 103 (48): 18054–9. Дои:10.1073 / pnas.0605389103. ЧВК 1838705. PMID 17110445.
- ^ Сиритунга Д., Сайре RT (июль 2003 г.). «Создание трансгенной маниоки, не содержащей цианогена». Planta. 217 (3): 367–73. Дои:10.1007 / s00425-003-1005-8. PMID 14520563. S2CID 13561249.
- ^ Le LQ, Lorenz Y, Scheurer S, Fötisch K, Enrique E, Bartra J, Biemelt S, Vieths S, Sonnewald U (март 2006 г.). «Дизайн плодов томата с пониженной аллергенностью за счет dsRNAi-опосредованного ингибирования экспрессии ns-LTP (Lyc e 3)». Журнал биотехнологии растений. 4 (2): 231–42. Дои:10.1111 / j.1467-7652.2005.00175.x. PMID 17177799.
- ^ Ниггевег Р., Майкл А.Дж., Мартин С. (июнь 2004 г.). «Инженерные заводы с повышенным уровнем антиоксиданта хлорогеновой кислоты». Природа Биотехнологии. 22 (6): 746–54. Дои:10,1038 / nbt966. PMID 15107863. S2CID 21588259.
- ^ Сандерс Р.А., Хиатт В. (март 2005 г.). «Структура трансгена томата и сайленсинг». Природа Биотехнологии. 23 (3): 287–9. Дои:10.1038 / nbt0305-287b. PMID 15765076. S2CID 21191589.
- ^ Chiang CH, Wang JJ, Jan FJ, Yeh SD, Gonsalves D (ноябрь 2001 г.). «Сравнительные реакции рекомбинантных вирусов кольцевой пятнистости папайи с генами химерного белка оболочки (CP) и вирусов дикого типа на CP-трансгенных папайях». Журнал общей вирусологии. 82 (Pt 11): 2827–36. Дои:10.1099/0022-1317-82-11-2827. PMID 11602796. S2CID 25659570.
- ^ Gilbert MK, Majumdar R, Rajasekaran K, Chen ZY, Wei Q, Sickler CM, Lebar MD, Cary JW, Frame BR, Wang K (июнь 2018). «Основанное на РНК-интерференции замалчивание гена альфа-амилазы (amy1) в Aspergillus flavus снижает рост грибов и продукцию афлатоксина в зернах кукурузы». Planta. 247 (6): 1465–1473. Дои:10.1007 / s00425-018-2875-0. PMID 29541880. S2CID 3918937.
- ^ Каточ Р., Такур Н. (март 2013 г.). «РНК-интерференция: перспективный метод улучшения традиционных культур». Международный журнал пищевых наук и питания. 64 (2): 248–59. Дои:10.3109/09637486.2012.713918. PMID 22861122. S2CID 45212581.
- ^ Каточ Р., Такур Н. (март 2013 г.). «Достижения в технологии интерференции РНК и ее влияние на улучшение питания, борьбу с болезнями и насекомыми у растений». Прикладная биохимия и биотехнология. 169 (5): 1579–605. Дои:10.1007 / s12010-012-0046-5. PMID 23322250. S2CID 23733295.
- ^ Гавилано Л. Б., Коулман Н. П., Бернли Л. Е., Боумен М. Л., Каленгамалиро Н. Э., Хейс А., Буш Л., Симински Б. (ноябрь 2006 г.). «Генная инженерия Nicotiana tabacum для снижения содержания норникотина». Журнал сельскохозяйственной и пищевой химии. 54 (24): 9071–8. Дои:10.1021 / jf0610458. PMID 17117792.
- ^ Аллен Р.С., Миллгейт А.Г., Читти Дж. А., Тислтон Дж., Миллер Дж. А., Фист А. Дж., Герлах В. Л., Ларкин П. Дж. (Декабрь 2004 г.). «РНКи-опосредованная замена морфина ненаркотическим алкалоидом ретикулином в опийном маке». Природа Биотехнологии. 22 (12): 1559–66. Дои:10.1038 / nbt1033. PMID 15543134. S2CID 8290821.
- ^ Заде А.Х., Фостер Г.Д. (2004). «Трансгенная устойчивость к вирусу кольцевой пятнистости табака». Acta Virologica. 48 (3): 145–52. PMID 15595207.
- ^ Ивашута С., Чжан И., Виггинс Б.Э., Рамасешадри П., Сегерс Г.К., Джонсон С., Мейер С.Е., Керстеттер Р.А., МакНалти BC, Болоньези Р., Хек Г.Р. (май 2015 г.). «Экологическая РНКи у травоядных насекомых». РНК. 21 (5): 840–50. Дои:10.1261 / rna.048116.114. ЧВК 4408792. PMID 25802407.
- ^ Миллер С.К., Мията К., Браун С.Дж., Томоясу Ю. (2012). «Рассечение системной РНК-интерференции у красного мучного жука Tribolium castaneum: параметры, влияющие на эффективность РНКи». PLOS ONE. 7 (10): e47431. Bibcode:2012PLoSO ... 747431M. Дои:10.1371 / journal.pone.0047431. ЧВК 3484993. PMID 23133513.
- ^ Петрик Дж. С., Фридрих Г. Е., Карлтон С. М., Кессенич К. Р., Сильванович А., Чжан Ю., Кох М. С. (ноябрь 2016 г.). «РНК DvSnf7, активная в отношении корневого червя кукурузы: оценка токсикологии при пероральном приеме многократной дозы в поддержку безопасности человека и млекопитающих». Нормативная токсикология и фармакология. 81: 57–68. Дои:10.1016 / j.yrtph.2016.07.009. PMID 27436086.
- ^ Терениус О., Папаниколау А., Гарбутт Дж. С., Элефтерианос И., Хувенн Х., Кангинакудру С. и др. (Февраль 2011 г.). «РНК-интерференция в Lepidoptera: обзор успешных и неудачных исследований и значение для экспериментального дизайна». Журнал физиологии насекомых. 57 (2): 231–45. Дои:10.1016 / j.jinsphys.2010.11.006. HDL:1854 / LU-1101411. PMID 21078327.
- ^ Khajuria C, Ivashuta S, Wiggins E, Flagel L, Moar W, Pleau M и др. (14 мая 2018 г.). «Разработка и характеристика первой популяции устойчивых к дцРНК насекомых от западного кукурузного корневого червя, Diabrotica virgifera virgifera LeConte». PLOS ONE. 13 (5): e0197059. Bibcode:2018PLoSO..1397059K. Дои:10.1371 / journal.pone.0197059. ЧВК 5951553. PMID 29758046.
- ^ Чжан, Сара (23 июня 2017 г.). «Новая технология генной инженерии Монсанто, одобренная Агентством по охране окружающей среды: впервые РНК-интерференция будет использована для уничтожения насекомых-вредителей». Атлантический океан.
- ^ Матсон Р.С. (2005). Применение технологий геномных и протеомных микрочипов в открытии лекарств. CRC Press. п.6. ISBN 978-0-8493-1469-8.
- ^ Чжан XHD (2011). Оптимальный высокопроизводительный скрининг: практический план экспериментов и анализ данных для исследования РНКи в масштабе генома. Издательство Кембриджского университета. стр. ix – xiii. ISBN 978-0-521-73444-8.
- ^ Мацке М.А., Мацке А.Дж. (2004). «Посадка семян новой парадигмы». ПЛОС Биол. 2 (5): e133. Дои:10.1371 / journal.pbio.0020133. ЧВК 406394. PMID 15138502.
- ^ Эккер-младший, Дэвис Р.В. (август 1986 г.). «Ингибирование экспрессии генов в растительных клетках путем экспрессии антисмысловой РНК». Труды Национальной академии наук Соединенных Штатов Америки. 83 (15): 5372–6. Bibcode:1986PNAS ... 83.5372E. Дои:10.1073 / пнас.83.15.5372. ЧВК 386288. PMID 16593734.
- ^ Napoli C, Lemieux C, Jorgensen R (апрель 1990 г.). «Введение химерного гена халкон-синтазы в петунию приводит к обратимой совместной супрессии гомологичных генов в транс». Растительная клетка. 2 (4): 279–289. Дои:10.1105 / tpc.2.4.279. ЧВК 159885. PMID 12354959.
- ^ Романо Н., Мачино Г. (ноябрь 1992 г.). «Подавление: временная инактивация экспрессии гена в Neurospora crassa путем трансформации гомологичными последовательностями». Молекулярная микробиология. 6 (22): 3343–53. Дои:10.1111 / j.1365-2958.1992.tb02202.x. PMID 1484489. S2CID 31234985.
- ^ Ван Блокланд Р., Ван дер Гест Н., Мол Дж. Н., Кутер Дж. М. (1994). «Опосредованное трансгеном подавление экспрессии халкон-синтазы в Петуния гибридная является результатом увеличения оборота РНК ». Завод J. 6 (6): 861–77. Дои:10.1046 / j.1365-313X.1994.6060861.x.
- ^ Мол Дж. Н., ван дер Крол А. Р. (1991). Антисмысловые нуклеиновые кислоты и белки: основы и приложения. М. Деккер. С. 4, 136. ISBN 978-0-8247-8516-1.
- ^ Кови С., Аль-Кафф Н., Лангара А., Тернер Д. (1997). «Растения борются с инфекцией путем замалчивания генов». Природа. 385 (6619): 781–2. Bibcode:1997Натура.385..781C. Дои:10.1038 / 385781a0. S2CID 43229760.
- ^ Кумагаи М.Х., Донсон Дж., Делла-Чиоппа Дж., Харви Д., Хэнли К., Гриль Л.К. (февраль 1995 г.). «Цитоплазматическое ингибирование биосинтеза каротиноидов вирусной РНК». Труды Национальной академии наук Соединенных Штатов Америки. 92 (5): 1679–83. Bibcode:1995PNAS ... 92.1679K. Дои:10.1073 / pnas.92.5.1679. ЧВК 42583. PMID 7878039.
- ^ Рэтклифф Ф., Харрисон Б.Д., Баулкомб, округ Колумбия (июнь 1997 г.). «Сходство между вирусной защитой и подавлением генов у растений». Наука. 276 (5318): 1558–60. Дои:10.1126 / science.276.5318.1558. PMID 18610513.
- ^ Guo S, Kemphues KJ (май 1995 г.). «par-1, ген, необходимый для установления полярности у эмбрионов C. elegans, кодирует предполагаемую киназу Ser / Thr, которая распределена асимметрично». Клетка. 81 (4): 611–20. Дои:10.1016/0092-8674(95)90082-9. PMID 7758115.
- ^ Пал-Бхадра М., Бхадра Ю., Бирчлер Дж. А. (август 1997 г.). «Косупрессия у Drosophila: подавление гена алкогольдегидрогеназы белыми трансгенами Adh зависит от Polycomb». Клетка. 90 (3): 479–90. Дои:10.1016 / S0092-8674 (00) 80508-5. PMID 9267028.
- ^ Огонь А., Сюй С., Монтгомери М.К., Костас С.А., Драйвер С.Е., Мелло СС (февраль 1998 г.). «Мощное и специфическое генетическое вмешательство двухцепочечной РНК в Caenorhabditis elegans». Природа. 391 (6669): 806–11. Bibcode:1998Натура.391..806F. Дои:10.1038/35888. PMID 9486653. S2CID 4355692.
внешняя ссылка
- Обзор процесса RNAi, из Голые ученые Кембриджского университета
- Анимация процесса RNAi, из Природа
- NOVA scienceNOW объясняет RNAi - 15-минутное видео Новая звезда трансляция, которая вышла в эфир PBS, 26 июля 2005 г.
- Заглушить геномы РНК-интерференция (РНКи) эксперименты и биоинформатика в C. elegans для образования. Из Центра изучения ДНК Долана лаборатории Колд-Спринг-Харбор.
- Скрининг РНКи в C. elegans в 96-луночном жидком формате и их применение для систематической идентификации генетических взаимодействий (протокол)
- 2 американских «Человека-червя» получили Нобелевскую премию за работу в области РНК, из Нью-Йорк Таймс
- Молекулярная терапия веб-фокус: «Разработка РНКи как терапевтической стратегии», сборник бесплатных статей о РНКи как терапевтической стратегии.
- ГеномРНКи: база данных фенотипов из экспериментов по скринингу РНК-интерференции у Drosophila melanogaster и Homo sapians
- Инструменты RNAi Предварительно разработанные и настраиваемые инструменты интерференции РНК