CCR5 - Википедия - CCR5





C-C хемокиновый рецептор 5 типа, также известный как CCR5 или же CD195, это белок на поверхности белые кровяные клетки который участвует в иммунная система поскольку он действует как рецептор за хемокины.[5]
У людей CCR5 ген, кодирующий белок CCR5, является расположен на коротком (p) плече в позиции 21 на хромосома 3. Определенные группы населения унаследовали Дельта 32 мутация, в результате чего генетическая делеция части гена CCR5. Гомозиготный носители этой мутации устойчивы к М-тропные штаммы из ВИЧ-1 инфекция.[6][7][8][9][10][11]
Функция
Белок CCR5 относится к бета-хемокиновые рецепторы семья интегральные мембранные белки.[12][13] Это Рецептор, связанный с G-белком[12] который функционирует как хемокиновый рецептор в CC Chemokine group.
Родственник CCR5 лиганды включают CCL3, CCL4 (также известный как MIP 1α и 1βсоответственно) и CCL3L1.[14][15] CCR5, кроме того, взаимодействует с CCL5 (а хемотаксический цитокин белок, также известный как RANTES).[14][16][17]
CCR5 преимущественно выразил на Т-клетки, макрофаги, дендритные клетки, эозинофилы, микроглия и субпопуляция клеток рака груди или простаты.[18][19] Экспрессия CCR5 избирательно индуцируется во время процесса трансформации рака и не экспрессируется в нормальных эпителиальных клетках молочной железы или предстательной железы. Приблизительно 50% рака груди человека экспрессирует CCR5, в первую очередь при тройном отрицательном раке груди.[18] Ингибиторы CCR5 блокировали миграцию и метастазирование клеток рака груди и простаты, экспрессирующих CCR5, что позволяет предположить, что CCR5 может функционировать как новая терапевтическая мишень.[18][19][20] Недавние исследования показывают, что CCR5 экспрессируется в подгруппе раковых клеток с характеристиками раковых стволовых клеток, которые, как известно, вызывают резистентность к терапии, и что ингибиторы CCR5 увеличивают количество клеток, убитых текущей химиотерапией.[21] Вероятно, что CCR5 играет роль в воспалительных ответах на инфекцию, хотя его точная роль в нормальной иммунной функции неясна. Области этого белка также имеют решающее значение для связывания хемокинового лиганда, функционального ответа рецептора и активности корецептора ВИЧ.[22]

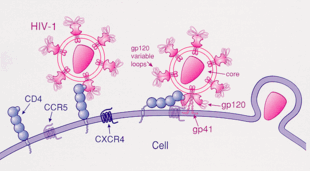
ВИЧ
ВИЧ-1 чаще всего использует хемокиновые рецепторы CCR5 и / или CXCR4 в качестве корецепторы для проникновения в иммунологические клетки-мишени.[23] Эти рецепторы расположены на поверхности иммунных клеток хозяина, посредством чего они обеспечивают способ проникновения вируса ВИЧ-1 для инфицирования клетки.[24] Структура гликопротеина оболочки ВИЧ-1 важна для обеспечения проникновения вируса ВИЧ-1 в целевую клетку-хозяин.[24] Структура гликопротеина оболочки состоит из двух белковых субъединиц, отщепленных от предшественника белка Gp160, кодируемого вирусом ВИЧ-1. env ген: внешняя субъединица Gp120 и трансмембранная субъединица Gp41.[24] Эта структура гликопротеина оболочки расположена в виде шиповидной структуры, расположенной на поверхности вириона, и состоит из тримеров гетеродимеров Gp120-Gp41.[24] Белок оболочки Gp120 является имитатором хемокинов.[23] Хотя ему не хватает уникальной структуры хемокина, он все же способен связываться с рецепторами хемокинов CCR5 и CXCR4.[23] Во время ВИЧ-1-инфекции субъединица гликопротеина оболочки Gp120 связывается с гликопротеином CD4 и корецептором ВИЧ-1, экспрессируемым на клетке-мишени, образуя гетеротримерный комплекс.[23] Образование этого комплекса стимулирует высвобождение фузогенного пептида, заставляя вирусную мембрану сливаться с мембраной целевой клетки-хозяина.[23] Поскольку связывание только с CD4 может иногда приводить к отщеплению gp120, gp120 должен затем связываться с корецептором CCR5, чтобы слияние продолжалось. Тирозин-сульфированный амино-конец этого корецептора является «существенным детерминантом» связывания с гликопротеином gp120.[25] Корецептор также распознает область V1-V2 gp120 и мостиковый лист (антипараллельный четырехцепочечный β-лист, который соединяет внутренний и внешний домены gp120). Стержень V1-V2 может влиять на «использование корецептора через свой пептидный состав, а также степенью N-связанного гликозилирования». Однако, в отличие от V1-V2, петля V3 очень вариабельна и, таким образом, является наиболее важным фактором, определяющим специфичность корецептора.[25] Нормальный лиганды для этого рецептора, RANTES, MIP-1β, и МИП-1α, способны подавлять инфекцию ВИЧ-1 in vitro. У людей, инфицированных ВИЧ, вирусы, использующие CCR5, являются преобладающими видами, выделенными на ранних стадиях вирусной инфекции,[26] предполагая, что эти вирусы могут иметь избирательное преимущество во время передачи или в острой фазе заболевания. Более того, по крайней мере, половина всех инфицированных на протяжении всего периода заражения несет в себе только вирусы, использующие CCR5.
CCR5 - это первичный корецептор, используемый gp120 последовательно с CD4. Это связывание приводит к тому, что gp41, другой белковый продукт gp160, высвобождается из своей метастабильной конформации и внедряется в мембрану клетки-хозяина. Хотя это не было подтверждено, связывание gp120-CCR5 включает два важных этапа: 1) Тирозин-сульфированный амино-конец этого корецептора является «существенной детерминантой» связывания с gp120 (как указано ранее) 2) После этапа 1 ., должно быть реципрокное действие (синергия, взаимосвязь) между gp120 и трансмембранными доменами CCR5.[25]
CCR5 необходим для распространения штамма R5 вируса ВИЧ-1.[27] Знание механизма, с помощью которого этот штамм ВИЧ-1 опосредует инфекцию, побудило исследовать разработку терапевтических вмешательств для блокирования функции CCR5.[28] Ряд новых экспериментальных препаратов от ВИЧ, названных Антагонисты рецепторов CCR5, были разработаны, чтобы препятствовать связыванию между белком оболочки Gp120 и корецептором ВИЧ CCR5.[27] Эти экспериментальные препараты включают PRO140 (CytoDyn ), Викривирок (Испытания фазы III были отменены в июле 2010 г.) (Schering Plough ), Аплавирок (GW-873140) (GlaxoSmithKline ) и Маравирок (UK-427857) (Pfizer ). Маравирок был одобрен для использования FDA в августе 2007 года.[27] На сегодняшний день это единственный препарат, одобренный FDA для клинического использования, и, таким образом, он стал первым ингибитором CCR5.[25] Проблема этого подхода состоит в том, что, хотя CCR5 является основным корецептором, с помощью которого ВИЧ заражает клетки, он не единственный такой корецептор. Возможно, что под давлением отбора ВИЧ эволюционирует и использует другой корецептор. Однако исследование вирусной устойчивости к AD101, молекулярной антагонист CCR5, показали, что устойчивые вирусы не переключаются на другой корецептор (CXCR4), но продолжают использовать CCR5: они либо связываются с альтернативными доменами CCR5, либо с рецептором с более высоким сродством. Однако, поскольку есть еще один доступный корецептор, вероятно, что отсутствие гена CCR5 не делает человека невосприимчивым к вирусу; для человека было бы просто сложнее заразиться этим. Также у вируса все еще есть доступ к CD4. В отличие от CCR5, который не требуется (о чем свидетельствуют люди, ведущие здоровый образ жизни даже при отсутствии гена в результате мутации delta32), CD4 имеет решающее значение для системы иммунной защиты организма.[29] Даже при отсутствии корецептора (даже CCR5) вирус все еще может проникать в клетки, если gp41 претерпит изменения (включая его цитоплазматический хвост), которые привели к независимости CD4 без необходимости CCR5 и / или CXCR4. как дверной проем.[30]
Рак
Экспрессия CCR5 индуцируется в эпителиальных клетках груди и простаты после трансформации. Индукция экспрессии CCR5 способствует клеточной инвазии, миграции и метастазированию. Для индукции метастазирования необходимо вернуться к месту метастазирования. Было показано, что ингибиторы CCR5, включая маравирок и леронлимаб, блокируют метастазирование в легкие клеточных линий рака молочной железы человека.[18] В доклинических исследованиях иммунокомпетентных мышей ингибиторы CCR5 блокировали метастазирование в кости и мозг.[19] Ингибиторы CCR5 также уменьшают инфильтрацию ассоциированных с опухолью макрофагов.[31] Фаза 1 клинического исследования ингибитора CCR5 у пациентов с метастатическим раком толстой кишки, подвергавшихся интенсивному предварительному лечению, продемонстрировала объективный клинический ответ и снижение метастатической опухолевой нагрузки.[32]
Мозг
Повышенные уровни CCR5 являются частью воспалительной реакции на инсульт. Блокирование CCR5 с помощью маравирока (препарата, одобренного для лечения ВИЧ) может ускорить восстановление после инсульта.[33][34]
В развивающемся мозге хемокины, такие как CCR5, влияют на миграцию и связь нейронов. После инсульта они, кажется, уменьшают количество участков соединения на нейронах вблизи места повреждения.[33]
CCR5-Δ32
CCR5-Δ32 (или CCR5-D32 или CCR5 delta 32) - это аллель из CCR5.[35][36]
CCR5 Δ32 представляет собой делецию из 32 пар оснований, которая приводит к преждевременной стоп-кодон в локус рецептора CCR5, в результате чего рецептор становится нефункциональным.[37][38] CCR5 необходим для проникновения M-тропного вируса ВИЧ-1.[39] Физическим лицам гомозиготный (обозначены Δ32 / Δ32) для CCR5 Δ32 не экспрессируют функциональные рецепторы CCR5 на своей клеточной поверхности и устойчивы к ВИЧ-1 инфекция, несмотря на многократные воздействия высокого риска.[39] Физическим лицам гетерозиготный (+ / Δ32) для мутанта аллель имеют более чем 50% снижение функциональных рецепторов CCR5 на их клеточных поверхностях из-за димеризации между мутантными рецепторами и рецепторами дикого типа, что препятствует транспорту CCR5 к поверхности клетки.[40] Гетерозиготы-носители устойчивы к ВИЧ-1 инфекции по сравнению с дикими типами, а при инфицировании гетерозиготы проявляют пониженную вирусные нагрузки и на 2-3 года медленнее СПИД относительно диких видов.[37][39][41] Также было показано, что гетерозиготность по этому мутантному аллелю улучшает вирусологический ответ на антиретровирусное лечение.[42] CCR5 Δ32 имеет частоту (гетерозиготного) аллеля 10% в Европе и частоту гомозигот 1%.
Недавние исследования показывают, что CCR5 Δ32 улучшает познавательные способности и память. В 2016 году исследователи показали, что удаление гена CCR5 у мышей значительно улучшило их память.[43] CCR5 является мощным супрессором нейрональной пластичности, обучения и памяти; Чрезмерная активация CCR5 вирусными белками может способствовать когнитивному дефициту, связанному с ВИЧ.[44]
Эволюционная история и возраст аллеля
Аллель CCR5 Δ32 отличается недавним происхождением, неожиданно высокой частотой и четким географическим распределением.[45] которые вместе предполагают, что (а) он возник в результате единственной мутации и (б) исторически подвергался положительному отбору.
Два исследования использовали анализ сцепления для оценки возраста делеции CCR5 Δ32, предполагая, что количество рекомбинации и мутации, наблюдаемых в геномных областях, окружающих делецию CCR5 Δ32, будет пропорционально возрасту делеции.[36][46] Используя выборку из 4000 человек из 38 этнических групп, Stephens et al. подсчитали, что делеция CCR5-Δ32 произошла 700 лет назад (275-1875, 95% доверительный интервал). Другая группа, Libert et al. (1998) использовали микросателлитные мутации для оценки возраста мутации CCR5 Δ32 как 2100 лет (700–4800, 95% доверительный интервал). На основе наблюдаемых событий рекомбинации они оценили возраст мутации в 2250 лет (900-4700, 95% доверительный интервал).[46] Третья гипотеза опирается на градиент частоты аллелей с севера на юг в Европе, который показывает, что самая высокая частота аллелей имела место в Скандинавские страны и самая низкая частота аллелей в южной Европе. Поскольку викинги исторически населяли эти страны, возможно, что аллель распространился по Европе из-за расселения викингов в 8-10 веках.[47] Позже викинги были заменены Варяги в России, что, возможно, способствовало наблюдаемому изменению частоты аллелей с востока на запад.[45][47]
Первоначально ВИЧ-1 передавался от шимпанзе (Пан троглодиты) людям в начале 1900-х годов в Юго-Восточном Камеруне, Африка,[48] через контакт с инфицированной кровью и биологическими жидкостями при разделке мяса диких животных.[49] Однако ВИЧ-1 фактически отсутствовал в Европе до 1980-х годов.[50] Следовательно, учитывая средний возраст примерно 1000 лет для аллеля CCR5-Δ32, можно установить, что ВИЧ-1 не оказывал давления отбора на человеческую популяцию достаточно долго, чтобы достичь нынешних частот.[45] Следовательно, другие патогены были предложены в качестве агентов положительного отбора для CCR5 Δ32, включая бубонная чума (Yersinia pestis ) и оспа (Вариола основной Другие данные предполагают, что частота аллелей испытывала отрицательное давление отбора в результате патогенов, которые стали более распространенными во время римской экспансии.[51] Идея о том, что отрицательный отбор играет роль в низкой частоте аллеля, также подтверждается экспериментами с использованием мышей с нокаутом и вирусом гриппа A, которые продемонстрировали, что присутствие рецептора CCR5 важно для эффективного ответа на патоген.[52][53]
Доказательства единственной мутации
Несколько линий доказательств предполагают, что аллель Δ32 CCR5 эволюционировал только один раз.[45] Во-первых, CCR5 Δ32 имеет относительно высокую частоту в нескольких различных европейских популяциях, но сравнительно отсутствует в популяциях азиатских, ближневосточных и американских индейцев,[36] предполагая, что единственная мутация произошла после расхождения европейцев от их африканского предка.[36][37][54] Во-вторых, анализ генетического сцепления показывает, что мутация происходит на гомогенном генетическом фоне, подразумевая, что мутация унаследована от общего предка.[46] Это было продемонстрировано, показывая, что аллель CCR5 Δ32 находится в сильном неравновесном сцеплении с высокополиморфными микросателлитами. Более 95% хромосом CCR5 Δ32 также несли аллель IRI3.1-0, а 88% несли аллель IRI3.2. Напротив, микросателлитные маркеры IRI3.1-0 и IRI3.2-0 были обнаружены только в 2 или 1,5% хромосом, несущих аллель CCR5 дикого типа.[46] Это свидетельство неравновесия по сцеплению поддерживает гипотезу о том, что большинство, если не все, аллели CCR5 Δ32 возникли в результате одного мутационного события. Наконец, аллель CCR5 Δ32 имеет уникальное географическое распределение, указывающее на единственное северное происхождение с последующей миграцией. Исследование, измеряющее частоты аллелей в 18 европейских популяциях, обнаружило градиент с севера на юг с самыми высокими частотами аллелей в финских и Мордовский населения (16%), а самый низкий на Сардинии (4%).[46]
Положительный выбор
В отсутствие отбора одной мутации потребуется примерно 127 500 лет, чтобы она выросла до 10% популяционной частоты.[36] По оценкам, основанным на генетической рекомбинации и частоте мутаций, возраст аллеля составляет от 1000 до 2000 лет. Это несоответствие является признаком положительного отбора.
Подсчитано, что ВИЧ-1 проник в популяцию людей в Африке в начале 1900-х годов;[48] однако о симптоматических инфекциях не сообщалось до 1980-х годов. Следовательно, эпидемия ВИЧ-1 еще слишком молода, чтобы быть источником положительного отбора, который привел к тому, что частота CCR5 Δ32 с нуля до 10% за 2000 лет. Стивенс и др. (1998), предполагают, что бубонная чума (Yersinia pestis) оказал положительное селективное давление на CCR5 Δ32.[36] Эта гипотеза была основана на сроках и серьезности пандемии черной смерти, в результате которой погибло 30% европейского населения всех возрастов между 1346 и 1352 годами.[55] После Черной смерти были менее серьезные периодические эпидемии. В отдельных городах наблюдалась высокая смертность, но общая смертность в Европе составляла всего несколько процентов.[55][56][57] В 1655–1656 годах вторая пандемия, получившая название «Великая чума», унесла жизни 15-20% населения Европы.[55][58][сомнительный ] Важно отметить, что эпидемии чумы носили прерывистый характер. Бубонная чума - это зоонозное заболевание, поражающее в первую очередь грызунов, распространяемое блохами и лишь изредка заражающее людей.[59] Передача от человека человеку бубонной чумы не происходит, хотя может произойти при легочной чуме, поражающей легкие.[60] Только при низкой плотности грызунов инфицированные блохи вынуждены питаться альтернативными хозяевами, такими как люди, и в этих обстоятельствах может возникнуть человеческая эпидемия.[59] Основываясь на популяционно-генетических моделях, Гальвани и Слаткин (2003) утверждают, что прерывистый характер эпидемий чумы не создавал достаточно сильной селективной силы, чтобы довести частоту аллелей CCR5 Δ32 до 10% в Европе.[35]
Чтобы проверить эту гипотезу, Гальвани и Слаткин (2003) смоделировали историческое давление отбора, вызванное чумой и оспой.[35] Чума была смоделирована в соответствии с историческими данными,[61][62] в то время как возрастная смертность от оспы была определена по возрастному распределению оспенных захоронений в Йорке (Англия) между 1770 и 1812 годами.[56] Оспа преимущественно поражает молодых представителей населения до репродуктивного возраста, поскольку они - единственные люди, которые не были иммунизированы или умерли от перенесенной инфекции. Поскольку оспа преимущественно убивает представителей популяции до репродуктивного возраста, она создает более сильное избирательное давление, чем чума.[35] В отличие от чумы, оспа не имеет животного резервуара и передается только от человека к человеку.[63][64] Авторы подсчитали, что если бы чума отбирала CCR5 Δ32, частота аллеля все равно была бы менее 1%, в то время как оспа проявляла силу отбора, достаточную для достижения 10%.
Гипотеза о том, что натуральная оспа оказывала положительный отбор на CCR5 Δ32, также является биологически правдоподобной, поскольку поксвирусы, такие как ВИЧ, проникают в лейкоциты с помощью хемокиновых рецепторов.[65] Напротив, Yersinia pestis это бактерия с совершенно другой биологией.
Хотя европейцы являются единственной группой, в которой есть субпопуляции с высокой частотой CCR5 Δ32, они не единственная популяция, которая была подвергнута селекции оспой, которая имела всемирное распространение до того, как была объявлена искорененной в 1980 году. Самые ранние безошибочные описания оспы. оспа появилась в 5 веке нашей эры в Китае, в 7 веке нашей эры в Индии и Средиземноморье и в 10 веке нашей эры в Юго-Западной Азии.[64] Напротив, CCR5 Δ32 мутация встречается только в популяциях Европы, Западной Азии и Северной Африки.[66] Аномально высокая частота CCR5 Δ32 в этих популяциях, по-видимому, требует как уникального происхождения в Северной Европе, так и последующей селекции оспой.
Возможные затраты
CCR5 Δ32 может быть полезным для хозяина при некоторых инфекциях (например, ВИЧ-1, возможно, оспой), но пагубным при других (например, клещевом энцефалите, вирусе Западного Нила). Будет ли функция CCR5 полезна или вредна в контексте данной инфекции, зависит от сложного взаимодействия между иммунной системой и патогеном.
В целом, исследования показывают, что мутация CCR5 Δ32 может играть пагубную роль в постинфекционных воспалительных процессах, которые могут повредить ткань и создать дополнительную патологию.[67] Лучшее доказательство этой предполагаемой антагонистической плейотропии найдено в флавивирус инфекции. Как правило, многие вирусные инфекции протекают бессимптомно или вызывают лишь легкие симптомы у подавляющего большинства населения. Однако у некоторых незадачливых людей протекает особенно разрушительное клиническое течение, которое иначе необъяснимо, но, по-видимому, является генетически опосредованным. Было обнаружено, что пациенты, гомозиготные по CCR5 Δ32, имеют более высокий риск нейроинвазивной формы клещевого энцефалита (флавивирус).[68] Кроме того, функциональный CCR5 может потребоваться для предотвращения симптоматического заболевания после инфицирования вирус Западного Нила, другой флавивирус; CCR5 Δ32 был связан с ранним развитием симптомов и более выраженными клиническими проявлениями после заражения вирусом Западного Нила.[69]
Это открытие на людях подтвердило ранее наблюдаемый эксперимент на животной модели гомозиготности CCR5 Δ32. После заражения вирусом Западного Нила у мышей CCR5 Δ32 заметно увеличились титры вируса в центральной нервной системе и повысилась смертность.[70] по сравнению с таковой у мышей дикого типа, что позволяет предположить, что экспрессия CCR5 необходима для создания сильной защиты хозяина от вируса Западного Нила.
Медицинские приложения
Генетический подход, включающий интратела, которые блокируют экспрессию CCR5, был предложен для лечения ВИЧ-1 инфицированные лица.[71] Когда Т-клетки модифицировались таким образом, что они больше не экспрессируют CCR5, были смешаны с немодифицированными Т-клетками, экспрессирующими CCR5, а затем заражены ВИЧ-1, модифицированные Т-клетки, которые не экспрессируют CCR5, в конечном итоге захватили культуру, как ВИЧ-1. убивает немодифицированные Т-клетки. Этот же метод можно использовать in vivo для создания пула устойчивых к вирусам клеток у инфицированных людей.[71]
Эта гипотеза была проверена на больном СПИДом, у которого также развился миелоидный лейкоз, и лечился химиотерапия чтобы подавить рак. А пересадка костного мозга содержащий стволовые клетки от подходящего донора затем использовали для восстановления иммунной системы. Однако трансплантат был выполнен от донора с 2 копиями гена мутации CCR5-Δ32. Через 600 дней пациент был здоров и имел неопределяемые уровни ВИЧ в крови и исследованных тканях мозга и прямой кишки.[7][72] Перед трансплантацией низкий уровень ВИЧ X4, который не использует рецептор CCR5, также были обнаружены. Однако после трансплантации этот тип ВИЧ также не был обнаружен.[7] Однако этот результат согласуется с наблюдением, что клетки, экспрессирующие вариантный белок CCR5-Δ32, лишены как CCR5, так и CXCR4 рецепторов на их поверхности, тем самым обеспечивая устойчивость к широкому спектру вариантов ВИЧ, включая ВИЧX4.[73] По прошествии более шести лет у пациента сохранилась устойчивость к ВИЧ, и было объявлено, что он излечился от ВИЧ-инфекции.[8]
Набор ВИЧ-положительных пациентов в клиническое испытание было начато в 2009 году, в котором клетки пациентов были генетически модифицированы с помощью нуклеаза цинкового пальца чтобы нести черту CCR5-Δ32, а затем повторно ввести в организм в качестве потенциального средства лечения ВИЧ.[74][75] Результаты, представленные в 2014 году, были многообещающими.[11]
Вдохновленный первым человеком, излечившимся от ВИЧ, Берлинский пациент, StemCyte начала сотрудничество с Банки пуповинной крови по всему миру систематически проверять Пуповинная кровь образцы на мутацию CCR5 начиная с 2011 г.[76][77][78]
В ноябре 2018 г. Цзянькуй Хэ объявил, что отредактировал два человеческих эмбриона, чтобы попытаться отключить ген CCR5, который кодирует рецептор, ВИЧ используется для входа в клетки. Он сказал, что девочки-близнецы, Лулу и Нана, родились несколькими неделями ранее, и что у девочек все еще есть функциональные копии CCR5 вместе с отключенным CCR5 (мозаика ), следовательно, он все еще уязвим для ВИЧ. Работа была широко осуждена как неэтичная, опасная и преждевременная.[79][80]
Смотрите также
- Открытие и разработка антагонистов рецептора CCR5
- Ингибитор входа
- ВИЧ-тропизм
- Стивен Крон
- Иммунитет к ВИЧ
Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000160791 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000079227 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «Ссылка на Mouse PubMed:». Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Цзяо X, Nawab O, Patel T, Kossenkov AV, Halama N, Jaeger D, Pestell RG (октябрь 2019 г.). «Последние достижения, направленные на лечение рака CCR5 и его роль в иммуноонкологии». Исследования рака. 79 (19): 4801–4807. Дои:10.1158 / 0008-5472.CAN-19-1167. ЧВК 6810651. PMID 31292161.
- ^ де Сильва, член парламента Штумпфа (декабрь 2004 г.). «ВИЧ и аллель устойчивости к CCR5-Delta32». Письма о микробиологии FEMS. 241 (1): 1–12. Дои:10.1016 / j.femsle.2004.09.040. PMID 15556703.
- ^ а б c Hütter G, Nowak D, Mossner M, Ganepola S, Müssig A, Allers K, Schneider T, Hofmann J, Kücherer C, Blau O, Blau IW, Hofmann WK, Thiel E (февраль 2009 г.). «Долгосрочный контроль ВИЧ с помощью трансплантации стволовых клеток CCR5 Delta32 / Delta32». Медицинский журнал Новой Англии. 360 (7): 692–8. Дои:10.1056 / NEJMoa0802905. PMID 19213682.
- ^ а б Аллерс К., Хюттер Дж., Хофманн Дж., Лодденкемпер С., Ригер К., Тиль Э., Шнайдер Т. (март 2011 г.). «Доказательства лечения ВИЧ-инфекции с помощью трансплантации стволовых клеток CCR5Δ32 / Δ32». Кровь. 117 (10): 2791–9. Дои:10.1182 / blood-2010-09-309591. PMID 21148083.
- ^ Чжэнь А., Кухня S (январь 2014 г.). «Генная терапия на основе стволовых клеток при ВИЧ-инфекции». Вирусы. 6 (1): 1–12. Дои:10.3390 / v6010001. ЧВК 3917429. PMID 24368413.
- ^ Кей М.А., Уокер Б.Д. (март 2014 г.). «Инженерия клеточной устойчивости к ВИЧ». Медицинский журнал Новой Англии. 370 (10): 968–9. Дои:10.1056 / NEJMe1400593. PMID 24597871.
- ^ а б Тебас П., Штейн Д., Тан В.В., Фрэнк И., Ван С.К., Ли Джи и др. (Март 2014 г.). «Редактирование генов CCR5 в аутологичных Т-лимфоцитах CD4 людей, инфицированных ВИЧ». Медицинский журнал Новой Англии. 370 (10): 901–10. Дои:10.1056 / NEJMoa1300662. ЧВК 4084652. PMID 24597865.
- ^ а б «CCR5 - хемокиновый (мотив C-C) рецептор 5 (ген / псевдоген)». Домашний справочник по генетике.
- ^ Самсон М., Лаббе О., Моллеро С., Вассар Г., Парментье М. (март 1996 г.). «Молекулярное клонирование и функциональная экспрессия нового гена рецептора CC-хемокина человека». Биохимия. 35 (11): 3362–7. Дои:10.1021 / bi952950g. PMID 8639485.
- ^ а б Struyf S, Menten P, Lenaerts JP, Put W, D'Haese A, De Clercq E, Schols D, Proost P, Van Damme J (июль 2001 г.). «Различная связывающая способность природных изоформ LD78beta макрофагального воспалительного протеина-1альфа с хемокиновыми рецепторами CC 1, 3 и 5 влияет на их активность против ВИЧ-1 и хемотаксические способности нейтрофилов и эозинофилов». Европейский журнал иммунологии. 31 (7): 2170–8. Дои:10.1002 / 1521-4141 (200107) 31: 7 <2170 :: AID-IMMU2170> 3.0.CO; 2-D. PMID 11449371.
- ^ Миякава Т., Обару К., Маеда К., Харада С., Мицуя Х (февраль 2002 г.). «Идентификация аминокислотных остатков, критических для LD78beta, варианта макрофагального воспалительного белка-1альфа человека, связывания с CCR5 и ингибирования репликации вируса иммунодефицита человека типа 1 R5». Журнал биологической химии. 277 (7): 4649–55. Дои:10.1074 / jbc.M109198200. PMID 11734558.
- ^ Slimani H, Charnaux N, Mbemba E, Saffar L, Vassy R, Vita C, Gattegno L (октябрь 2003 г.). «Взаимодействие RANTES с синдеканом-1 и синдеканом-4, экспрессируемым первичными макрофагами человека». Biochimica et Biophysica Acta (BBA) - Биомембраны. 1617 (1–2): 80–8. Дои:10.1016 / j.bbamem.2003.09.006. PMID 14637022.
- ^ Proudfoot AE, Fritchley S, Borlat F, Shaw JP, Vilbois F, Zwahlen C, Trkola A, Marchant D, Clapham PR, Wells TN (апрель 2001 г.). «Мотив BBXB в RANTES является основным сайтом связывания гепарина и контролирует селективность рецептора». Журнал биологической химии. 276 (14): 10620–6. Дои:10.1074 / jbc.M010867200. PMID 11116158.
- ^ а б c d Веласко-Веласкес М., Цзяо X, Де Ла Фуэнте М., Пестелл Т.Г., Эртель А., Лисанти М.П., Пестелл Р.Г. (август 2012 г.). «Антагонист CCR5 блокирует метастазирование базальных клеток рака молочной железы». Исследования рака. 72 (15): 3839–50. Дои:10.1158 / 0008-5472.CAN-11-3917. PMID 22637726.
- ^ а б c Sicoli D, Jiao X, Ju X, Velasco-Velazquez M, Ertel A, Addya S, Li Z, Andò S, Fatatis A, Paudyal B, Cristofanilli M, Thakur ML, Lisanti MP, Pestell RG (декабрь 2014 г.). «Антагонисты рецептора CCR5 блокируют метастазирование в кость линий метастатических клеток рака предстательной железы, трансформированных онкогеном v-Src». Исследования рака. 74 (23): 7103–14. Дои:10.1158 / 0008-5472.CAN-14-0612. ЧВК 4294544. PMID 25452256.
- ^ Веласко-Веласкес М., Ксолальпа В., Пестелл Р.Г. (ноябрь 2014 г.). «CCL5 / CCR5 при раке груди». Экспертное мнение Ther Target. 18 (11): 1265–75. Дои:10.1517/14728222.2014.949238. PMID 25256399. S2CID 7976259.
- ^ Цзяо Х, Веласко-Веласкес М.А., Ван М., Ли З., Руи Х., Пек А.Р., Коркола Дж. Э., Чен Х, Сюй С., Ду Хадавей Дж. Б., Герреро-Родригес С., Аддя С., Сиколи Д., Му З., Чжан Дж., Стаки A, Zhang X, Cristofanilli M, Fatatis A, Gray JW, Zhong JF, Prendergast GC, Pestell RG (апрель 2018 г.). «CCR5 управляет восстановлением повреждений ДНК и увеличением стволовых клеток рака груди». Рак Res. 78 (7): 1657–71. Дои:10.1158 / 0008-5472.CAN-17-0915. ЧВК 6331183. PMID 29358169.
- ^ Бармания Ф, Пеппер М.С. (2013). «C-C хемокиновый рецептор пятого типа (CCR5): новая мишень для борьбы с ВИЧ-инфекцией». Прикладная и трансляционная геномика. 2 (а): 3–16. Дои:10.1016 / j.atg.2013.05.004. ЧВК 5133339. PMID 27942440.
- ^ а б c d е Мерфи PM (февраль 2001 г.). «Вирусная эксплуатация и подрыв иммунной системы посредством мимикрии хемокинов». Иммунология природы. 2 (2): 116–22. Дои:10.1038/84214. PMID 11175803. S2CID 29364407.
- ^ а б c d Алхатиб Г (март 2009 г.). «Биология CCR5 и CXCR4». Текущее мнение о ВИЧ и СПИДе. 4 (2): 96–103. Дои:10.1097 / COH.0b013e328324bbec. ЧВК 2718543. PMID 19339947.
- ^ а б c d «Ингибиторы CCR5 и ВИЧ».
- ^ Андерсон Дж., Аккина Р. (сентябрь 2007 г.). «Полный нокдаун CCR5 siRNA, экспрессируемые лентивирусным вектором, и защита трансгенных макрофагов от ВИЧ-1 инфекции». Генная терапия. 14 (17): 1287–97. Дои:10.1038 / sj.gt.3302958. PMID 17597795.
- ^ а б c Либерман-Блюм SS, Fung HB, Bandres JC (июль 2008 г.). «Маравирок: антагонист рецептора CCR5 для лечения ВИЧ-1 инфекции». Клиническая терапия. 30 (7): 1228–50. Дои:10.1016 / s0149-2918 (08) 80048-3. PMID 18691983.
- ^ Назари Р., Джоши С. (август 2008 г.). «CCR5 как мишень для генной терапии ВИЧ-1». Современная генная терапия. 8 (4): 264–72. Дои:10.2174/156652308785160674. PMID 18691022.
- ^ "Понимание генетики". Технический музей инноваций, Сан-Хосе, Калифорния.
- ^ Taylor BM, Foulke JS, Flinko R, Heredia A, DeVico A, Reitz M (июнь 2008 г.). «Изменение gp41 вируса иммунодефицита человека приводит к снижению зависимости CCR5 и независимости от CD4». Журнал вирусологии. 82 (11): 5460–71. Дои:10.1128 / JVI.01049-07. ЧВК 2395218. PMID 18353949.
- ^ Франкенбергер С., Рабе Д., Байнер Р., Шанкарашарма Д., Чада К., Краус Т., Гилад И., Беккер Л., Роснер М. Р. (октябрь 2015 г.). «Подавители метастазов регулируют микросреду опухоли путем блокирования рекрутирования прометастатических макрофагов, связанных с опухолью». Рак Res. 75 (19): 4063–73. Дои:10.1158 / 0008-5472.CAN-14-3394. ЧВК 4592465. PMID 26238785.
- ^ Halama N, Zoernig I, Berthel A, Kahlert C, Klupp F, Suarez-Carmona M, Suetterlin T, Brand K, Krauss J, Lasitschka F, Lerchl T, Luckner-Minden C, Ulrich A, Koch M, Weitz J, Schneider M, Buechler MW, Zitvogel L, Herrmann T, Benner A, Kunz C, Luecke S, Springfeld C, Grabe N, Falk CS, Jaeger D (апрель 2016 г.). «Использование опухолевых иммунных клеток при метастазах колоректального рака может быть эффективно нацелено с помощью терапии против CCR5 у онкологических больных». Раковая клетка. 29 (4): 587–601. Дои:10.1016 / j.ccell.2016.03.005. PMID 27070705.
- ^ а б «Препарат от ВИЧ может улучшить восстановление после инсульта». ScienceMag. 2019-02-21. Получено 2019-02-22.
- ^ Joy MT, Assayag EB, Shabashov-Stone D, Liraz-Zaltsman S, Mazzitelli J, Arenas M, Abduljawad N, Kliper E, Korczyn AD, Thareja NS, Kesner EL, Zhou M, Huang S, Silva T.K, Katz N, Bornstein НМ, Сильва А.Дж., Шохами Э., Кармайкл СТ (21.02.2019). «CCR5 - терапевтическая мишень для восстановления после инсульта и травмы головного мозга». Клетка. 176 (5): 1143–1157. Дои:10.1016 / j.cell.2019.01.044. ЧВК 7259116. PMID 30794775.
- ^ а б c d Гальвани А.П., Слаткин М. (декабрь 2003 г.). «Оценка чумы и оспы как исторических факторов селективного воздействия на аллель устойчивости к ВИЧ CCR5-Delta 32». Труды Национальной академии наук Соединенных Штатов Америки. 100 (25): 15276–9. Bibcode:2003ПНАС..10015276Г. Дои:10.1073 / pnas.2435085100. ЧВК 299980. PMID 14645720.
- ^ а б c d е ж Stephens JC, Reich DE, Goldstein DB, Shin HD, Smith MW, Carrington M и др. (Июнь 1998 г.). «Определение происхождения аллеля устойчивости к СПИДу CCR5-Delta32 по слиянию гаплотипов». Американский журнал генетики человека. 62 (6): 1507–15. Дои:10.1086/301867. ЧВК 1377146. PMID 9585595.
- ^ а б c Дин М., Кэррингтон М., Винклер С., Хаттли Г.А., Смит М.В., Алликметс Р. и др. (Сентябрь 1996 г.). «Генетическое ограничение инфекции ВИЧ-1 и прогрессирование до СПИДа за счет делеционного аллеля структурного гена CKR5. Исследование роста и развития гемофилии, многоцентровое когортное исследование СПИДа, многоцентровое когортное исследование гемофилии, городская когорта Сан-Франциско, исследование ALIVE». Наука. 273 (5283): 1856–62. Bibcode:1996Научный ... 273.1856D. Дои:10.1126 / science.273.5283.1856. PMID 8791590. S2CID 21160580.
- ^ Samson M, Libert F, Doranz BJ, Rucker J, Liesnard C, Farber CM, Saragosti S, Lapoumeroulie C, Cognaux J, Forceille C, Muyldermans G, Verhofstede C, Burtonboy G, Georges M, Imai T, Rana S, Yi Y , Смит Р.Дж., Коллман Р.Г., Домс Р.В., Вассарт Дж., Парментье М. (август 1996 г.). «Устойчивость к ВИЧ-1 инфекции у людей европеоидной расы, несущих мутантные аллели гена хемокинового рецептора CCR-5». Природа. 382 (6593): 722–5. Bibcode:1996Натура.382..722S. Дои:10.1038 / 382722a0. PMID 8751444. S2CID 4370818.
- ^ а б c Лю Р., Пакстон В.А., Чоу С., Серадини Д., Мартин С.Р., Хорук Р., Макдональд М.Э., Стулманн Х., Куп Р.А., Ландау Н.Р. (август 1996 г.). «Гомозиготный дефект корецептора ВИЧ-1 является причиной устойчивости некоторых многократно подвергавшихся воздействию людей к инфекции ВИЧ-1». Клетка. 86 (3): 367–77. Дои:10.1016 / s0092-8674 (00) 80110-5. PMID 8756719. S2CID 16965624.
- ^ Бенкиран М., Джин Д.Й., Чун Р.Ф., Куп Р.А., Джанг К.Т. (декабрь 1997 г.). «Механизм трансдоминантного ингибирования CCR5-опосредованной инфекции ВИЧ-1 с помощью ccr5delta32». Журнал биологической химии. 272 (49): 30603–6. Дои:10.1074 / jbc.272.49.30603. PMID 9388191.
- ^ Майкл Н.Л., Луи Л.Г., Рорбо А.Л., Шульц К.А., Дайхофф Д.Е., Ван С.Е., Шеппард Х.В. (октябрь 1997 г.). «Роль полиморфизмов CCR5 и CCR2 в передаче ВИЧ-1 и прогрессировании заболевания». Природа Медицина. 3 (10): 1160–2. Дои:10,1038 / нм 1097-1160. PMID 9334732. S2CID 23196048.
- ^ Laurichesse JJ, Persoz A, Theodorou I, Rouzioux C, Delfraissy JF, Meyer L (май 2007 г.). «Улучшенный вирусологический ответ на высокоактивную антиретровирусную терапию у ВИЧ-1-инфицированных пациентов с делецией CCR5 Delta32». ВИЧ Медицина. 8 (4): 213–9. Дои:10.1111 / j.1468-1293.2007.00455.x. PMID 17461848. S2CID 24900064.
- ^ «У китайских близнецов CRISPR, возможно, был непреднамеренно улучшен мозг». Обзор технологий. Получено 2019-02-22.
- ^ Чжоу М., Гринхилл С., Хуанг С., Сильва Т.К., Сано И., Ву С., Цай Ю., Нагаока Ю., Сегал М., Цай Диджей, Ли Ю.С., Фокс К., Сильва А.Дж. (2016-12-20). «CCR5 является супрессором корковой пластичности, обучения и памяти гиппокампа». eLife. 5. Дои:10.7554 / eLife.20985. ЧВК 5213777. PMID 27996938.
- ^ а б c d Galvani AP, Novembre J (февраль 2005 г.). «Эволюционная история мутации устойчивости к ВИЧ CCR5-Delta32». Микробы и инфекции / Institut Pasteur. 7 (2): 302–9. Дои:10.1016 / j.micinf.2004.12.006. PMID 15715976.
- ^ а б c d е Либерт Ф., Кошо П., Бекман Дж., Самсон М., Аксенова М., Цао А. и др. (Март 1998 г.). «Мутация deltaccr5, обеспечивающая защиту от ВИЧ-1 в кавказских популяциях, имеет единственное и недавнее происхождение в Северо-Восточной Европе». Молекулярная генетика человека. 7 (3): 399–406. Дои:10,1093 / чмг / 7,3,399. PMID 9466996.
- ^ а б Люкотт Дж. (2001). «Распространение делеции 32 пары оснований гена CCR5 в Западной Европе. Гипотеза о возможном распространении мутации викингами в исторические времена». Иммунология человека. 62 (9): 933–6. Дои:10.1016 / S0198-8859 (01) 00292-0. PMID 11543895.
- ^ а б Кил Б.Ф., Ван Хеуверсвин Ф., Ли И, Бейлес Э, Такехиса Дж., Сантьяго М.Л., Биболлет-Руш Ф, Чен И, Уэйн Л.В., Льежуа Ф, Лоул С., Нголе Э.М., Бьенвеню Y, Делапорте Э, Брукфилд Дж. Ф., Шарп П.М. , Shaw GM, Peeters M, Hahn BH (июль 2006 г.). «Шимпанзе - резервуары пандемического и непандемического ВИЧ-1». Наука. 313 (5786): 523–6. Bibcode:2006Научная ... 313..523K. Дои:10.1126 / science.1126531. ЧВК 2442710. PMID 16728595.
- ^ Хан Б. Х., Шоу GM, Де Кок К. М., Sharp PM (январь 2000 г.). «СПИД как зооноз: последствия для науки и здравоохранения». Наука. 287 (5453): 607–14. Bibcode:2000Sci ... 287..607H. Дои:10.1126 / science.287.5453.607. PMID 10649986.
- ^ Матич С. «ВИЧ / СПИД в Европе: от смертного приговора к лечению хронических заболеваний» (PDF). Всемирная организация здоровья.
- ^ Фор Э, Ройер-Карензи М (2008). «Формируется ли европейское пространственное распределение ВИЧ-1-устойчивого аллеля CCR5-Δ32 в результате распада патоценоза из-за исторической римской экспансии?». Инфекция, генетика и эволюция. 8 (6): 864–874. Дои:10.1016 / j.meegid.2008.08.007. PMID 18790087.
- ^ Доусон Т.С., Бек М.А., Кузиэль В.А., Хендерсон Ф., Маеда Н. (2000). «Противопоставление эффектов дефицита CCR5 и CCR2 в воспалительном ответе легких на вирус гриппа A». Американский журнал патологии. 156 (6): 1951–1959. Дои:10.1016 / S0002-9440 (10) 65068-7. ЧВК 1850091. PMID 10854218.
- ^ Escosteguy Vargas A, Cechim G, Ferraz Correa J, Gomes PA, de Souza Macedo G, de Medeiros RM и др. (2009). «Плюсы и минусы отсутствующего хемокинового рецептора. Комментарии на тему« Формируется ли европейское пространственное распределение ВИЧ-1-устойчивого аллеля CCR5-Δ32 в результате разрушения патоценоза из-за исторической римской экспансии? »Эрика Фор и Мануэла Ройер. -Carenzi (2008) ». Инфекция, генетика и эволюция. 9 (4): 387–389. Дои:10.1016 / j.meegid.2009.01.001. PMID 19472441.
- ^ O'Brien SJ, Dean M (сентябрь 1997 г.). «В поисках генов устойчивости к СПИДу». Scientific American. 277 (3): 44–51. Bibcode:1997SciAm.277c..44O. Дои:10.1038 / scientificamerican0997-44. PMID 9274039.
- ^ а б c McEvedy C (февраль 1988 г.). «Бубонная чума». Scientific American. 258 (2): 118–23. Bibcode:1988SciAm.258b.118M. Дои:10.1038 / scientificamerican0288-118. PMID 3055286.
- ^ а б Камбуз C (1998). Демография ранних современных городов: Йорк в шестнадцатом и семнадцатом веках. Лондон: Макмиллан.
- ^ Хэтчер Дж. (1977). Чума, население и английская экономика 1348-1530 гг.. Лондон: Макмиллан.
- ^ Готфрид Р. (1983). Черная смерть: природные и гуманитарные катастрофы в средневековой Европе. Нью-Йорк: Свободная пресса.
- ^ а б Килинг MJ, Гиллиган Калифорния (ноябрь 2000 г.). «Бубонная чума: метапопуляционная модель зооноза». Труды Королевского общества B: биологические науки. 267 (1458): 2219–30. Дои:10.1098 / rspb.2000.1272. ЧВК 1690796. PMID 11413636.
- ^ Appleby AB (1980). «Исчезновение чумы: непрекращающаяся загадка». Обзор экономической истории. 33 (2): 161–73. Дои:10.2307/2595837. JSTOR 2595837. PMID 11614424.
- ^ Рассел Дж. (1948). Британское средневековое население. Альбукерке: Издательство Университета Нью-Мексико.
- ^ Твигг G (1984). Черная смерть: биологическая переоценка. Лондон: Батсфод.
- ^ Чапин С (1913). "Различия в типах инфекционных заболеваний, как показывает история оспы в Соединенных Штатах, 1815-1912 гг.". J Infect Dis. 13 (2): 171–196. Дои:10.1093 / infdis / 13.2.171.
- ^ а б Феннер Ф (1998). Историческая виньетка: жизнь с поксвирусами и издателями. Достижения в вирусных исследованиях. 51. С. 1–33. Дои:10.1016 / S0065-3527 (08) 60782-4. ISBN 9780120398515. PMID 9891584.
- ^ Лалани А.С., Мастерс Дж., Зенг В., Барретт Дж., Панну Р., Эверетт Х., Арендт К.В., Макфадден Дж. (Декабрь 1999 г.). «Использование хемокиновых рецепторов поксвирусами». Наука. 286 (5446): 1968–71. Дои:10.1126 / science.286.5446.1968. PMID 10583963.
- ^ Новембре Дж., Гальвани А.П., Слаткин М. (ноябрь 2005 г.). «Географическое распространение аллеля устойчивости к ВИЧ CCR5 Delta32». PLOS Биология. 3 (11): e339. Дои:10.1371 / journal.pbio.0030339. ЧВК 1255740. PMID 16216086.
- ^ Klein RS (январь 2008 г.). «Движущаяся цель: многочисленные роли CCR5 в инфекционных заболеваниях». Журнал инфекционных болезней. 197 (2): 183–6. Дои:10.1086/524692. PMID 18179384.
- ^ Киндберг Э., Микиене А., Акс С, Акерлинд Б., Вен С., Линдквист Л., Лундквист А., Свенссон Л. (январь 2008 г.). «Делеция в гене хемокинового рецептора 5 (CCR5) связана с клещевым энцефалитом». Журнал инфекционных болезней. 197 (2): 266–9. Дои:10.1086/524709. PMID 18179389.
- ^ Лим Дж. К., Луи С.Й., Глейзер С., Джин С., Джонсон Б., Джонсон Х., Макдермотт Д.Х., Мерфи П.М. (январь 2008 г.). «Генетический дефицит хемокинового рецептора CCR5 является сильным фактором риска симптоматической инфекции вируса Западного Нила: метаанализ 4 когорт эпидемии в США». Журнал инфекционных болезней. 197 (2): 262–5. Дои:10.1086/524691. PMID 18179388.
- ^ Glass WG, Лим Дж. К., Холера Р., Плетнев А. Г., Гао Дж. Л., Мерфи П. М. (октябрь 2005 г.). «Хемокиновый рецептор CCR5 способствует переносу лейкоцитов в мозг и выживанию при вирусной инфекции Западного Нила». Журнал экспериментальной медицины. 202 (8): 1087–98. Дои:10.1084 / jem.20042530. ЧВК 2213214. PMID 16230476.
- ^ а б Штейнбергер П., Андрис-Видхопф Дж., Бюлер Б., Торбетт Б.Э., Барбас К.Ф. (январь 2000 г.). «Функциональная делеция рецептора CCR5 с помощью внутриклеточной иммунизации дает клетки, устойчивые к CCR5-зависимой ВИЧ-1 инфекции и слиянию клеток». Труды Национальной академии наук Соединенных Штатов Америки. 97 (2): 805–10. Bibcode:2000ПНАС ... 97..805С. Дои:10.1073 / pnas.97.2.805. ЧВК 15412. PMID 10639161.
- ^ Schoofs M (2007-11-07). «Врач, мутация и потенциальное лекарство от СПИДа». Журнал "Уолл Стрит. Получено 2010-12-15.
- ^ Agrawal L, Lu X, Qingwen J, VanHorn-Ali Z, Nicolescu IV, McDermott DH, Murphy PM, Alkhatib G (март 2004 г.). «Роль белка CCR5Delta32 в устойчивости к вирусу иммунодефицита человека типа 1 R5, R5X4 и X4 в первичных CD4 + клетках». Журнал вирусологии. 78 (5): 2277–87. Дои:10.1128 / JVI.78.5.2277-2287.2004. ЧВК 369216. PMID 14963124.
- ^ «Аутологичные Т-клетки, генетически модифицированные в гене CCR5 нуклеазами цинковых пальцев SB-728 для ВИЧ (Zinc-Finger)». Национальные институты здравоохранения США. 2009-12-09. Получено 2009-12-30.
- ^ Уэйд Н. (28 декабря 2009 г.). «Цинковые пальцы могут стать ключом к возрождению генной терапии». Нью-Йорк Таймс. Получено 2009-12-30.
- ^ Corbyn Z (июль 2012 г.). «Запущен план по поиску лекарства от ВИЧ». Ланцет. 380 (9838): 203–4. Дои:10.1016 / с0140-6736 (12) 61199-4. PMID 22826833. S2CID 34773481.
- ^ Гонсалес Г., Парк С., Чен Д., Армитаж С., Шпалл Е., Берингер Р. (сентябрь 2011 г.). «Идентификация и частота CCR5Δ32 / Δ32 ВИЧ-устойчивых единиц пуповинной крови в больницах Хьюстона». ВИЧ Медицина. 12 (8): 481–6. Дои:10.1111 / j.1468-1293.2010.00911.x. ЧВК 4021858. PMID 21375684.
- ^ Петц Л.Д., Бернетт Дж. К., Ли Х, Ли С., Тонай Р., Бакалинская М., Шпалл Э. Дж., Армитаж С., Курцберг Дж., Реган Д. М., Кларк П., Куэрол С., Гутман Дж. А., Спеллман С. Р., Грагерт Л., Росси Дж. Дж. (2015) . «Прогресс в лечении ВИЧ-инфекции с помощью трансплантации гемопоэтических клеток». Стволовые клетки и клонирование: достижения и приложения. 8: 109–16. Дои:10.2147 / SCCAA.S56050. ЧВК 4524463. PMID 26251620.
- ^ Бегли С. (28 ноября 2018 г.). «На фоне шумихи китайский ученый выступает за создание генно-отредактированных младенцев». СТАТ.
- ^ Коллинз Ф.С. (28.11.2018). "Заявление китайского исследователя о первых младенцах, отредактированных генами". Национальные институты здоровья (NIH). Министерство здравоохранения и социальных служб США.
дальнейшее чтение
- Уилкинсон Д. (сентябрь 1996 г.). «Кофакторы предоставляют ключи входа. ВИЧ-1». Текущая биология. 6 (9): 1051–3. Дои:10.1016 / S0960-9822 (02) 70661-1. PMID 8805353. S2CID 18710567.
- Бродер CC, Димитров Д.С. (1996). «ВИЧ и рецепторы трансмембранного домена 7». Патобиология. 64 (4): 171–9. Дои:10.1159/000164032. PMID 9031325.
- Чхве Х., Мартин К.А., Фарзан М., Содроски Дж., Джерард Н.П., Джерард С. (июнь 1998 г.). «Структурные взаимодействия между рецепторами хемокинов, gp120 Env и CD4». Семинары по иммунологии. 10 (3): 249–57. Дои:10.1006 / smim.1998.0127. PMID 9653051.
- Шеппард Х.В., Селум С., Майкл Н.Л., О'Брайен С., Дин М., Кэррингтон М., Дондеро Д., Бухбиндер С.П. (март 2002 г.). «Инфекция ВИЧ-1 у лиц с генотипом CCR5-Delta32 / Delta32: приобретение вируса, индуцирующего синцитий, при сероконверсии». Журнал синдромов приобретенного иммунодефицита. 29 (3): 307–13. Дои:10.1097/00042560-200203010-00013. PMID 11873082.
- Фридман Б.Д., Лю QH, Дель Корно М., Коллман Р.Г. (2003). «Передача сигналов, опосредованная хемокиновым рецептором gp120 ВИЧ-1 в макрофагах человека». Иммунологические исследования. 27 (2–3): 261–76. Дои:10.1385 / ИК: 27: 2-3: 261. PMID 12857973. S2CID 32006625.
- Esté JA (сентябрь 2003 г.). «Попадание вирусов в качестве мишени для интервенции против ВИЧ». Современная лекарственная химия. 10 (17): 1617–32. Дои:10.2174/0929867033457098. PMID 12871111.
- Галло С.А., Финнеган С.М., Виард М., Равив Ю., Димитров А., Рават С.С., Пури А., Дурелл С., Блюменталь Р. (июль 2003 г.). «Реакция слияния, опосредованная Env ВИЧ». Biochimica et Biophysica Acta (BBA) - Биомембраны. 1614 (1): 36–50. Дои:10.1016 / S0005-2736 (03) 00161-5. PMID 12873764.
- Зайцева М., Педен К., Голдинг Х (июль 2003 г.). «Корецепторы ВИЧ: роль структуры, посттрансляционных модификаций и интернализации в слиянии вирусных клеток и в качестве мишеней для ингибиторов входа». Biochimica et Biophysica Acta (BBA) - Биомембраны. 1614 (1): 51–61. Дои:10.1016 / S0005-2736 (03) 00162-7. PMID 12873765.
- Ли К., Лю К. Х., Томкович Б., Йи Ю., Фридман Б. Д., Коллман Р. Г. (ноябрь 2003 г.). «Активация макрофагов посредством CCR5- и CXCR4-опосредованных сигнальных путей, вызванных gp120». Журнал биологии лейкоцитов. 74 (5): 676–82. Дои:10.1189 / jlb.0503206. PMID 12960231. S2CID 11362623.
- Йи Й, Ли К., Лю К. Х., Фридман Б. Д., Коллман Р. Г. (2004). «Использование рецепторов хемокинов и передача сигналов макрофагов вирусом иммунодефицита человека типа 1 gp120: значение для нейропатогенеза». Журнал нейровирологии. 10 (Дополнение 1): 91–6. Дои:10.1080/753312758. PMID 14982745. S2CID 9065929.
- Зайберт С., Сакмар Т.П. (2004). «Низкомолекулярные антагонисты CCR5 и CXCR4: новый многообещающий класс препаратов против ВИЧ-1». Текущий фармацевтический дизайн. 10 (17): 2041–62. Дои:10.2174/1381612043384312. PMID 15279544.
- Катлер К. В., Джотвани Р. (2006). «Экспрессия в слизистой оболочке полости рта рецепторов, корецепторов и альфа-дефенсинов ВИЧ-1: таблица устойчивости или восприимчивости к ВИЧ-инфекции?». Достижения в стоматологических исследованиях. 19 (1): 49–51. Дои:10.1177/154407370601900110. ЧВК 3750741. PMID 16672549.
- Ajuebor MN, Кэри JA, Swain MG (август 2006 г.). "CCR5 при Т-клеточных заболеваниях печени: что происходит?". Журнал иммунологии. 177 (4): 2039–45. Дои:10.4049 / jimmunol.177.4.2039. PMID 16887960.
- Липп М, Мюллер Г (2003). «Формирование адаптивного иммунитета: влияние CCR7 и CXCR5 на трафик лимфоцитов». Verhandlungen der Deutschen Gesellschaft für Pathologie. 87: 90–101. PMID 16888899.
- Балистрери ЧР, Карузо С., Гримальди М.П., Листи Ф, Васто С., Орландо В., Кампанья А.М., Лио Д., Кандоре Г. (апрель 2007 г.). «Рецептор CCR5: биологические и генетические последствия возрастных заболеваний». Летопись Нью-Йоркской академии наук. 1100 (1): 162–72. Bibcode:2007НЯСА1100..162Б. Дои:10.1196 / летопись.1395.014. PMID 17460174. S2CID 8437349.
- Madsen HO, Poulsen K, Dahl O, Clark BF, Hjorth JP (март 1990 г.). «Ретропсевдогены составляют основную часть семейства генов альфа фактора удлинения 1 человека». Исследования нуклеиновых кислот. 18 (6): 1513–6. Дои:10.1093 / nar / 18.6.1513. ЧВК 330519. PMID 2183196.
- Уэцуки Т., Наито А., Нагата С., Казиро Ю. (апрель 1989 г.). «Выделение и характеристика человеческого хромосомного гена фактора удлинения полипептидной цепи-1 альфа». Журнал биологической химии. 264 (10): 5791–8. PMID 2564392.
- Whiteheart SW, Shenbagamurthi P, Chen L, Cotter RJ, Hart GW (август 1989 г.). «Мышиный фактор элонгации 1 альфа (EF-1 альфа) посттрансляционно модифицирован новыми амидно-связанными этаноламин-фосфоглицериновыми фрагментами. Добавление этаноламин-фосфоглицерина к специфическим остаткам глутаминовой кислоты на EF-1 альфа». Журнал биологической химии. 264 (24): 14334–41. PMID 2569467.
- Энн Д.К., Ву М.М., Хуанг Т., Карлсон Д.М., Ву Р. (март 1988 г.). «Регулируемая ретинолом экспрессия гена в трахеобронхиальных эпителиальных клетках человека. Повышенная экспрессия фактора элонгации EF-1 альфа». Журнал биологической химии. 263 (8): 3546–9. PMID 3346208.
- Brands JH, Maassen JA, van Hemert FJ, Amons R, Möller W. (февраль 1986 г.). «Первичная структура альфа-субъединицы фактора удлинения человека 1. Структурные аспекты сайтов связывания гуанин-нуклеотидов». Европейский журнал биохимии / FEBS. 155 (1): 167–71. Дои:10.1111 / j.1432-1033.1986.tb09472.x. PMID 3512269.
внешняя ссылка
- CCR5 - Хемокиновый рецептор 5 с мотивом C-C (ген / псевдоген) - Homo sapiens (человек) Информация из База данных NCBI Gene
- Кристаллическая структура рецептора хемокина CCR5 Вращаемое и масштабируемое трехмерное изображение с сайта rcsb.org
- Видео и текст из документального фильма PBS об открытии CCR5
- «Хемокиновые рецепторы: CCR5». База данных рецепторов и ионных каналов IUPHAR. Международный союз фундаментальной и клинической фармакологии.
- Литература HuGENavigator по ВИЧ-инфекции и CCR5 из CDC - (обратите внимание, авторы не могут быть сотрудниками CDC, и на странице нет уведомления об общественном достоянии, поэтому нельзя предполагать, что это всеобщее достояние )
- Schering-Plough начинает фазу III исследований CCR5-викривирока у опытных пациентов с ВИЧ.
- HIVcoPred Сервер для прогнозирования использования корецепторов ВИЧ (CCR5). PLoS ONE 8 (4): e61437
- Человек CCR5 расположение генома и CCR5 страница сведений о генах в Браузер генома UCSC.
- Обзор всей структурной информации, доступной в PDB за UniProt: P51681 (C-C хемокиновый рецептор 5 типа) на PDBe-KB.