Рецептор вкуса - Taste receptor
| Рецептор вкуса | |
|---|---|
 Вкусовые рецепторы языка присутствуют во вкусовых сосочках сосочков. | |
| Идентификаторы | |
| FMA | 84662 |
| Анатомическая терминология | |
А вкусовый рецептор это тип рецептор что облегчает ощущение вкус. Когда пища или другие вещества попадают в рот, молекулы взаимодействуют со слюной и связываются со вкусовыми рецепторами в полости рта и других местах. Молекулы, придающие ощущение вкуса, считаются «живыми».[1]
Вкусовые рецепторы делятся на два семейства:[нужна цитата ]
- Тип 1, милая, впервые охарактеризованные в 2001 году:[2] TAS1R2 – TAS1R3
- Тип 2, Горький, впервые охарактеризованные в 2000 году:[3] У людей известно 25 различных рецепторов горечи, у кошек - 12, у кур - три, а у мышей - 35 известных различных рецепторов горечи.[4]
Визуальный, обонятельный, «сапиктивный» (восприятие вкусов), тройничный (горячий, холодный), механический - все они способствуют восприятию вкус. Из них переходный рецепторный потенциальный катионный канал подсемейства V член 1 (TRPV1 ) ваниллоидные рецепторы отвечают за восприятие тепла некоторыми молекулами, такими как капсаицин, и Рецептор CMR1 отвечает за восприятие холода такими молекулами, как ментол, эвкалиптол, и ицилин.[1]
Распределение тканей
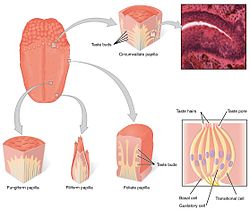
В вкусовая система состоит из вкусовых рецепторных клеток в вкусовые рецепторы. Вкусовые рецепторы, в свою очередь, содержатся в структурах, называемых сосочки. Во вкусе участвуют три типа сосочков: грибовидные сосочки, листовые сосочки, и огибающие сосочки. (Четвертый тип - нитевидные сосочки не содержат вкусовых рецепторов). Помимо сосочков вкусовые рецепторы также находятся в вкус и ранние части пищеварительная система словно гортань и верхний пищевод. Есть три черепные нервы которые иннервируют язык; в блуждающий нерв, языкоглоточный нерв, а лицевой нерв. В языкоглоточный нерв и chorda tympani филиал лицевой нерв иннервируют вкусовые рецепторы TAS1R и TAS2R. Наряду со вкусовыми рецепторами на языке эпителий кишечника также оснащен тонкой хемосенсорной системой, которая передает сенсорную информацию нескольким эффекторным системам, участвующим в регуляции аппетита, иммунных реакций и моторики желудочно-кишечного тракта. [5]
В 2010 году исследователи обнаружили горькие рецепторы в легочной ткани, которые вызывают расслабление дыхательных путей при попадании горького вещества. Они считают, что этот механизм является эволюционно адаптивным, поскольку помогает избавиться от инфекций легких, но также может быть использован для лечения астма и хроническая обструктивная болезнь легких.[6]
Функция
Вкус помогает идентифицировать токсины, поддерживать питание и регулируют аппетит, иммунные реакции и перистальтику желудочно-кишечного тракта.[5] Сегодня различают пять основных вкусов: соленый, сладкий, горький, кислый и умами. Ощущения соленого и кислого вкуса обнаруживаются через ионные каналы. Однако сладкий, горький вкус и вкус умами можно определить по G-протеин-связанный вкусовые рецепторы.[7]
Кроме того, некоторые агенты могут функционировать как модификаторы вкуса, так как миракулин или же куркулин для сладкого или стерубин к маска горькая.
Механизм действия
Стандартный рецептор вкуса горького, сладкого или умами - это Рецептор, связанный с G-белком с семью трансмембранные домены. Связывание лиганда на вкусовых рецепторах активирует второй посланник каскады для деполяризации вкусовой клетки. Gustducin является наиболее распространенной субъединицей Gα вкуса, играющей главную роль в восприятии горького вкуса TAS2R. Gustducin является гомологом трансдуцин, G-белок, участвующий в трансдукции зрения.[8] Кроме того, вкусовые рецепторы используют ионный канал TRPM5, а также фосфолипазу PLCβ2.[9]
Закуски или глутаматы (умами)
В TAS1R1 +TAS1R3 гетеродимерный рецептор функционирует как рецептор умами, реагируя на L-аминокислота связывание, особенно L-глутамат.[2] В умами вкус чаще всего ассоциируется с пищевой добавкой глутамат натрия (MSG) и может быть улучшен за счет привязки монофосфат инозина (IMP) и гуанозинмонофосфат (GMP) молекулы.[10][11] Клетки, экспрессирующие TAS1R1 + 3, находятся в основном в грибовидные сосочки на кончике и краях языка и вкусовых рецепторных клетках неба в нёбе.[2] Показано, что эти клетки синапсируют на chorda tympani нервы, чтобы посылать свои сигналы в мозг, хотя некоторая активация языкоглоточный нерв был найден.[10][12]
Альтернативные кандидаты вкусовых рецепторов умами включают варианты сплайсинга метаботропных рецепторов глутамата, mGluR4 и mGluR1, а N-метил-D-аспартат типа рецептора глутамат-ионного канала.[7][13][14][15]
Милая
В TAS1R2 +TAS1R3 гетеродимерный рецептор функционирует как милая рецептора путем связывания с широким спектром сахара и заменители сахара.[2][16] Клетки, экспрессирующие TAS1R2 + 3, обнаружены в округлые сосочки и листовые сосочки возле задней части язык и вкус вкусовые рецепторные клетки в нёбе.[2] Эти ячейки показаны синапс на chorda tympani и языкоглоточные нервы посылать свои сигналы в мозг.[7][12] В TAS1R3 гомодимер также действует как рецептор сладкого во многом так же, как TAS1R2 + 3, но имеет пониженную чувствительность к сладким веществам. Натуральный сахар легче обнаруживается TAS1R3 рецептор, чем заменители сахара. Это может помочь объяснить, почему сахар и искусственные подсластители имеют разные вкусы.[17] Генетический полиморфизм в TAS1R3 отчасти объясняет разницу в восприятии сладкого вкуса и потреблении сахара между людьми афроамериканского происхождения и людьми европейского и азиатского происхождения.[18][19]
Горький
Белки TAS2R (ИнтерПро: IPR007960 ) функционируют как Горький вкусовые рецепторы.[20] Есть 43 человека TAS2R гены, каждый из которых (за исключением пяти псевдогены ) не хватает интроны и коды для GPCR белок.[7] Эти белки, в отличие от белков TAS1R, имеют короткие внеклеточные домены и расположены в огибающие сосочки, вкус, листовые сосочки, и надгортанник вкусовые рецепторы, с пониженной экспрессией в грибовидные сосочки.[3][7] Хотя несомненно, что несколько TAS2R экспрессируются в одной клетке вкусового рецептора, до сих пор ведутся споры о том, могут ли млекопитающие различать вкусы разных горьких веществ. лиганды.[3][7] Однако должно происходить некоторое перекрытие, поскольку горьких соединений гораздо больше, чем генов TAS2R. Общие горькие лиганды включают: циклогексимид, денатоний, ПРОП (6-п-пропил-2-тиоурацил ), PTC (фенилтиокарбамид ) и β-глюкопиранозиды.[7]
Передача сигнала горьких раздражителей осуществляется через α-субъединицу густдуцин. Эта субъединица G-белка активирует вкус фосфодиэстераза и уменьшается циклический нуклеотид уровни. Дальнейшие шаги на пути трансдукции пока неизвестны. Βγ-субъединица густдуцина также опосредует вкус, активируя IP3 (инозитолтрифосфат ) и DAG (диглицерид ). Эти вторичные посланники могут открывать закрытые ионные каналы или вызывать выброс внутренних кальций.[21] Хотя все TAS2R расположены в клетках, содержащих густдуцин, нокаут густдуцина не устраняет полностью чувствительность к горьким соединениям, что предполагает избыточный механизм горького вкуса.[9] (неудивительно, учитывая, что горький вкус обычно сигнализирует о наличии токсин ).[9] Одним из предложенных механизмов для независимого от густдуцина горького вкуса является взаимодействие ионных каналов со специфическими горькими лигандами, подобное взаимодействию ионных каналов, которое происходит при дегустации кислых и соленых стимулов.[7]
Один из наиболее изученных белков TAS2R - это TAS2R38, что способствует дегустации как PROP, так и PTC. Это первый рецептор вкуса, чей полиморфизмы показано, что они ответственны за различия во вкусовом восприятии. Текущие исследования сосредоточены на определении других таких полиморфизмов, определяющих вкусовой фенотип.[7] Более поздние исследования показывают, что генетический полиморфизм в других генах рецепторов горького вкуса влияет на восприятие горького вкуса кофеина, хинина и бензоата денатония.[22]
Кислый
Исторически считалось, что кислый вкус получается только в свободном ионы водорода (ЧАС+) непосредственно деполяризованные вкусовые рецепторы. Однако сейчас предлагаются специфические рецепторы кислого вкуса с другими методами действия. HCN1 и HCN4 (Каналы HCN ) было два таких предложения; оба этих рецептора представляют собой каналы, управляемые циклическими нуклеотидами. Два ионных канала, которые, как предполагается, вносят вклад в кислый вкус: ACCN1 и ЗАДАЧА-1.
Соль
Также были предложены различные рецепторы для соленого вкуса, а также возможное определение вкуса липидов, сложных углеводов и воды. Однако данные об этих рецепторах в лучшем случае сомнительны и часто неубедительны в исследованиях на млекопитающих. Например, предлагаемый ENaC рецептор для определения натрия может быть показан только как способствующий вкусу натрия в Дрозофилия.[7]
Карбонизация
Фермент, связанный с кислым рецептором, передает информацию о газированной воде.[23]
Толстый
Возможный вкусовой рецептор жира, CD36, был идентифицирован.[24] CD36 был локализован в окружной и листовой сосочки, которые присутствуют в вкусовые рецепторы[25] и где лингвальная липаза продуцируется, и исследования показали, что рецептор CD36 связывает длинноцепочечные жирные кислоты.[26] Различия в количестве CD36 выражение у людей было связано с их способностью ощущать вкус жиров,[27] создание аргументов в пользу отношения рецептора к вкусу жира. Дальнейшие исследования рецептора CD36 могут быть полезны для определения существования настоящего рецептора вкуса жира.
GPR120 и GPR40 были вовлечены в реакцию на жир в полости рта,[28] и их отсутствие приводит к снижению предпочтения жиров и снижению нейрональной реакции на пероральные жирные кислоты.[29]
TRPM5 было показано, что он участвует в реакции орального жира и идентифицирован как возможный рецептор орального жира, но недавние данные показывают, что он в первую очередь находится ниже по течению.[30][31]
Типы
Гены рецепторов горького вкуса человека имеют названия от TAS2R1 до TAS2R64, с множеством пробелов из-за несуществующих генов, псевдогенов или предполагаемых генов, которые не были аннотированы в самой последней сборке генома человека. Многие гены рецепторов горького вкуса также имеют непонятные синонимы с несколькими разными названиями генов, относящимися к одному и тому же гену. В таблице ниже представлен полный список генов рецепторов горького вкуса человека:
| Учебный класс | Ген | Синонимы | Псевдонимы | Locus | Описание |
|---|---|---|---|---|---|
| Тип 1 (милая) | TAS1R1 | GPR70 | 1п36.23 | ||
| TAS1R2 | GPR71 | 1п36.23 | |||
| TAS1R3 | 1п36 | ||||
| тип 2 (Горький) | TAS2R1 | 5п15 | |||
| TAS2R2 | 7p21.3 | псевдоген | |||
| TAS2R3 | 7q31.3-q32 | ||||
| TAS2R4 | 7q31.3-q32 | ||||
| TAS2R5 | 7q31.3-q32 | ||||
| TAS2R6 | 7 | не аннотировано в сборке генома человека | |||
| TAS2R7 | 12p13 | ||||
| TAS2R8 | 12p13 | ||||
| TAS2R9 | 12p13 | ||||
| TAS2R10 | 12p13 | ||||
| TAS2R11 | отсутствует у людей | ||||
| TAS2R12 | TAS2R26 | 12p13.2 | псевдоген | ||
| TAS2R13 | 12p13 | ||||
| TAS2R14 | 12p13 | ||||
| TAS2R15 | 12p13.2 | псевдоген | |||
| TAS2R16 | 7q31.1-q31.3 | ||||
| TAS2R17 | отсутствует у людей | ||||
| TAS2R18 | 12p13.2 | псевдоген | |||
| TAS2R19 | TAS2R23, TAS2R48 | 12p13.2 | |||
| TAS2R20 | TAS2R49 | 12p13.2 | |||
| TAS2R21 | отсутствует у людей | ||||
| TAS2R22 | 12 | не аннотировано в сборке генома человека | |||
| TAS2R24 | отсутствует у людей | ||||
| TAS2R25 | отсутствует у людей | ||||
| TAS2R27 | отсутствует у людей | ||||
| TAS2R28 | отсутствует у людей | ||||
| TAS2R29 | отсутствует у людей | ||||
| TAS2R30 | TAS2R47 | 12p13.2 | |||
| TAS2R31 | TAS2R44 | 12p13.2 | |||
| TAS2R32 | отсутствует у людей | ||||
| TAS2R33 | 12 | не аннотировано в сборке генома человека | |||
| TAS2R34 | отсутствует у людей | ||||
| TAS2R35 | отсутствует у людей | ||||
| TAS2R36 | 12 | не аннотировано в сборке генома человека | |||
| TAS2R37 | 12 | не аннотировано в сборке генома человека | |||
| TAS2R38 | 7q34 | ||||
| TAS2R39 | 7q34 | ||||
| TAS2R40 | GPR60 | 7q34 | |||
| TAS2R41 | 7q34 | ||||
| TAS2R42 | 12p13 | ||||
| TAS2R43 | 12p13.2 | ||||
| TAS2R45 | GPR59 | 12 | |||
| TAS2R46 | 12p13.2 | ||||
| TAS2R50 | TAS2R51 | 12p13.2 | |||
| TAS2R52 | отсутствует у людей | ||||
| TAS2R53 | отсутствует у людей | ||||
| TAS2R54 | отсутствует у людей | ||||
| TAS2R55 | отсутствует у людей | ||||
| TAS2R56 | отсутствует у людей | ||||
| TAS2R57 | отсутствует у людей | ||||
| TAS2R58 | отсутствует у людей | ||||
| TAS2R59 | отсутствует у людей | ||||
| TAS2R60 | 7 | ||||
| TAS2R62P | 7q34 | псевдоген | |||
| ТАС2Р63П | 12p13.2 | псевдоген | |||
| TAS2R64P | 12p13.2 | псевдоген |
Потеря функции
У многих видов вкусовые рецепторы потеряли свои функции. Эволюционный процесс, в котором вкусовые рецепторы утратили свою функцию, считается адаптивной эволюцией, связанной с экологией кормления, определяющей специализацию и бифуркацию вкусовых рецепторов.[32] Показано, что из всех вкусовых рецепторов горький, сладкий и умами имеют корреляцию между инактивацией вкусовых рецепторов и пищевым поведением.[32] Однако нет убедительных доказательств того, что у каких-либо позвоночных отсутствуют гены рецепторов горького вкуса.[32]
Рецептор сладкого вкуса является одним из вкусовых рецепторов, функция которого утрачена. У млекопитающих преобладающим рецептором сладкого вкуса является вкусовой рецептор типа 1 Tas1r2 / Tas1r3.[33] Некоторые виды млекопитающих, такие как кошки и летучие мыши-вампиры, показали неспособность чувствовать сладкий вкус.[33] У этих видов причина потери функции сладкого рецептора связана с псевдогенизацией Tas1r2.[33] Псевдогенизация Tas1r2 также наблюдается у видов, не относящихся к млекопитающим, таких как куры и безъязычная западная когтистая лягушка, и эти виды также демонстрируют неспособность ощущать сладкий вкус.[33] Псевдогенизация Tas1r2 широко распространена и независима в отряде Carnivora.[33] Многие исследования показали, что псевдогенизация вкусовых рецепторов вызывается вредной мутацией в открытых рамках считывания (ORF).[34] В ходе исследования было обнаружено, что у хищных видов, отличных от кошачьих, эти виды демонстрировали мутации Tas1r2, нарушающие ORF, и они возникали независимо среди видов.[33] Они также показали высокую вариативность в своих родословных.[33] Предполагается, что псевдогенизация Tas1r2 произошла в результате конвергентной эволюции, когда плотоядные виды потеряли способность ощущать сладкий вкус из-за пищевого поведения.[33]
Умами также является вкусовым рецептором, функция которого у многих видов утрачена. Преобладающими рецепторами вкуса умами являются Tas1r1 / Tas1r3.[33] У двух линий водных млекопитающих, включая дельфинов и морских львов, было обнаружено, что Tas1r1 является псевдогенизированным.[33] Псевдогенизация Tas1r1 также была обнаружена у наземных плотоядных видов.[33] Хотя панда принадлежит к отряду Carnivora, она травоядна, где 99% ее рациона составляет бамбук, и она не может попробовать умами.[35] Последовательность генома панды показывает, что ее ген Tas1r1 псевдогенизирован.[35] В ходе исследования было обнаружено, что у всех видов отряда Carnivora, кроме панды, открытая рамка считывания сохранялась.[35] У панды соотношение несинонимичных и синонимичных замен оказалось намного выше, чем у других видов отряда Carnivora.[35] Эти данные коррелируют с датой окаменелостей панды, чтобы показать, где она перешла с диеты хищников на диету травоядных.[33] Таким образом, предполагается, что потеря функции умами у панды вызвана изменением диеты, когда панда стала меньше зависеть от мяса.[33] Однако эти исследования не объясняют травоядных животных, таких как лошади и коровы, которые сохранили рецептор Tas1r1.[35]
В целом, потеря функции вкусового рецептора - это эволюционный процесс, произошедший из-за изменения диеты у видов.[34]
Рекомендации
- ^ а б Это, Эрве (2012). «Наука о духовке - отрывок из главы 1». Получено 30 апреля 2014.
- ^ а б c d е Нельсон Г., Хун М.А., Чандрашекар Дж., Чжан Й., Рыба Н.Дж., Цукер С.С. (август 2001 г.). «Рецепторы сладкого вкуса млекопитающих». Клетка. 106 (3): 381–90. Дои:10.1016 / S0092-8674 (01) 00451-2. PMID 11509186.
- ^ а б c Адлер Э., Хун М.А., Мюллер К.Л., Чандрашекар Дж., Рыба Н.Дж., Цукер С.С. (март 2000 г.). «Новое семейство вкусовых рецепторов млекопитающих». Клетка. 100 (6): 693–702. Дои:10.1016 / S0092-8674 (00) 80705-9. PMID 10761934.
- ^ http://bitterdb.agri.huji.ac.il/dbbitter.php#receptorBrowse
- ^ а б Стинселс S, Depoortere I (2018). «Хеморецепторы в кишечнике». Ежегодный обзор физиологии. 80: 117–141. Дои:10.1146 / аннурьев-физиол-021317-121332. PMID 29029594.
- ^ Дешпанде Д.А., Ван В.К., Макилмойл Е.Л., Робинетт К.С., Шиллингер Р.М., Ан С.С., Шам Дж. С., Лиггетт С.Б. (ноябрь 2010 г.). «Рецепторы горького вкуса бронходилата гладкой мускулатуры дыхательных путей посредством локальной передачи сигналов кальция и обратной обструкции». Природа Медицина. 16 (11): 1299–304. Дои:10,1038 / нм.2237. ЧВК 3066567. PMID 20972434.
- ^ а б c d е ж грамм час я j Бахманов А.А., Бошам Г.К. (2007). «Гены вкусовых рецепторов». Ежегодный обзор питания. 27: 389–414. Дои:10.1146 / annurev.nutr.26.061505.111329. ЧВК 2721271. PMID 17444812.
- ^ Сайнс Э., Кавенаг М.М., Лопес Хименес Н.Д., Гутьеррес Дж. К., Бэтти Дж. Ф., Нортуп Дж. К., Салливан С. Л. (июнь 2007 г.). «Свойства связывания G-белка рецепторов сладкого и аминокислотного вкуса человека». Нейробиология развития. 67 (7): 948–59. Дои:10.1002 / dneu.20403. PMID 17506496.
- ^ а б c Чжан Й, Хун М.А., Чандрашекар Дж., Мюллер К.Л., Кук Б., Ву Д., Цукер С.С., Рыба, штат Нью-Джерси (февраль 2003 г.). «Кодирование сладкого, горького и умами вкусов: разные рецепторные клетки имеют сходные пути передачи сигналов». Клетка. 112 (3): 293–301. Дои:10.1016 / S0092-8674 (03) 00071-0. PMID 12581520.
- ^ а б Нельсон Дж., Чандрашекар Дж., Хун М.А., Фэн Л., Чжао Дж., Рыба Нью-Джерси, Цукер С.С. (март 2002 г.). «Аминокислотный рецептор вкуса». Природа. 416 (6877): 199–202. Bibcode:2002Натура.416..199Н. Дои:10.1038 / природа726. PMID 11894099.
- ^ Delay ER, Бивер А.Дж., Вагнер К.А., Стэплтон Дж. Р., Харбо Дж. О., Катрон К. Д., Ропер С. Д. (октябрь 2000 г.). «Синергия вкусовых предпочтений между агонистами рецептора глутамата и монофосфатом инозина у крыс». Химические чувства. 25 (5): 507–15. Дои:10.1093 / chemse / 25.5.507. PMID 11015322.
- ^ а б Данилова В, Хеллекант Г (март 2003). «Сравнение ответов барабанной хорды и языкоглоточного нервов на вкусовые стимулы у мышей C57BL / 6J». BMC Neuroscience. 4: 5. Дои:10.1186/1471-2202-4-5. ЧВК 153500. PMID 12617752.
- ^ Бренд JG (апрель 2000 г.). «Рецепторные и трансдукционные процессы для вкуса умами». Журнал питания. 130 (4S Доп.): 942S – 5S. Дои:10.1093 / jn / 130.4.942S. PMID 10736357.
- ^ Чаудхари Н., Лэндин А.М., Ропер С.Д. (февраль 2000 г.). «Вариант метаботропного рецептора глутамата действует как рецептор вкуса». Природа Неврология. 3 (2): 113–9. Дои:10.1038/72053. PMID 10649565.
- ^ Тойоно Т., Сета Й, Катаока С., Кавано С., Шигемото Р., Тойосима К. (июль 2003 г.). «Экспрессия метаботропных рецепторов глутамата I группы во вкусовых сосочках крыс». Исследования клеток и тканей. 313 (1): 29–35. Дои:10.1007 / s00441-003-0740-2. PMID 12898387.
- ^ Ли Х, Сташевский Л., Сюй Х, Дурик К., Золлер М., Адлер Э. (апрель 2002 г.). «Человеческие рецепторы сладкого вкуса и вкуса умами». Труды Национальной академии наук Соединенных Штатов Америки. 99 (7): 4692–6. Bibcode:2002PNAS ... 99.4692L. Дои:10.1073 / pnas.072090199. ЧВК 123709. PMID 11917125.
- ^ Zhao GQ, Zhang Y, Hoon MA, Chandrashekar J, Erlenbach I., Ryba NJ, Zuker CS (октябрь 2003 г.). «Рецепторы сладкого вкуса и вкуса умами млекопитающих». Клетка. 115 (3): 255–66. Дои:10.1016 / S0092-8674 (03) 00844-4. PMID 14636554.
- ^ Hwang LD, Lin C, Gharahkhani P, Cuellar-Partida G, Ong JS, An J, Gordon SD, Zhu G, MacGregor S, Lawlor DA, Breslin PA, Wright MJ, Martin NG, Reed DR (апрель 2019 г.). «Новое понимание сладкого вкуса человека: общегеномное ассоциативное исследование восприятия и потребления сладких веществ». Американский журнал клинического питания. 109: 1724–1737. Дои:10.1093 / ajcn / nqz043. ЧВК 6537940. PMID 31005972.
- ^ Юсиф, Рагид (март 2020 г.). «Изучение молекулярных взаимодействий между неокулином и рецепторами сладкого вкуса человека с помощью вычислительных подходов» (PDF). Sains Malaysiana. 49: 517. Дои:10.17576 / jsm-2020-4903-06.
- ^ Чандрашекар Дж., Мюллер К.Л., Хун М.А., Адлер Э., Фенг Л., Го В., Цукер С.С., Рыба Нью-Джерси (март 2000 г.). «T2R действуют как рецепторы горького вкуса». Клетка. 100 (6): 703–11. Дои:10.1016 / S0092-8674 (00) 80706-0. PMID 10761935.
- ^ Маргольское РФ (январь 2002 г.). «Молекулярные механизмы передачи горького и сладкого вкуса». Журнал биологической химии. 277 (1): 1–4. Дои:10.1074 / jbc.R100054200. PMID 11696554.
- ^ Hwang LD, Gharahkhani P, Breslin PA, Gordon SD, Zhu G, Martin NG, Reed DR, Wright MJ (сентябрь 2018 г.). «Двухфакторный анализ ассоциации всего генома усиливает роль кластеров горьких рецепторов на хромосомах 7 и 12 в горьком вкусе человека». BMC Genomics. 19 (1): 678. Дои:10.1186 / s12864-018-5058-2. ЧВК 6142396. PMID 30223776.
- ^ «Архивная копия». Архивировано из оригинал на 2015-07-03. Получено 2014-10-06.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Laugerette F, Passilly-Degrace P, Patris B, Niot I, Febbraio M, Montmayeur JP, Besnard P (ноябрь 2005 г.). «Участие CD36 в оросенсорном обнаружении пищевых липидов, спонтанном предпочтении жиров и пищеварительной секреции». Журнал клинических исследований. 115 (11): 3177–84. Дои:10.1172 / JCI25299. ЧВК 1265871. PMID 16276419.
- ^ Саймонс П.Дж., Куммер Дж. А., Лукен Дж. Дж., Бун Л. (декабрь 2011 г.). «Иммунолокализация апикального CD36 во вкусовых сосочках человека и свиньи из кольцевидных и листовых сосочков». Acta Histochemica. 113 (8): 839–43. Дои:10.1016 / j.acthis.2010.08.006. PMID 20950842.
- ^ Baillie AG, Coburn CT, Abumrad NA (сентябрь 1996 г.). «Обратимое связывание длинноцепочечных жирных кислот с очищенным FAT, гомологом CD36 жировой ткани». Журнал мембранной биологии. 153 (1): 75–81. Дои:10.1007 / s002329900111. PMID 8694909.
- ^ Пепино М.Ю., Лав-Грегори Л., Кляйн С., Абумрад Н.А. (март 2012 г.). «Ген транслоказы жирных кислот CD36 и лингвальная липаза влияют на оральную чувствительность к жиру у субъектов с ожирением». Журнал липидных исследований. 53 (3): 561–6. Дои:10.1194 / мл. M021873. ЧВК 3276480. PMID 22210925.
- ^ ДиПатрицио Н.В. (сентябрь 2014 г.). "Готов к прайм-тайму жирный вкус?". Физиология и поведение. 136: 145–54. Дои:10.1016 / j.physbeh.2014.03.002. ЧВК 4162865. PMID 24631296.
- ^ Картони К., Ясумацу К., Окури Т., Шигемура Н., Йошида Р., Годино Н., Ле Кутр Дж., Ниномия Ю., Дамак С. (июнь 2010 г.). «Вкусовые предпочтения жирных кислот опосредуются GPR40 и GPR120» (PDF). Журнал неврологии. 30 (25): 8376–82. Дои:10.1523 / JNEUROSCI.0496-10.2010. ЧВК 6634626. PMID 20573884.
- ^ Mattes RD (сентябрь 2011 г.). «Накапливающиеся данные подтверждают наличие у людей вкусовых компонентов свободных жирных кислот». Физиология и поведение. 104 (4): 624–31. Дои:10.1016 / j.physbeh.2011.05.002. ЧВК 3139746. PMID 21557960.
- ^ Лю П., Шах Б.П., Кроасделл С., Гилбертсон Т.А. (июнь 2011 г.). «Переходный канал потенциального рецептора типа M5 необходим для жирного вкуса». Журнал неврологии. 31 (23): 8634–42. Дои:10.1523 / JNEUROSCI.6273-10.2011. ЧВК 3125678. PMID 21653867.
- ^ а б c Фэн П, Чжао Х (июнь 2013 г.). «Сложная эволюционная история генов рецепторов сладкого / умами позвоночных». Китайский научный бюллетень. 58 (18): 2198–2204. Bibcode:2013ЧСБУ..58.2198Ф. Дои:10.1007 / s11434-013-5811-5.
- ^ а б c d е ж грамм час я j k л м Jiang P, Josue J, Li X, Glaser D, Li W, Brand JG, Margolskee RF, Reed DR, Beauchamp GK (март 2012 г.). «Сильная потеря вкуса у хищных млекопитающих». Труды Национальной академии наук Соединенных Штатов Америки. 109 (13): 4956–61. Дои:10.1073 / pnas.1118360109. ЧВК 3324019. PMID 22411809.
- ^ а б Антинуччи М., Риссо Д. (28 ноября 2017 г.). «Вопрос вкуса: специфическая потеря функции генов вкусовых рецепторов у позвоночных». Границы молекулярных биологических наук. 4: 81. Дои:10.3389 / fmolb.2017.00081. ЧВК 5712339. PMID 29234667.
- ^ а б c d е Чжао Х, Ян Дж., Сюй Х, Чжан Дж. (Декабрь 2010 г.). «Псевдогенизация гена вкусового рецептора умами Tas1r1 у гигантской панды совпала с ее диетическим переходом на бамбук». Молекулярная биология и эволюция. 27 (12): 2669–73. Дои:10.1093 / molbev / msq153. ЧВК 3108379. PMID 20573776.
внешняя ссылка
- Адлер Э., Хун М.А., Мюллер К.Л., Чандрашекар Дж., Рыба Дж. П., Цукер С.С., Паттон А. (2000). "Новое семейство вкусовых рецепторов млекопитающих - исследовательский обзор". Департамент биологии колледжа Дэвидсон. Получено 2008-08-11.
- вкус + рецепторы, + тип + 1 в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- вкус + рецепторы, + тип + 2 в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)