Сенсорно-моторная связь - Sensory-motor coupling
Сенсорно-моторная связь это соединение или интеграция сенсорная система и двигательная система. Сенсомоторная интеграция - это не статический процесс. Для данного стимул, нет единой команды двигателя. "Нейронные реакции почти на каждом этапе сенсомоторного пути изменяются в краткосрочной и долгосрочной перспективе за счет биофизический и синаптический процессы, повторяющийся и Обратная связь связи и учусь, а также многие другие внутренние и внешние переменные ».[1]
Обзор
Интеграция сенсорной и моторной систем позволяет животному принимать сенсорная информация и использовать его для полезных двигательных действий. Кроме того, выходные данные двигательной системы можно использовать для изменения реакции сенсорной системы на будущие стимулы.[1][2] Чтобы быть полезной, необходимо, чтобы сенсорно-моторная интеграция была гибкий процесс, потому что свойства мира и нас самих меняются со временем. Гибкая сенсомоторная интеграция позволит животному исправлять ошибки и быть полезным в различных ситуациях.[1][3] Для достижения желаемой гибкости возможно, что нервная система использовать использование внутренние модели и копии efference.[2][3][4]
Преобразование сенсорных координат в моторные
Перед движением текущее сенсорное состояние животного используется для генерации двигательной команды. Чтобы сгенерировать команду двигателя, сначала текущее сенсорное состояние сравнивается с желаемым или целевым состоянием. Затем нервная система преобразует сенсорные координаты в координаты двигательной системы, а двигательная система генерирует необходимые команды для движения мышц, чтобы достичь целевого состояния.[2]
Efference copy
Важным аспектом сенсомоторной интеграции является efference copy. Копия efference - это копия моторной команды, которая используется во внутренних моделях, чтобы предсказать, каким будет новое сенсорное состояние после завершения моторной команды. Копия efference может использоваться нервной системой для различения самогенерируемых изменений окружающей среды, сравнения ожидаемой реакции с тем, что на самом деле происходит в окружающей среде, и для увеличения скорости, с которой может быть отдана команда, путем прогнозирования состояния организма до получения сенсорного ввода.[2][5]
Внутренняя модель
Внутренняя модель - это теоретическая модель, используемая нервной системой для прогнозирования изменений окружающей среды, возникающих в результате двигательного действия. Предполагается, что нервная система имеет внутреннее представление о том, как двигательный аппарат, часть тела, которая будет перемещаться, ведет себя в окружающей среде.[6][7] Внутренние модели можно классифицировать как прямую или обратную модель.
Форвардная модель
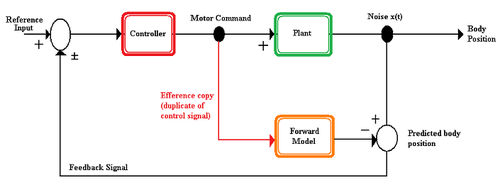
Прямая модель - это модель, используемая нервной системой для прогнозирования нового состояния двигательного аппарата и сенсорных стимулов, возникающих в результате движения. Прямая модель принимает копию эффекта в качестве входных данных и выводит ожидаемые сенсорные изменения.[4] Прямые модели предлагают организму несколько преимуществ.
Преимущества:
- Предполагаемое будущее состояние может использоваться для координации движения до возврата сенсорной обратной связи.[3][4]
- Выходные данные прямой модели можно использовать для различения стимулов, генерируемых самим собой, и стимулов, не генерируемых самим собой.[4]
- Предполагаемая сенсорная обратная связь может использоваться для изменения чувствительности животного. восприятие связанные с самопроизвольным движением.[3]
- Разницу между ожидаемым сенсорным состоянием и сенсорной обратной связью можно использовать для исправления ошибок в движении и модели.[3]
Обратная модель
Обратная модель ведет себя противоположно прямой модели. Обратные модели используются нервной системой для оценки двигательной команды, вызвавшей изменение сенсорной информации.[4] или для определения команды двигателя, которая достигнет целевого состояния.[6]
Примеры
Стабилизация взгляда
Во время полета мухе важно сохранять ровный взгляд; однако муха может вращаться. Вращение определяется визуально как вращение среды, называемое оптический поток. Затем входной оптический поток преобразуется в моторную команду для мышц шеи мухи, так что муха будет сохранять ровный взгляд. Этот рефлекс уменьшается у неподвижной мухи по сравнению с тем, когда она летит или идет.[1]
Поющие сверчки
Сверчки-самцы поют, потирая передние крылья друг о друга. Производимые звуки достаточно громкие, чтобы уменьшить крикет сверчка. слуховая система реакция на другие звуки. Эта десенсибилизация вызвана гиперполяризация нейрона Омега-1 (ON1), слуховой интернейрон, из-за активация слуховой стимуляцией.[5] Чтобы уменьшить самодесенсибилизацию, грудной отдел сверчка центральный генератор шаблонов отправляет побочный разряд, efference copy, который используется для подавления реакции организма на стимулы, генерируемые самим слуховой системой.[1][5] Последующий разряд используется для подавления реакции слуховой системы на собственную песню сверчка и предотвращения снижения чувствительности. Это торможение позволяет сверчку реагировать на внешние звуки, такие как песня соревнующегося самца.[8]
Речь
Сенсомоторная интеграция участвует в разработка, производство и восприятие речи.[9][10]
Развитие речи
Два ключевых элемента развития речи: лепет и прослушивание. Считается, что можно научиться связывать двигательное действие со слышимым звуком. Одна из причин этого в том, что глухой младенцы канонически не лепечут. Другое дело, что младенческий восприятие известно, что на него влияет его лепет. Одна модель развития речи предполагает, что звуки, производимые лепетом, сравниваются со звуками, издаваемыми на языке, который используется в окружении младенца, и усваивается ассоциация моторной команды со звуком.[10]
Производство речи
Прослушивание играет важную роль в производстве и поддержании речи. Например, люди, которые страдают глухотой у взрослых, теряют способность произносить точную речь. Это снижение связано с отсутствием слуховой обратной связи. Другой пример - приобретение нового акцент в результате проживания в районе с другим акцентом.[9] Эти изменения можно объяснить с помощью форвардной модели.
В этой прямой модели моторная кора посылает моторную команду в голосовой тракт и копия для внутренней модели речевого тракта. Внутренняя модель предсказывает, какие звуки будут издаваться. Этот прогноз используется для проверки того, что команда двигателя будет издавать звук цели, чтобы можно было внести исправления. Оценка внутренней модели также сравнивается с произведенным звуком, чтобы произвести оценку ошибки. Оценка ошибки используется для исправления внутренней модели. Обновленная внутренняя модель затем будет использоваться для генерации будущих команд двигателя.[9]
Восприятие речи
Сенсомоторная интеграция не критична для восприятия речи; однако он выполняет модулирующую функцию. Это подтверждается тем фактом, что люди, у которых либо нарушение речевой деятельности, либо неспособность говорить, по-прежнему способны воспринимать речь. Кроме того, эксперименты, в которых стимулировались двигательные области, связанные с речью, изменяли, но не препятствовали восприятию речи.[9]
Пациент Р.В.
Пациент Р.В. был мужчиной, у которого был поврежден теменный и затылочные доли, области мозга, связанные с обработка визуальной информации, из-за Инсульт. В результате инсульта он испытал головокружение когда он пытался следить глазами за движущимся объектом. Головокружение было вызвано тем, что его мозг интерпретировал мир как движущийся. У нормальных людей мир не воспринимается как движущийся при отслеживании объекта, несмотря на то, что изображение мира перемещается через сетчатка как движется глаз. Причина этого в том, что мозг предсказывает движение мира по сетчатке как следствие движения глаз. Однако R.W. не смог сделать этого прогноза.[3]
Расстройства
Болезнь Паркинсона
Пациенты с болезнь Паркинсона часто проявляются симптомы брадикинезия и гипометрия. Эти пациенты больше зависят от внешних сигналов, чем от проприоцепция и кинестезия по сравнению со здоровыми людьми.[11] Фактически, исследования с использованием внешних вибраций для создания проприоцептивных ошибок в движении показывают, что пациенты с болезнью Паркинсона работают лучше, чем здоровые люди. Исследователи также показали, что пациенты недооценивают движение конечности, когда она двигалась.[11] Дополнительно исследования по соматосенсорные вызванные потенциалы доказали, что моторные проблемы, вероятно, связаны с неспособностью правильно обрабатывать сенсорную информацию, а не с ее генерацией.
Хантингтона
Хантингтона пациенты часто имеют проблемы с блок управления двигателем. В обоих хинолиновый На моделях и пациентах было показано, что люди с синдромом Хантингтона обладают ненормальными сенсорными данными. Кроме того, у пациентов было показано снижение ингибирования испуганный рефлекс. Это уменьшение указывает на проблему с правильной сенсомоторной интеграцией. «Различные проблемы в интеграции сенсорной информации объясняют, почему пациенты с HD не могут контролировать произвольные движения точно ".[11]
Дистония
Дистония - еще одно моторное расстройство, которое проявляется нарушениями сенсомоторной интеграции. Есть несколько свидетельств того, что очаговая дистония связано с неправильным связыванием или обработкой афферентный сенсорная информация в двигательных областях мозга.[11] Например, от дистонии можно частично избавиться с помощью сенсорный трюк. Сенсорный трюк - это нанесение стимула на область рядом с местом, пораженным дистонией, которое приносит облегчение. Позитронно-эмиссионная томография исследования показали, что активность как в дополнительная моторная зона и первичная моторная кора уменьшаются сенсорным трюком. Необходимы дополнительные исследования дисфункции сенсомоторной интеграции, поскольку она связана с нефокальной дистонией.[11]
Синдром беспокойных ног
Синдром беспокойных ног (СБН) - сенсомоторное расстройство. Людей с RLS преследуют чувства дискомфорт и желание пошевелить ногами. Эти симптомы чаще всего возникают в состоянии покоя. Исследования показали, что моторная кора головного мозга увеличилась. возбудимость у пациентов с СБН по сравнению со здоровыми людьми. Соматосенсорные вызванные потенциалы от стимуляции обоих задний нерв и срединный нерв нормальные.[12] Нормальные SEP указывают на то, что RLS связан с аномальной сенсомоторной интеграцией. В 2010 году Винченцо Риццо и др. предоставили доказательства того, что страдающие RLS имеют более низкое, чем обычно, короткое латентное афферентное торможение (SAI), торможение моторной коры афферентными сенсорными сигналами. Снижение SAI указывает на наличие аномальной сенсомоторной интеграции у пациентов с RLS.[12]
Смотрите также
- Блок управления двигателем
- Моторное обучение
- Моторная цель
- Моторная координация
- Мультисенсорная интеграция
- Сенсорная обработка
Рекомендации
- ^ а б c d е Хьюстон, Стивен Дж; Джаяраман, Вивек (2011). «Изучение сенсомоторной интеграции у насекомых». Текущее мнение в нейробиологии. 21 (4): 527–534. Дои:10.1016 / j.conb.2011.05.030. ISSN 0959-4388. PMID 21705212.
- ^ а б c d Flanders M (февраль 2011 г.). «Что является биологической основой сенсомоторной интеграции?». Биол Киберн. 104 (1–2): 1–8. Дои:10.1007 / s00422-011-0419-9. ЧВК 3154729. PMID 21287354.
- ^ а б c d е ж Шадмер, Реза; Smith, Maurice A .; Кракауэр, Джон В. (2010). «Коррекция ошибок, сенсорное прогнозирование и адаптация в управлении двигателем» (PDF). Ежегодный обзор нейробиологии. 33 (1): 89–108. Дои:10.1146 / annurev-neuro-060909-153135. ISSN 0147-006X. PMID 20367317.
- ^ а б c d е Wolpert, D .; Гахрамани, Z; Джордан, М. (1995). «Внутренняя модель для сенсомоторной интеграции» (PDF). Наука. 269 (5232): 1880–1882. Дои:10.1126 / science.7569931. ISSN 0036-8075. PMID 7569931.
- ^ а б c Пуле Дж. Ф., Хедвиг Б. (март 2003 г.). «Механизм выброса следствия модулирует центральную слуховую обработку у поющих сверчков». J. Neurophysiol. 89 (3): 1528–40. Дои:10.1152 / ян.0846.2002. PMID 12626626.
- ^ а б Кавато М (декабрь 1999 г.). «Внутренние модели для управления двигателем и планирования траектории» (PDF). Текущее мнение в нейробиологии. 9 (6): 718–27. Дои:10.1016 / S0959-4388 (99) 00028-8. PMID 10607637.
- ^ Олово C, Poon CS (сентябрь 2005 г.). «Внутренние модели в сенсомоторной интеграции: перспективы теории адаптивного управления». J Neural Eng. 2 (3): S147–63. Дои:10.1088 / 1741-2560 / 2/3 / S01. ЧВК 2263077. PMID 16135881.
- ^ Уэбб Б. (май 2004 г.). «Нейронные механизмы для предсказания: есть ли у насекомых прямые модели?». Тенденции Neurosci. 27 (5): 278–82. Дои:10.1016 / j.tins.2004.03.004. PMID 15111010.
- ^ а б c d Хикок Дж., Хоуд Дж., Ронг Ф. (февраль 2011 г.). «Сенсомоторная интеграция в обработке речи: вычислительные основы и нейронная организация». Нейрон. 69 (3): 407–22. Дои:10.1016 / j.neuron.2011.01.019. ЧВК 3057382. PMID 21315253.
- ^ а б Вестерманн Г., Рек Миранда Е. (май 2004 г.). «Новая модель сенсомоторной связи в развитии речи». Brain Lang. 89 (2): 393–400. CiteSeerX 10.1.1.3.6041. Дои:10.1016 / S0093-934X (03) 00345-6. PMID 15068923.
- ^ а б c d е Abbruzzese G, Berardelli A (март 2003 г.). «Сенсомоторная интеграция при двигательных расстройствах». Mov. Disord. 18 (3): 231–40. Дои:10.1002 / mds.10327. PMID 12621626.
- ^ а б Риццо V, Арико I, Лиотта Г. и др. (Декабрь 2010 г.). «Нарушение сенсорно-моторной интеграции у пациентов, страдающих СБН». J. Neurol. 257 (12): 1979–85. Дои:10.1007 / s00415-010-5644-у. PMID 20635185.
Анатомия глобус из человеческий глаз | |||||||
|---|---|---|---|---|---|---|---|
| Волокнистая туника (внешний) |
|   | |||||
| Увеа / сосудистая оболочка (середина) |
| ||||||
| Сетчатка (внутренний) |
| ||||||
| Анатомические области глаза |
| ||||||
| Другой | |||||||