Нейротоксин - Neurotoxin

Нейротоксины находятся токсины которые разрушительны для нервная ткань (вызывая нейротоксичность ).[3] Нейротоксины - это обширный класс экзогенный химический неврологический оскорбления[4] которые могут отрицательно повлиять на функцию как развивающейся, так и зрелой нервной ткани.[5] Этот термин также может использоваться для классификации эндогенный соединения, которые при ненормальном контакте могут оказаться неврологически токсичными.[4] Хотя нейротоксины часто неврологически деструктивны, их способность целенаправленно воздействовать на нервные компоненты важна при изучении нервной системы.[6] Общие примеры нейротоксинов включают: вести,[7] этиловый спирт (употребление алкоголя),[8] глутамат,[9] оксид азота,[10] ботулинический токсин (например, ботокс),[11] столбнячный токсин,[12] и тетродотоксин.[6] Некоторые вещества, такие как оксид азота и глутамат, на самом деле необходимы для правильного функционирования организма и оказывают нейротоксическое действие только при чрезмерных концентрациях.
Нейротоксины подавляют нейрон контроль над ион концентрации по всему клетка мембрана[6] или связь между нейронами через синапс.[13] Местный патология воздействия нейротоксина часто включает нейрон эксайтотоксичность или же апоптоз[14] но может также включать глиальная клетка повреждать.[15] Макроскопические проявления воздействия нейротоксина могут включать широко распространенные Центральная нервная система ущерб, такой как Интеллектуальная недееспособность,[5] настойчивый объем памяти обесценения,[16] эпилепсия, и слабоумие.[17] Кроме того, нейротоксин-опосредованный периферическая нервная система ущерб, такой как невропатия или же миопатия обычное дело. Была показана поддержка ряда методов лечения, направленных на ослабление травм, опосредованных нейротоксинами, таких как антиоксидант[8] и антитоксин[18] администрация.
Фон
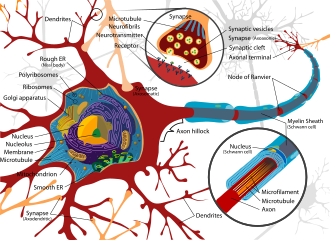
Воздействие нейротоксинов в обществе не ново,[19] поскольку цивилизации на протяжении тысячелетий подвергались воздействию неврологически деструктивных соединений. Одним из ярких примеров является возможное значительное воздействие свинца во время Римская империя в результате развития обширных водопроводные сети и привычка кипятить вино с уксусом в свинцовых кастрюлях, чтобы подсластить его, в процессе вырабатывается ацетат свинца, известный как «сахар свинца».[20] Частично нейротоксины были частью человек истории из-за хрупкой и уязвимой природы нервной системы, что делает ее очень подверженной нарушениям.
Нервная ткань, обнаруженная в мозг, спинной мозг, а периферия представляет собой чрезвычайно сложную биологическую систему, которая в значительной степени определяет многие уникальные черты людей. Однако, как и в случае любой очень сложной системы, даже небольшие возмущения в ее среде могут привести к значительным функциональным нарушениям. Свойства, приводящие к восприимчивости нервной ткани, включают большую площадь поверхности нейронов, высокую липид содержание, удерживающее липофильные токсины, высокое кровь приток к мозгу, вызывающий повышенное эффективное воздействие токсинов, и сохранение нейронов на протяжении всей жизни человека, что приводит к накоплению повреждений.[21] В результате нервная система имеет ряд механизмов, предназначенных для защиты от внутренних и внешних атак, включая гематоэнцефалический барьер.
В гематоэнцефалический барьер (BBB) является одним из важнейших примеров защиты, которая предотвращает попадание токсинов и других вредных соединений в мозг.[22] Поскольку мозг требует поступления питательных веществ и удаления отходов, он перфузируется кровотоком. Однако кровь может нести ряд проглоченных токсинов, которые могут вызвать значительную гибель нейронов, если они достигнут нервной ткани. Таким образом, защитные клетки получили название астроциты окружают капилляры в мозг и поглощают питательные вещества из крови и впоследствии транспортируют их к нейронам, эффективно изолируя мозг от ряда потенциальных химических повреждений.[22]
Этот барьер создает плотный гидрофобный слой вокруг капилляры в головном мозге, препятствуя транспортировке больших или гидрофильный соединения. Помимо BBB, сосудистое сплетение обеспечивает уровень защиты от поглощения токсинов в головном мозге. Сосудистые сплетения - это васкуляризированные слои ткани, расположенные в третьем, четвертом и латеральном отделах. желудочки головного мозга, которые через функцию своих эпендимальный клетки, отвечают за синтез спинномозговая жидкость (CSF).[23] Важно отметить, что путем выборочного прохождения ионы питательные вещества и ловушки тяжелые металлы такие как свинец, сосудистые сплетения поддерживают строго регулируемую среду, в которую входят головной и спинной мозг.[22][23]

Будучи гидрофобными и маленькими или подавляя функцию астроцитов, некоторые соединения, включая определенные нейротоксины, способны проникать в мозг и вызывать значительные повреждения. В наше время, ученые и врачи были поставлены перед проблемой идентификации и лечения нейротоксинов, что привело к растущему интересу как к исследованиям нейротоксикологии, так и к клиническим исследованиям.[24] Хотя клиническая нейротоксикология - в значительной степени развивающаяся область, значительные успехи были сделаны в идентификации многих нейротоксинов окружающей среды, что привело к классификации от 750 до 1000 известных потенциально нейротоксичных соединений.[21] Из-за критической важности обнаружения нейротоксинов в обычных средах, специальные протоколы были разработаны Агентство по охране окружающей среды США (EPA) для тестирования и определения нейротоксического действия соединений (USEPA 1998). Кроме того, in vitro системы стали чаще использоваться, поскольку они обеспечивают значительные улучшения по сравнению с более распространенными in vivo системы прошлого. Примеры улучшений включают податливую, однородную среду и устранение загрязняющих эффектов системного метаболизма.[24] Системы in vitro, однако, создают проблемы, поскольку было трудно должным образом воспроизвести сложности нервной системы, такие как взаимодействия между поддерживающими астроцитами и нейронами при создании ГЭБ.[25] Чтобы еще больше усложнить процесс определения нейротоксинов при тестировании in vitro, нейротоксичность и цитотоксичность может быть трудно отличить, поскольку непосредственное воздействие на нейроны соединений может быть невозможно in vivo, как это происходит in vitro. Кроме того, ответ клетки к химическим веществам может неточно передавать различие между нейротоксинами и цитотоксинами, поскольку такие симптомы, как окислительный стресс или же скелетный модификации могут происходить в ответ на любой из них.[26]
Чтобы решить эту проблему, нейрит разрастания (аксональные или дендритные) в ответ на применяемые соединения недавно были предложены как более точное различие между настоящими нейротоксинами и цитотоксины в среде тестирования in-vitro. Однако из-за значительных неточностей, связанных с этим процессом, он медленно получил широкую поддержку.[27] Кроме того, биохимические механизмы стали более широко использоваться в тестировании нейротоксинов, так что соединения могут быть проверены на достаточность для индукции вмешательства клеточных механизмов, таких как ингибирование ацетилхолинэстераза емкость органофосфаты (включает ДДТ и зарин газ).[28] Хотя методы определения нейротоксичности все еще требуют значительного развития, идентификация вредных соединений и симптомов воздействия токсинов претерпела значительные улучшения.
Приложения в нейробиологии
Несмотря на то, что нейротоксины различаются по химическим свойствам и функциям, они обладают общим свойством: они действуют посредством определенного механизма, приводящего либо к нарушению, либо к разрушению необходимых компонентов в организме. нервная система. Однако нейротоксины по своей природе могут быть очень полезны в области нейробиология. Поскольку нервная система большинства организмов очень сложна и необходима для выживания, она, естественно, стала мишенью для нападений как хищников, так и жертв. В качестве ядовитые организмы часто используют свои нейротоксины, чтобы очень быстро подчинить себе хищника или жертву, токсины эволюционировали, чтобы стать высокоспецифичными для своих целевых каналов, так что токсин с трудом связывает другие цели[29] (видеть Токсины ионного канала). Таким образом, нейротоксины являются эффективным средством, с помощью которого можно точно и эффективно воздействовать на определенные элементы нервной системы. Ранний пример использования таргетинга на основе нейротоксинов радиоактивно меченый тетродотоксин для анализа натриевые каналы и получить точные измерения их концентрации вдоль нервной мембраны.[29] Точно так же за счет изоляции определенных каналов активности нейротоксины предоставили возможность улучшить исходный Модель Ходжкина-Хаксли нейрона, в котором предполагалось, что единственный родовой натрий и калиевые каналы может составлять большинство функций нервной ткани.[29] Исходя из этого основного понимания, использование обычных соединений, таких как тетродотоксин, тетраэтиламмоний, и бунгаротоксины привели к гораздо более глубокому пониманию различных способов поведения отдельных нейронов.
Механизмы деятельности
Поскольку нейротоксины представляют собой соединения, которые отрицательно влияют на нервную систему, ряд механизмов, посредством которых они действуют, заключается в ингибировании клеточных процессов нейронов. Эти ингибированные процессы могут варьироваться от механизмов деполяризации мембраны до межнейронная связь. Подавляя способность нейронов выполнять ожидаемые внутриклеточные функции или передавать сигнал соседней клетке, нейротоксины могут вызывать остановку системной нервной системы, как в случае ботулинический токсин,[13] или даже смерть нервной ткани.[30] Время, необходимое для появления симптомов при воздействии нейротоксина, может варьироваться для разных токсинов, составляя порядка часов для ботулинического токсина.[18] и годы на свинец.[31]
| Классификация нейротоксинов | Нейротоксины |
|---|---|
| Ингибиторы Na-каналов | Тетродотоксин[6] |
| Ингибиторы К-канала | Тетраэтиламмоний[32] |
| Ингибиторы каналов Cl | Хлоротоксин,[33] |
| Ингибиторы Са-канала | Конотоксин[34] |
| Ингибиторы высвобождения синаптических везикул | Ботулинический токсин,[35] столбнячный токсин[36] |
| Ингибиторы рецепторов | Бунгаротоксин[37] |
| Агонисты рецепторов | 25I-NBOMe[39] |
| Ингибиторы гематоэнцефалического барьера | Меркурий[41] |
| Вмешательство цитоскелета | Мышьяк,[42] аммиак[43] |
| Са-опосредованная цитотоксичность | Свинец[44] |
| Множественные эффекты | Этиловый спирт[45][46] |
| Источники эндогенных нейротоксинов | Оксид азота,[47] Глутамат,[48] Дофамин[49] |
Ингибиторы
Натриевый канал
Тетродотоксин

Тетродотоксин (ТТХ) - это яд, вырабатываемый организмами, принадлежащими к Порядок Tetraodontiformes, который включает рыба фугу, океанская солнечная рыба, и рыба-дикобраз.[50] Внутри рыбы фугу TTX находится в печень, гонады, кишечник, и кожа.[6][51] ТТХ может быть смертельным при употреблении и стал распространенной формой отравления во многих странах. Общие симптомы потребления ТТХ включают: парестезия (часто ограничивается рот и конечности ), мышечная слабость, тошнота, и рвота[50] и часто проявляются в течение 30 минут после проглатывание.[52] Основной механизм токсичности ТТХ заключается в подавлении функции натриевых каналов, что снижает функциональную способность нейронной связи. Это ингибирование в значительной степени влияет на чувствительную подгруппу натриевых каналов, известных как TTX-чувствительные (TTX-s), которые также в значительной степени ответственны за натриевый ток, который управляет фаза деполяризации нейрона потенциалы действия.[6]

ТТХ-резистентный (ТТХ-r) - еще одна форма натриевых каналов, которая имеет ограниченную чувствительность к ТТХ и в основном встречается в аксоны малого диаметра такие как найденные в ноцицепционные нейроны.[6] Когда поступает значительный уровень ТТХ, он связывает натриевые каналы на нейронах и снижает их проницаемость мембраны к натрию. Это приводит к увеличению эффективного порога необходимых возбуждающих сигналов, чтобы вызвать потенциал действия в постсинаптическом нейроне.[6] Эффект от этого увеличился порог сигнализации пониженная возбудимость постсинаптические нейроны и последующая потеря двигательной и сенсорной функции, которая может привести к параличу и смерти. Хотя вспомогательная вентиляция легких может увеличить шанс выживания после воздействия ТТХ, в настоящее время антитоксина нет. Использование ингибитора ацетилхолинэстеразы Неостигмин или мускариновый ацетилхолин антагонист Атропин (что подавляет парасимпатическую активность), однако может увеличить активность симпатического нерва достаточно, чтобы повысить шансы на выживание после воздействия ТТХ.[50]
Калиевый канал
Тетраэтиламмоний
Тетраэтиламмоний (ТЭА) - это соединение, которое, как и ряд нейротоксинов, было впервые идентифицировано благодаря его повреждающему действию на нервную систему, и было показано, что оно обладает способностью подавлять функцию двигательных нервов и, таким образом, сокращать нервную систему. мускулатура аналогично кураре.[53] Кроме того, хроническое введение TEA может вызвать мышечную атрофию.[53] Позже было установлено, что TEA функционирует in vivo прежде всего благодаря своей способности ингибировать оба калиевых канала, ответственных за выпрямитель с задержкой видел в потенциал действия и некоторая популяция кальций-зависимых калиевых каналов.[32] Именно эта способность подавлять приток калия в нейроны сделала TEA одним из наиболее важных инструментов в нейробиологии. Было высказано предположение, что способность TEA ингибировать калиевые каналы происходит от его структуры, заполняющей пространство, аналогичной ионам калия.[53] Что делает чай очень полезным для нейробиологи это его специфическая способность устранять активность калиевых каналов, что позволяет изучать вклад нейронов в ответ других ионных каналов, таких как потенциалзависимые натриевые каналы.[54] В дополнение к его многочисленным применениям в исследованиях неврологии, TEA, как было показано, действует как эффективное лечение болезнь Паркинсона благодаря своей способности ограничивать прогрессирование болезни.[55]
Хлоридный канал
Хлоротоксин
Хлортоксин (Cltx) - это активное соединение, содержащееся в скорпион яд, и в первую очередь токсичен из-за его способности подавлять проводимость хлоридные каналы.[33] Проглатывание летальных объемов Cltx приводит к параличу из-за нарушения этого ионного канала. Было показано, что, как и ботулинический токсин, Cltx обладает значительной терапевтической ценностью. Доказано, что Cltx может подавлять способность к глиомы для проникновения в здоровую нервную ткань головного мозга, что значительно снижает потенциальный инвазивный вред, причиняемый опухолями.[56][57]
Кальциевый канал
Конотоксин
Конотоксины представляют собой категорию ядов, вырабатываемых морской улиткой-конусом, и способны ингибировать активность ряда ионных каналов, таких как кальциевые, натриевые или калиевые каналы.[58][59] Во многих случаях токсины, выделяемые различными типами конусные улитки включают ряд различных типов конотоксинов, которые могут быть специфичными для разных ионных каналов, создавая, таким образом, яд, способный к обширному нарушению нервной функции.[58] Одна из уникальных форм конотоксинов, ω-конотоксин (ω-CgTx ) очень специфичен для Са-каналов и показал свою полезность при их изоляции от системы.[60] Поскольку приток кальция необходим для правильной возбудимости клетки, любое значительное ингибирование может помешать большой функциональности. Примечательно, что ω-CgTx способен долгое время связываться и ингибировать потенциал-зависимые кальциевые каналы, расположенные в мембранах нейронов, но не в мембранах мышечных клеток.[61]
Высвобождение синаптических везикул
Ботулинический токсин
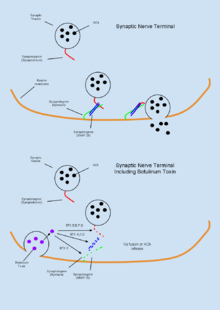
Ботулинический токсин (BTX) - это группа нейротоксинов, состоящая из восьми различных соединений, называемых BTX-A, B, C, D, E, F, G, H, которые продуцируются бактериями. Clostridium botulinum и привести к мышечной паралич.[62] Примечательно уникальной особенностью BTX является его относительно обычное терапевтическое использование при лечении дистония и спастичность расстройства,[62] а также в побуждении мышечная атрофия[11] несмотря на то, что это самое ядовитое вещество из известных.[18] BTX функционирует периферически, чтобы подавлять ацетилхолин (АЧ) выпуск на нервномышечное соединение через деградацию SNARE белки требуется для АЧ слияние пузырьков и мембран.[35] Поскольку токсин очень биологически активен, расчетной дозы в 1 мкг / кг веса тела достаточно, чтобы вызвать недостаточный дыхательный объем и, как следствие, смерть к удушье.[13] Из-за своей высокой токсичности антитоксины BTX были активной областью исследований. Было показано, что капсаицин (активное соединение, отвечающее за тепло в перцы чили ) может связать Рецептор TRPV1 выражено на холинергические нейроны и подавляют токсические эффекты BTX.[18]
Столбнячный токсин
Столбнячный нейротоксин (TeNT) - это соединение, которое функционально снижает тормозящие передачи в нервной системе, приводящие к мышечной тетании. TeNT похож на BTX и на самом деле очень похож по структуре и происхождению; оба принадлежат к одной категории клостридиальные нейротоксины.[12] Как и BTX, TeNT подавляет межнейронное взаимодействие посредством высвобождения везикулярного нейромедиатора (NT).[36] Одно заметное различие между двумя соединениями заключается в том, что хотя БТК ингибирует мышечные сокращения, TeNT их индуцирует. Хотя оба токсина ингибируют высвобождение везикул в синапсах нейронов, причина этого различного проявления заключается в том, что BTX функционирует в основном в периферической нервной системе (PNS), в то время как TeNT в значительной степени активен в Центральная нервная система (ЦНС).[63] Это результат миграции TeNT через двигательные нейроны к тормозящие нейроны спинного мозга после прохождения через эндоцитоз.[64] Это приводит к потере функции тормозных нейронов в ЦНС, что приводит к системному мышечные сокращения. Подобно прогноз смертельной дозы BTX, TeNT приводит к параличу и последующему удушье.[64]
Гематоэнцефалический барьер
Алюминий
Известно, что нейротоксическое поведение алюминия проявляется при попадании в сердечно-сосудистая система, откуда он может мигрировать в мозг и подавлять некоторые важные функции гематоэнцефалического барьера (ГЭБ).[65] Утрата функции ГЭБ может привести к значительному повреждению нейронов ЦНС, поскольку барьер, защищающий мозг от других токсинов, обнаруженных в крови, больше не будет способен к такому действию. Хотя металл известен как нейротоксичный, эффекты обычно ограничиваются пациенты неспособны удалить избыточные ионы из крови, например, те, кто испытывает почечная недостаточность.[66] Пациенты, испытывающие отравление алюминием, могут проявлять симптомы например, нарушение обучения и снижение моторная координация.[67] Кроме того, известно, что системные уровни алюминия повышаются с возрастом и, как было показано, коррелируют с Болезнь Альцгеймера, предполагая, что это нейротоксическое соединение, вызывающее заболевание.[68] Несмотря на его известную токсичность, алюминий по-прежнему широко используется при упаковке и приготовлении пищевых продуктов, в то время как другие токсичные металлы, такие как свинец, почти полностью выведены из употребления в этих отраслях промышленности.
Меркурий
Ртуть способна вызывать повреждение ЦНС, мигрируя в мозг, пересекая ГЭБ.[41] Ртуть существует в различных соединениях, хотя метилртуть (MeHg+), диметилртуть и диэтилртуть являются единственными существенно нейротоксическими формами. Диэтилртуть и диметилртуть считаются одними из самых сильнодействующих нейротоксинов, когда-либо обнаруженных.[41] MeHg+ обычно приобретается через потребление морепродукты, поскольку он имеет тенденцию концентрироваться в организмах, находящихся на вершине пищевой цепи.[69] Известно, что ион ртути ингибирует аминокислота (AA) и глутамат (Glu) транспорт, потенциально приводящий к эксайтотоксическим эффектам.[70]
Агонисты и антагонисты рецепторов
Анатоксин-а
| Внешнее видео | |
|---|---|
Ноттингемский университет |
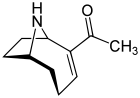
Расследование анатоксин-а, также известный как «фактор очень быстрой смерти», возник в 1961 году после смерти коров, которые пили из озера, содержащего цветение водорослей в Саскачеване, Канада.[71][72] Это цианотоксин производится как минимум четырьмя разными родами цианобактерии, и было зарегистрировано в Северной Америке, Европе, Африке, Азии и Новой Зеландии.[73]
Токсические эффекты от анатоксина-а прогрессирует очень быстро, поскольку действует непосредственно на нервные клетки (нейроны ). Прогрессирующие симптомы анатоксина-а воздействия - потеря координации, подергивание, судороги и быстрая смерть от респираторный паралич. Нервные ткани, которые сообщаются с мышцами, содержат рецептор называется никотиновый рецептор ацетилхолина. Стимуляция этих рецепторов вызывает мышечное сокращение. Анатоксин-а молекула имеет форму, соответствующую этому рецептору, и таким образом имитирует естественный нейротрансмиттер обычно используется рецептором, ацетилхолин. Как только это вызвало сокращение, анатоксин-а не позволяет нейронам вернуться в состояние покоя, потому что не деградирует холинэстераза который обычно выполняет эту функцию. В результате мышечные клетки постоянно сокращаются, связь между мозгом и мышцами нарушается, и дыхание прекращается.[74][75]
Когда он был впервые обнаружен, токсин был назван фактором очень быстрой смерти (VFDF), потому что когда он был вводится в полость тела у мышей это вызвало тремор, паралич и смерть в течение нескольких минут. В 1977 г. была определена структура VFDF как вторичная, бициклическая. амин алкалоид, и он был переименован в анатоксин-а.[76][77] Структурно он похож на кокаин.[78] Интерес к анатоксину-а из-за опасности, которую он представляет для рекреационной и питьевой воды, а также из-за того, что это особенно полезная молекула для исследования рецепторов ацетилхолина в нервной системе.[79] Смертоносность токсина означает, что он имеет высокий военный потенциал как токсинное оружие.[80]
Бунгаротоксин
Бунгаротоксин - это соединение с известным взаимодействием с никотиновые рецепторы ацетилхолина (нАХР), которые составляют семейство ионные каналы активность которого запускается связыванием нейромедиатора.[81] Бунгаротоксин производится в различных формах, хотя одной из наиболее часто используемых форм является альфа-форма с длинной цепью, α-бунгаротоксин, который изолирован от полосатая змея крайта.[37] Хотя альфа-бунгаротоксин чрезвычайно токсичен при проглатывании, он продемонстрировал широкую полезность в нейробиологии, поскольку он особенно эффективен для выделения nAChR из-за его высокого сродства к рецепторам.[37] Поскольку существует несколько форм бунгаротоксина, существуют разные формы nAChR, с которыми они будут связываться, и α-бунгаротоксин особенно специфичен для α7-нАХР.[82] Функция α7-nAChR позволяет ион кальция попадание в клетки, и, таким образом, при блокировании проглоченным бунгаротоксином будет вызывать повреждающие эффекты, поскольку передача сигналов ACh будет подавлена.[82] Точно так же использование α-бунгаротоксина может быть очень полезным в неврологии, если желательно блокировать поток кальция, чтобы изолировать эффекты других каналов. Кроме того, различные формы бунгаротоксина могут быть полезны для изучения ингибированных nAChR и возникающего в результате потока ионов кальция в различных системах организма. Например, α-бунгаротоксин специфичен для нАХР, обнаруженных в мускулатуре и κ-бунгаротоксин специфичен для nAChR, обнаруженных в нейронах.[83]
Карамбоксин
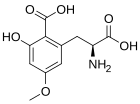
Карамбоксин (CBX) - это токсин нашел в карамбол (Averrhoa carambola). Люди с некоторыми типами заболеваний почек подвержены неблагоприятным неврологическим эффектам, включая интоксикацию, судороги и даже смерть после употребления в пищу карамболы или употребления сока из этого фрукта. Карамбоксин - это новый непептидный аминокислотный токсин, который стимулирует рецепторы глутамата в нейронах. Карамбоксин является агонистом обоих NMDA и AMPA глутаматергические ионотропные рецепторы с сильными возбуждающими, судорожными и нейродегенеративными свойствами.[84]
Кураре
Период, термин "кураре "является двусмысленным, потому что он использовался для описания ряда ядов, которые во время наименования понимались иначе, чем в настоящее время. В прошлом характеристика означала яды, используемые Южноамериканские племена на стрелки или же дартс, хотя он созрел для определения конкретной категории ядов, которые действуют на нервномышечное соединение для подавления передачи сигналов и, таким образом, для расслабления мышц.[85] Категория нейротоксинов содержит ряд различных ядов, хотя все они были первоначально очищены из растений, происходящих из Южной Америки.[85] Эффект, с которым вводится яд кураре, обычно связан с параличом мышц и, как следствие, смертью.[86] Кураре особенно действует, чтобы подавлять никотиновые рецепторы ацетилхолина на нервномышечное соединение. Обычно эти рецепторные каналы позволяют ионам натрия проникать в мышечные клетки, чтобы инициировать потенциал действия, который приводит к сокращению мышц. Блокируя рецепторы, нейротоксин способен значительно снижать передачу сигналов нервно-мышечного соединения, что привело к его использованию анестезиологи производить мышечное расслабление.[87]
Вмешательство цитоскелета
Мышьяк
Мышьяк - это нейротоксин, обычно концентрирующийся в областях, подверженных воздействию сельскохозяйственные стоки, добыча полезных ископаемых, и плавка сайты (Мартинес-Финли 2011). Одним из эффектов попадания мышьяка в организм во время развития нервной системы является угнетение нейрит рост[88] что может происходить как в ПНС, так и в ЦНС.[89] Это ингибирование роста нейритов часто может приводить к дефектам нейронная миграция, а также значительные морфологические изменения нейронов во время разработка,[90]) часто приводит к нервная трубка дефекты в новорожденные.[91] Как метаболит мышьяка, арсенит образуется после приема внутрь мышьяка и проявляет значительную токсичность для нейронов в течение примерно 24 часов после воздействия. Механизм этой цитотоксичности функционирует за счет индуцированного арсенитом увеличения внутриклеточный уровни ионов кальция в нейронах, что впоследствии может снизить трансмембранный потенциал митохондрий, который активирует каспасы, вызывая гибель клеток.[90] Другая известная функция арсенита - его деструктивная природа по отношению к цитоскелет через подавление нейрофиламент транспорт.[42] Это особенно разрушительно, так как нейрофиламенты используются в основной клеточной структуре и поддержке. Литий Однако введение показало многообещающее восстановление некоторых из утраченных подвижностей нейрофиламентов.[92] Кроме того, как и в случае с другими видами лечения нейротоксинами, введение определенных антиоксидантов показало некоторые перспективы снижения нейротоксичности проглоченного мышьяка.[90]
Аммиак

Токсичность аммиака часто проявляется двумя путями введения, либо через потребление, либо через эндогенные заболевания, такие как отказ печени.[93][94] Один примечательный случай, когда токсичность аммиака является обычным явлением, - это реакция на цирроз из печень что приводит к печеночная энцефалопатия, и может привести к отек мозга (Haussinger 2006). Этот отек мозга может быть результатом ремоделирования нервных клеток. Как следствие повышенных концентраций, активность аммиака in vivo, как было показано, вызывает набухание астроцитов в головном мозге за счет увеличения производства цГМФ (циклический гуанозинмонофосфат) внутри клеток, что приводит к Опосредованный протеинкиназой G (PKG) модификации цитоскелета.[43] Результатом этой токсичности может быть снижение энергии мозга. метаболизм и функция. Важно отметить, что токсические эффекты аммиака на ремоделирование астроцитов могут быть уменьшены путем введения L-карнитин.[93] Это ремоделирование астроцитов, по-видимому, опосредовано аммиаком. митохондриальный переход проницаемости. Этот митохондриальный переход является прямым результатом глутамин активность соединение, которое образуется из аммиака in vivo.[95] Администрация антиоксиданты или же глутаминаза ингибитор может уменьшить этот митохондриальный переход и, возможно, также ремоделирование астроцитов.[95]
Цитотоксичность, опосредованная кальцием

Свинец
Свинец - это мощный нейротоксин, токсичность которого признавалась не менее тысячи лет.[96] Хотя нейротоксические эффекты свинца обнаруживаются в обоих взрослые люди и молодой дети, развивающийся мозг особенно подвержен вредному воздействию свинца, последствия которого могут включать: апоптоз и эксайтотоксичность.[96] Основным механизмом, с помощью которого свинец может причинить вред, является его способность переноситься насосы кальциевой АТФазы через ГЭБ, обеспечивая прямой контакт с хрупкими клетками центральной нервной системы.[97] Нейротоксичность возникает из-за способности свинца действовать аналогично ионам кальция, поскольку концентрированный свинец приводит к клеточному поглощению кальция, что нарушает работу клеток. гомеостаз и вызывает апоптоз.[44] Именно это повышение внутриклеточного кальция активирует протеинкиназа C (PKC), который проявляется как нарушение обучаемости у детей в результате раннего воздействия свинца.[44] Помимо индукции апоптоза, свинец подавляет передачу сигналов между нейронами за счет нарушения высвобождения опосредованного кальцием нейромедиатора.[98]
Нейротоксины с множественными эффектами
Этиловый спирт

Было показано, что этанол как нейротоксин вызывает повреждение нервной системы и влияет на организм различными способами. Среди известных эффектов воздействия этанола есть как временные, так и долгосрочные последствия. Некоторые из длительных эффектов включают долгосрочное снижение нейрогенез в гиппокамп,[99][100] распространенная атрофия головного мозга,[101] и побудил воспаление в мозгу.[102] Следует отметить, что хроническое употребление этанола также вызывает реорганизацию компонентов клеточной мембраны, что приводит к липидный бислой отмечены повышенными мембранными концентрациями холестерин и насыщенный жир.[46] Это важно, поскольку транспорт нейротрансмиттеров может быть нарушен из-за ингибирования везикулярного транспорта, что приводит к снижению функции нейронной сети. Одним из важных примеров снижения межнейронной связи является способность этанола подавлять Рецепторы NMDA в гиппокампе, что снижает долгосрочное потенцирование (LTP) и получение памяти.[45] Было показано, что NMDA играет важную роль в LTP и, следовательно, в формировании памяти.[103] Однако при хроническом потреблении этанола чувствительность этих NMDA-рецепторов к индукции LTP увеличивается в мезолимбические дофаминовые нейроны в инозитол 1,4,5-трифосфат (IP3) зависимым образом.[104] Эта реорганизация может привести к цитотоксичности нейронов как из-за гиперактивации постсинаптических нейронов, так и из-за индуцированной зависимости от постоянного потребления этанола. Кроме того, было показано, что этанол непосредственно снижает накопление внутриклеточных ионов кальция за счет ингибирования активности рецептора NMDA и, таким образом, снижает способность к возникновению LTP.[105]
Помимо нейротоксического действия этанола на зрелые организмы, хроническое употребление в пищу способно вызвать серьезные дефекты развития. Свидетельства были впервые представлены в 1973 году о связи между хроническим потреблением этанола матерями и дефектами у их потомства.[106] Эта работа была ответственна за создание классификации алкогольный синдром плода, заболевание, характеризующееся общим морфогенез аберрации, такие как дефекты черепно-лицевой формирование, развитие конечностей и сердечно-сосудистый формирование. Величина нейротоксичности этанола у плоды было показано, что ведущий к алкогольному синдрому плода зависит от уровней антиоксидантов в мозге, таких как витамин Е.[107] Поскольку мозг плода относительно хрупок и подвержен индуцированным стрессам, тяжелые пагубные последствия воздействия алкоголя можно увидеть в таких важных областях, как гиппокамп и др. мозжечок. Тяжесть этих эффектов напрямую зависит от количества и частоты потребления этанола матерью, а также от стадии развития плода.[108] Известно, что воздействие этанола приводит к снижению уровня антиоксидантов, митохондриальной дисфункции (Chu 2007) и последующей гибели нейронов, по-видимому, в результате повышенного образования реактивные окислительные формы (ROS).[30] Это вероятный механизм, поскольку в мозге плода снижается содержание антиоксидантных ферментов, таких как каталаза и пероксидаза.[109] В поддержку этого механизма администрация высоких уровней диетический витамин E снижает или устраняет нейротоксические эффекты, вызванные этанолом, у плода.[8]
н-гексан
н-Гексан - нейротоксин, который в последние годы стал причиной отравления нескольких рабочих китайских заводов электроники.[110][111][112][113]
Рецепторно-селективные нейротоксины
MPP +
MPP +, токсичный метаболит MPTP селективный нейротоксин, который препятствует окислительного фосфорилирования в митохондрии подавляя комплекс I, что приводит к истощению АТФ и последующая гибель клеток. Это происходит почти исключительно в дофаминергических нейронах черная субстанция, в результате чего постоянные паркинсонизм у подвергшихся воздействию субъектов через 2–3 дня после администрации.
Источники эндогенных нейротоксинов
В отличие от наиболее распространенных источников нейротоксинов, которые попадают в организм при приеме внутрь, эндогенные нейротоксины как возникают, так и оказывают свое действие. in vivo. Кроме того, хотя большинство ядов и экзогенных нейротоксинов редко обладают полезными способностями in vivo, эндогенные нейротоксины обычно используются организмом полезным и здоровым образом, например оксид азота, который используется в клеточной коммуникации.[114] Часто только когда эти эндогенные соединения становятся высококонцентрированными, они приводят к опасным последствиям.[9]
Оксид азота
Хотя оксид азота (NO) обычно используется нервной системой в межнейронной коммуникации и передаче сигналов, он может быть активным в механизмах, ведущих к ишемия в головном мозге (Iadecola 1998). Нейротоксичность NO основана на его важности для эксайтотоксичности глутамата, поскольку NO вырабатывается кальций-зависимым образом в ответ на опосредованную глутаматом активацию NMDA, которая происходит с повышенной скоростью при эксайтотоксичности глутамата.[47] Хотя NO способствует усилению кровотока в потенциально ишемизированных областях мозга, он также способен увеличивать окислительный стресс,[115] индуцирование повреждения ДНК и апоптоза.[116] Таким образом, повышенное присутствие NO в ишемической области ЦНС может вызывать значительные токсические эффекты.
Глутамат
Глутамат, как и оксид азота, представляет собой эндогенно продуцируемое соединение, используемое нейронами для нормальной работы, и присутствует в небольших концентрациях на протяжении всего организма. серое вещество ЦНС.[9] Одним из наиболее заметных применений эндогенного глутамата является его функция в качестве возбуждающего нейромедиатора.[48] Однако в концентрированном состоянии глутамат становится токсичным для окружающих нейронов. Эта токсичность может быть как результатом прямой летальности глутамата для нейронов, так и результатом индуцированного притока кальция в нейроны, приводящего к отеку и некрозу.[48] Была показана поддержка этих механизмов, играющих важную роль в заболеваниях и осложнениях, таких как: болезнь Хантингтона, эпилепсия, и Инсульт.[9]
Дофамин
Дофамин представляет собой эндогенное соединение, которое используется в качестве нейротрансмиттера для модуляции ожидания вознаграждения. Дофамин убивает нейроны, производящие дофамин, вмешиваясь в электронная транспортная цепь в нейронах. Это вмешательство приводит к подавлению клеточное дыхание, что приводит к гибели нейронов.
Смотрите также
Примечания
- ^ Сивонен, К. (1999). «Токсины, вырабатываемые цианобактериями». Пузырчатый. 5: 11–18.
- ^ Правительство Шотландии Сине-зеленые водоросли (цианобактерии) во внутренних водах: оценка и контроль рисков для общественного здоровья Проверено 15 декабря 2011 года.
- ^ Медицинский словарь Дорланда для потребителей здравоохранения
- ^ а б Спенсер 2000
- ^ а б Олни 2002
- ^ а б c d е ж грамм час Кирнан 2005
- ^ Лидский 2003
- ^ а б c Хитон; Барроу, Мариета; Митчелл, Дж. Джин; Пайва, Майкл (2000). «Снижение нейротоксичности, вызванной этанолом, в центральной нервной системе новорожденных крыс с помощью антиоксидантной терапии». Алкоголизм: клинические и экспериментальные исследования. 24 (4): 512–18. Дои:10.1111 / j.1530-0277.2000.tb02019.x.
- ^ а б c d Цой 1987
- ^ Чжан 1994
- ^ а б Rosales, Raymond L .; Аримура, Кимиёси; Такенага, Сатоши; Осаме, Мицухиро (1996). «Экстрафузальные и интрафузальные мышечные эффекты в экспериментальной инъекции ботулинического токсина-А». Мышцы и нервы. 19 (4): 488–96. Дои:10.1002 / (sici) 1097-4598 (199604) 19: 4 <488 :: aid-mus9> 3.0.co; 2-8. PMID 8622728.
- ^ а б Симпсон 1986
- ^ а б c Арнон 2001
- ^ Дикранян 2001
- ^ Дэн 2003
- ^ Евтович-Тодорович 2003
- ^ Надлер 1978
- ^ а б c d Тьягараджан 2009
- ^ Нейротоксины: определение, эпидемиология, этиология
- ^ Ходж 2002
- ^ а б Доббс 2009
- ^ а б c Видмайер, Эрик П., Хершел Рафф, Кевин Т. Стренг и Артур Дж. Вандер (2008) «Физиология человека Вандера: механизмы функционирования тела». Бостон: Высшее образование Макгроу-Хилла.
- ^ а б Мартини 2009
- ^ а б Коста 2011
- ^ Гарри 1998
- ^ Gartlon 2006
- ^ Радио, Николай М .; Манди, Уильям Р. (2008). «Тестирование нейротоксичности развития in vitro: модели для оценки химического воздействия на рост нейритов». Нейротоксикология. 29 (3): 361–376. Дои:10.1016 / j.neuro.2008.02.011. PMID 18403021.
- ^ Лотти 2005
- ^ а б c Адамс 2003
- ^ а б Брокардо 2011
- ^ Левендон 2001
- ^ а б Хагдуст-Язди 2011
- ^ а б ДеБин 1993
- ^ МакКлески 1987
- ^ а б Гарсия-Родригес 2011
- ^ а б Уильямсон 1996
- ^ а б c Дутертр 2006
- ^ Коллер 1988
- ^ Рутгрере 2012
- ^ Ролик 1994
- ^ а б c Ашнер 1990
- ^ а б DeFuria 2006
- ^ а б Конопацкая 2009
- ^ а б c Бресслер 1999
- ^ а б Lovinger, D .; Белый, G .; Вес, Ф. (1989). «Этанол ингибирует NMDA-активированный ионный ток в нейронах гиппокампа». Наука. 243 (4899): 1721–724. Дои:10.1126 / science.2467382. PMID 2467382.
- ^ а б Леонард, Б. Э. (1986). «Является ли этанол нейротоксином ?: Влияние этанола на структуру и функцию нейронов». Алкоголь и алкоголизм. 21 (4): 325–38. Дои:10.1093 / oxfordjournals.alcalc.a044638.
- ^ а б Гартвейт 1988
- ^ а б c Цой 1990
- ^ Бен-Шахар Д., Зук Р., Глинка Ю. (1995). «Нейротоксичность дофамина: угнетение митохондриального дыхания». J. Neurochem. 64 (2): 718–23. Дои:10.1046 / j.1471-4159.1995.64020718.x. PMID 7830065.
- ^ а б c Чоудхури, Ф. Р .; Ахасан, Х.А. М. Назмул; Рашид, А.К. Мамунур; Мамун, А. Аль; Халидуцзаман, С. М. (2007). «Отравление тетродотоксином: клинический анализ, роль неостигмина и краткосрочные результаты в 53 случаях». Сингапурский медицинский журнал. 48 (9): 830–33. PMID 17728964.
- ^ Ахасан 2004 г.
- ^ Лау 1995
- ^ а б c Standfield 1983
- ^ Roed 1989
- ^ Хагдуст-Ясди 2011
- ^ Deshane 2003
- ^ Сорочану 1998
- ^ а б Джейкоб 2010
- ^ Оливера 1987
- ^ Круз 1986
- ^ МакКлески 1987
- ^ а б Брин, Митчелл F (1997) "Ботулинический токсин: химия, фармакология, токсичность и иммунология". Мышцы и нервы, 20 (S6): 146–68.
- ^ Монтекукко 1986
- ^ а б Пираццини 2011
- ^ Банки 1988 г.
- ^ Король 1981
- ^ Рабе 1982
- ^ Уолтон 2006
- ^ Чан 2011
- ^ Брукс 1988
- ^ Кармайкл 1978
- ^ Кармайкл 1975
- ^ Ян 2007
- ^ Дерево 2007
- ^ Национальный центр экологической оценки
- ^ Девлин 1977
- ^ Мур 1977 г.
- ^ Меткалф 2009
- ^ Стюарт 2008
- ^ Диксит 2005
- ^ Цетлин 2003
- ^ а б Лю 2008
- ^ Оттенок 2007
- ^ Garcia-Cairasco, N .; Мойсес-Нето, М .; Del Vecchio, F .; Oliveira, J.A.C .; Душ Сантуш, Ф. Л .; Castro, O.W .; Arisi, G.M .; Dantas, M. R .; Carolino, R.O.G .; Coutinho-Netto, J .; Дагостин, А. Л. А .; Родригес, М. С. А .; Leão, R.M .; Quintiliano, S.A.P .; Сильва, Л. Ф .; Гоббо-Нето, Л .; Лопес, Н. П. (2013). «Выяснение нейротоксичности звездчатого фрукта». Angewandte Chemie International Edition. 52 (49): 13067–13070. Дои:10.1002 / anie.201305382. PMID 24281890.
- ^ а б Биссет 1992
- ^ Шлезингер 1946
- ^ Гриффит, Гарольд Р .; Джонсон, Дж. Энид (1942). «Использование кураре в общей анестезии». Анестезиология. 3 (4): 418–420. Дои:10.1097/00000542-194207000-00006.
- ^ Лю 2009[требуется полная цитата ]
- ^ Вахидния 2007
- ^ а б c Роча 2011
- ^ Брендер 2005
- ^ DeFuria 2007
- ^ а б Мацуока 1991
- ^ Бузанская (2000)
- ^ а б Норенберг 2004
- ^ а б Лидски 2003
- ^ Брэдбери 1993
- ^ Лэсли 1999
- ^ Taffe 2010
- ^ Моррис 2009
- ^ Блайх 2003
- ^ Бланко 2005
- ^ Дэвис 1992
- ^ Бернье 2011
- ^ Такадера 1990
- ^ Джонс 1973
- ^ Митчелл 1999
- ^ Гиль-Мохапель 2010
- ^ Бергамини 2004
- ^ Рабочие отравились при изготовлении iPhone ABC News, 25 октября 2010 г.
- ^ Грязные секреты В архиве 2017-05-25 в Wayback Machine Иностранный корреспондент ABC, 26 октября 2010 г.
- ^ Мистер Дейзи и фабрика яблок, This American Life, 6 января 2012 г.
- ^ Руководство по безопасности и гигиене труда для н-гексана В архиве 2011-12-18 на Wayback Machine, OSHA.gov
- ^ Иадекола 1998
- ^ Бекман 1990
- ^ Bonfoco 1995
Рекомендации
- Адамс, Майкл Э .; Оливера, Бальдомеро М. (1994). «Нейротоксины: Обзор новых технологий исследования». Тенденции в неврологии. 17 (4): 151–55. Дои:10.1016/0166-2236(94)90092-2. PMID 7517594.
- Арнон, Стивен С .; Шехтер, Роберт; Inglesby, Thomas V .; Хендерсон, Дональд А .; Бартлетт, Джон Дж .; Ашер, Майкл С .; Эйцен, Эдвард; Хорошо, Энн Д .; Хауэр, Джером; Лейтон, Марсель; Лиллибридж, Скотт; Остерхольм, Майкл Т .; О'Тул, Тара; Паркер, Джеральд; Perl, Trish M .; Рассел, Филип К .; Swerdlow, Дэвид Л .; Тонат, Кевин (2001). «Ботулинический токсин как биологическое оружие». Журнал Американской медицинской ассоциации. 285 (8): 1059–069. Дои:10.1001 / jama.285.8.1059. PMID 11209178.
- Aschner, M .; Ашнер, Дж. (1990). «Нейротоксичность ртути: механизмы гематоэнцефалического барьерного транспорта». Неврология и биоповеденческие обзоры. 14 (2): 169–76. Дои:10.1016 / s0149-7634 (05) 80217-9. PMID 2190116.
- Бэнкс, Уильям А .; Кастин, Абба Дж. (1989). «Алюминий-индуцированная нейротоксичность: изменения мембранной функции на гематоэнцефалическом барьере». Неврология и биоповеденческие обзоры. 13 (1): 47–53. Дои:10.1016 / s0149-7634 (89) 80051-х. PMID 2671833.
- Баум-Байкер, Синтия (1985). «Польза умеренного потребления алкоголя для здоровья: обзор литературы». Наркотическая и алкогольная зависимость. 15 (3): 207–27. Дои:10.1016/0376-8716(85)90001-8. PMID 4028954.
- Бекман, Дж. С. (1990). «Производство видимых гидроксильных радикалов пероксинитритом: последствия для повреждения эндотелия из-за оксида азота и супероксида». Труды Национальной академии наук. 87 (4): 1620–624. Дои:10.1073 / pnas.87.4.1620. ЧВК 53527. PMID 2154753.
- Бергамини, Карло М .; Гамбетти, Стефани; Донди, Алессия; Червеллати, Карло (2004). «Кислород, активные формы кислорода и повреждение тканей». Текущий фармацевтический дизайн. 10 (14): 1611–626. Дои:10.2174/1381612043384664. PMID 15134560.
- Бернье Брайан Э., Уитакер Лесли Р., Морикава Хитоши (2011). «Предыдущий опыт использования этанола усиливает синаптическую пластичность рецепторов NMDA в вентральной тегментальной области». Журнал неврологии. 31 (14): 5305–212. Дои:10.1523 / JNEUROSCI.5282-10.2011. ЧВК 3086894. PMID 21471355.CS1 maint: несколько имен: список авторов (связь)
- Биссет, Норман Дж. (1992). «Войны и охотничьи яды Нового Света. Часть 1. Заметки по ранней истории Кураре». Журнал этнофармакологии. 36 (1): 1–26. Дои:10.1016 / 0378-8741 (92) 90056-в.. PMID 1501489.
- Бланко, Ана М .; Valles, Soraya L .; Паскуаль, Мария; Герри, Консуэло (2005). «Вовлечение передачи сигналов рецептора TLR4 / IL-1 типа I в индукцию медиаторов воспаления и гибель клеток, вызванную этанолом в культивируемых астроцитах». Журнал иммунологии. 175 (10): 6893–899. Дои:10.4049 / jimmunol.175.10.6893. PMID 16272348.
- Блайх, S (2003). «Гипергомоцистеинемия как новый фактор риска уменьшения размера мозга у больных алкоголизмом». Письма о неврологии. 335 (3): 179–82. Дои:10.1016 / s0304-3940 (02) 01194-1. PMID 12531462.
- Бонфоко Э (1995). «Апоптоз и некроз: два различных события, вызванных, соответственно, легким и интенсивным воздействием N-метил-D-аспартата или оксида азота / супероксида в культурах корковых клеток». Труды Национальной академии наук. 92 (16): 7162–166. Дои:10.1073 / пнас.92.16.7162. ЧВК 41299. PMID 7638161.
- Брэдбери, штат Массачусетс; Дин, Р. (1993). «Проницаемость гематоэнцефалического барьера для свинца. [Обзор]». Нейротоксикология. 14 (2–3): 131–6. PMID 8247388.
- Brender, J .; Suarez, L .; Felkner, M .; Gilani, Z .; Stinchcomb, D .; Муди, К .; Генри, Дж .; Хендрикс, К. (2006). «Материнское воздействие мышьяка, кадмия, свинца и ртути, а также дефекты нервной трубки у потомства». Экологические исследования. 101 (1): 132–39. Дои:10.1016 / j.envres.2005.08.003. PMID 16171797.
- Бресслер, Дж; Kim, KA; Чакраборти, Т; Гольдштейн, Г. (1999). «Молекулярные механизмы нейротоксичности свинца. [Обзор]». Neurochem Res. 24 (4): 595–600. Дои:10.1023 / А: 1022596115897. PMID 10227691.
- Brocardo, Patricia S .; Гиль-Мохапель, Джоана; Кристи, Брайан Р. (2011). «Роль окислительного стресса в расстройствах алкогольного спектра плода». Обзоры исследований мозга. 67 (1–2): 209–25. Дои:10.1016 / j.brainresrev.2011.02.001. PMID 21315761.
- Брукс, Н. (1988). «Специфичность и обратимость ингибирования HgCl транспорта глутамата в культурах астроцитов». Журнал нейрохимии. 50 (4): 1117–122. Дои:10.1111 / j.1471-4159.1988.tb10581.x. PMID 2894409.
- Бузанская, Л .; Заблока, Б .; Dybel, A .; Доманская-Яник, К .; Альбрехт, Дж. (2000). «Задержка индукции апоптоза аммиаком в клетках глиомы C6». Neurochemistry International. 37 (2–3): 287–97. Дои:10.1016 / s0197-0186 (00) 00030-9. PMID 10812214.
- Кармайкл У.В., Биггс Д.Ф., Горхэм PR (1975). «Токсикология и фармакологическое действие токсина Anabaena flos-aquae». Наука. 187 (4176): 542–544. Дои:10.1126 / science.803708. PMID 803708.
- Кармайкл У.В., Горэм PR (1978). «Анатоксины из клонов Anabaena flos-aquae, выделенных из озер западной Канады». Mitt. Адский. Verein. Лимнол. 21: 285–295.
- Чан, Х. М. (2011) «Ртуть в рыбе: риски для здоровья человека». Энциклопедия гигиены окружающей среды: 697–704.
- Чой, Д. (1988). «Опосредованная кальцием нейротоксичность: связь с конкретными типами каналов и роль в ишемическом повреждении». Тенденции в неврологии. 11 (10): 465–69. Дои:10.1016/0166-2236(88)90200-7. PMID 2469166.
- Choi, D. W .; Ротман, С. М. (1990). «Роль нейротоксичности глутамата в гипоксически-ишемической смерти нейронов». Ежегодный обзор нейробиологии. 13 (1): 171–82. Дои:10.1146 / annurev.neuro.13.1.171.
- Чой, Деннис В. (1987). «Ионная зависимость нейротоксичности глутамата». Журнал неврологии. 7 (2): 369–79. Дои:10.1523 / JNEUROSCI.07-02-00369.1987.
- Чой, Деннис В .; Маулуччи-Гедде, Маргарет; Кригштейн, Арнольд Р. (1987). «Нейротоксичность глутамата в культуре клеток коры». Журнал неврологии. 7 (2): 357–68. Дои:10.1523 / JNEUROSCI.07-02-00357.1987.
- Чу, Дженнифер; Тонг, Мин; Монте, Сюзанна М. (2007). «Хроническое воздействие этанола вызывает дисфункцию митохондрий и окислительный стресс в незрелых нейронах центральной нервной системы». Acta Neuropathologica. 113 (6): 659–73. Дои:10.1007 / s00401-007-0199-4. PMID 17431646.
- Клэнси, Барбара; Finlay, Barbara L .; Дарлингтон, Ричард Б .; Ананд, К.Дж.С. (2007). «Экстраполяция развития мозга экспериментальных видов на людей». Нейротоксикология. 28 (5): 931–37. Дои:10.1016 / j.neuro.2007.01.014. ЧВК 2077812. PMID 17368774.
- Коста, Лучио Дж., Дженнаро Джордано и Марина Гицзетти (2011) Нейротоксикология in vitro: методы и протоколы. Нью-Йорк: Humana.
- Койл, Джозеф Т .; Шварц, Роберт. «Поражение стриатальных нейронов каиновой кислотой обеспечивает модель хореи Хантингтона». Природа. 246: 244–46.
- Cruz, Lourdes J .; Оливера, Бальдомеро М. (1987). «Антагонисты кальциевых каналов ω-конотоксин определяет новый сайт с высоким сродством». Журнал биологической химии. 14 (261): 6230–233.
- Davis, S .; Butcher, S.P .; Моррис Р. (1992). «Антагонист рецепторов NMDA D-2-амино-5-фосфонопентаноат (D-AP5) ухудшает пространственное обучение и LTP in vivo при внутримозговых концентрациях, сопоставимых с концентрациями, которые блокируют LTP in vitro». Журнал неврологии. 12 (1): 21–34. Дои:10.1523 / JNEUROSCI.12-01-00021.1992.
- Доусон, В. Л. (1991). «Оксид азота опосредует нейротоксичность глутамата в первичных корковых культурах». Труды Национальной академии наук. 88 (14): 6368–371. Дои:10.1073 / pnas.88.14.6368. ЧВК 52084. PMID 1648740.
- Дебин, Джон А., Джон Э. Маджо и Гэри Р. Стрихарц (1993) "Очистка и характеристика хлоротоксина, лиганда хлоридного канала из яда скорпиона". Американское физиологическое обществоС. 361–69.
- ДеФурия, Джейсон; Ши, Томас Б. (2007). «Мышьяк ингибирует транспорт нейрофиламентов и индуцирует накопление фосфорилированных нейрофиламентов в перикариале: роль JNK и GSK-3β». Исследование мозга. 1181: 74–82. Дои:10.1016 / j.brainres.2007.04.019. PMID 17961518.
- Дефурия, Джейсон (2006) "Нейротоксин окружающей среды, мышьяк, влияет на динамику нейрофиламентов за счет сверхактивации терминальной киназы C-JUN: потенциальная роль в боковом амиотрофическом склерозе". UMI, стр. 1–16.
- Дэн Вэньбинь, Порец Рональд Д. (2003). «Олигодендроглии в нейротоксичности развития». Нейротоксикология. 24 (2): 161–78. Дои:10.1016 / s0161-813x (02) 00196-1. PMID 12606289.
- Дешейн, Джесси; Гарнер, Крейг С.; Sontheimer, Харальд (2003). «Хлоротоксин ингибирует инвазию клеток глиомы через матриксную металлопротеиназу-2». Журнал биологической химии. 278 (6): 4135–144. Дои:10.1074 / jbc.m205662200. PMID 12454020.
- Девлин Дж. П., Эдвардс О. Е., Горхэм ПР, Хантер Н. Р., Пайк Р. К., Ставрик Б. (1977). «Анатоксин-а, токсичный алкалоид из Anabaena flos-aquae NRC-44h». Может. J. Chem. 55 (8): 1367–1371. Дои:10.1139 / v77-189. Архивировано из оригинал на 2012-07-09.
- Дикранян, К (2001). «Апоптоз в переднем мозге млекопитающих in vivo». Нейробиология болезней. 8 (3): 359–79. Дои:10.1006 / nbdi.2001.0411. PMID 11447994.
- Диксит А., Дакед Р.К., Алам С.И., Сингх Л. (2005). «Военный потенциал биологических нейротоксинов». Отзывы о токсинах. 24 (2): 175–207. Дои:10.1081 / TXR-200057850.
- Доббс, Майкл Р. (2009) Клиническая нейротоксикология. Филадельфия: Сондерс-Эльзевьер.
- Dutertre, S .; Льюис, Р. (2006). «Токсины в никотиновых рецепторах ацетилхолина». Биохимическая фармакология. 72 (6): 661–70. Дои:10.1016 / j.bcp.2006.03.027. PMID 16716265.
- Garcia-Rodriguez, C .; Герен, И. Н .; Lou, J .; Конрад, Ф .; Forsyth, C .; Wen, W .; Chakraborti, S .; Zao, H .; Manzanarez, G .; Smith, T.J .; Brown, J .; Tepp, W. H .; Liu, N .; Wijesuriya, S .; Tomic, M. T .; Johnson, E. A .; Smith, L.A .; Маркс, Дж. Д. (2011). «Response Re:« Нейтрализация человеческих моноклональных антител, связывающих множественные серотипы нейротоксина ботулина », Гарсия-Родригес и др., PEDS, 2011; 24: 321–331». Разработка и отбор протеинов. 24 (9): 633–34. Дои:10.1093 / белок / gzr012.
- Гартуэйт, Джон; Чарльз, Сара Л .; Чесс-Уильямс, Рассел (1988). «Производное эндотелима высвобождение расслабляющего фактора при активации рецепторов NMDA предполагает роль межклеточного посредника в мозге». Природа. 336 (24): 385–88. Дои:10.1038 / 336385a0. PMID 2904125.
- Gartlon, J .; Кинснер, А .; Balprice, A .; Coecke, S .; Clothier, Р. (2006). «Оценка предлагаемой стратегии тестирования in vitro с использованием нейрональных и ненейрональных клеточных систем для обнаружения нейротоксичности». Токсикология in vitro. 20 (8): 1569–581. Дои:10.1016 / j.tiv.2006.07.009. PMID 16959468.
- Гиль-Мохапель, Джоана; Беме, Фанни; Кайнер, Лия; Кристи, Брайан Р. (2010). «Потеря клеток гиппокампа и нейрогенез после воздействия алкоголя на плод: выводы из различных моделей грызунов». Обзоры исследований мозга. 64 (2): 283–303. Дои:10.1016 / j.brainresrev.2010.04.011. PMID 20471420.
- Хагдуст-Язди, Хашем; Фараджи, Айда; Фрайдуни, Негин; Мовахеди, Мохадесе; Хадибейги, Эльхам; Ваези, Фатемех (2011). «Значительные эффекты 4-аминопиридина и тетраэтиламмония в лечении болезни Паркинсона, вызванной 6-гидроксидофамином». Поведенческие исследования мозга. 223 (1): 70–74. Дои:10.1016 / j.bbr.2011.04.021. PMID 21540059.
- Гарри, Г. Дж .; Биллингсли, Мелвин; Бруининк, Арендд; Кэмпбелл, Иэн Л .; Классен, Вернер; Дорман, Дэвид С .; Галли, Коррадо; Рэй, Дэвид; Смит, Роберт А .; Тилсон, Хью А. (1998). «Методы in vitro для оценки нейротоксичности». Перспективы гигиены окружающей среды. 106 (Дополнение 1): 131–58. Дои:10.2307/3433917. JSTOR 3433917. ЧВК 1533280. PMID 9539010.
- Häussinger, Дитер (2006). «Отек мозга низкой степени и патогенез печеночной энцефалопатии при циррозе». Гепатология. 43 (6): 1187–190. Дои:10.1002 / hep.21235. PMID 16729329.
- Хенсли, К. (1994). «Модель агрегации β-амилоида и нейротоксичности на основе образования свободных радикалов пептидом: отношение к болезни Альцгеймера». Труды Национальной академии наук. 91 (8): 3270–274. Дои:10.1073 / пнас.91.8.3270. ЧВК 43558. PMID 8159737.
- Герберт, М. Р. (2006) "Аутизм и экологическая геномика". НейротоксикологияС. 671–84. Интернет.
- Ходж, А. Тревор (2002) Римские акведуки и водоснабжение. Лондон: Дакворт.
- Как, C (2003). «Отравление тетродотоксином». Американский журнал неотложной медицины. 21 (1): 51–54. Дои:10.1053 / ajem.2003.50008. PMID 12563582.
- Хюэ, Бернард; Букингем, Стивен Д .; Букингем, Дэвид; Sattelle, Дэвид Б. (2007). «Действия змеиного нейротоксина на никотиновый холинергический синапс насекомых». Неврология беспозвоночных. 7 (3): 173–78. Дои:10.1007 / s10158-007-0053-3. PMID 17710455.
- Иадекола Константино (1997). «Яркие и темные стороны оксида азота при ишемической травме головного мозга». Тенденции в неврологии. 20 (3): 132–39. Дои:10.1016 / s0166-2236 (96) 10074-6. PMID 9061868.
- Джейкоб, Рид Б .; Макдугал, Оуэн М. (2010). «М-суперсемейство конотоксинов: обзор». Клеточные и молекулярные науки о жизни. 67 (1): 17–27. Дои:10.1007 / s00018-009-0125-0. ЧВК 3741454. PMID 19705062.
- Евтович-Тодорович, Весна; Хартман, Ричард Э .; Изуми, Юкитоши; Беншофф, Николас Д .; Дикранян, Крикор; Зорумски, Чарльз Ф .; Олни, Джон В .; Возняк, Дэвид Ф. (2003). «Раннее воздействие общих анестетиков вызывает повсеместную нейродегенерацию в развивающемся мозге крыс и стойкие нарушения обучения». Журнал неврологии. 23 (3): 876–82. Дои:10.1523 / JNEUROSCI.23-03-00876.2003.
- Джонс, К. (1973). "Образец уродства в потомстве матерей-хронических алкоголиков". Ланцет. 301 (7815): 1267–271. Дои:10.1016 / с0140-6736 (73) 91291-9. PMID 4126070.
- Кирнан, Мэтью С .; Isbister, Джеффри К .; Синди; Lin, S.-Y .; Берк, Дэвид; Босток, Хью (2005). «Острая нейротоксичность, вызванная тетродотоксином, после проглатывания фугу». Анналы неврологии. 57 (3): 339–48. Дои:10.1002 / ana.20395. PMID 15732107.
- Кинг, Стивен У .; Чабер, Джон; Уиллс, Майкл Р .; Гительман, Х. Дж. (1981). «Клиническая биохимия алюминия». Критические обзоры в клинических лабораторных науках. 14 (1): 1–20. Дои:10.3109/10408368109105861. PMID 7016437.
- Конопацкая, Агнешка; Konopacki, Filip A .; Альбрехт, янв (2009). «Протеинкиназа G участвует в набухании астроцитов, вызванном аммиаком». Журнал нейрохимии. 109: 246–51. Дои:10.1111 / j.1471-4159.2009.05802.x. PMID 19393034.
- Лафон-Казаль, Мирей; Пьетри, Сильвия; Кулькаси, Марсель; Бокерт, Джоэл (1993). «NMDA-зависимое производство супероксида и нейротоксичность». Природа. 364 (6437): 535–37. Дои:10.1038 / 364535a0. PMID 7687749.
- Ласли, С.М.; Грин, МС; Гилберт, ME (1999). «Влияние периода воздействия на глутамат гиппокампа in vivo и высвобождение ГАМК у крыс, хронически подвергающихся воздействию свинца». Нейротоксикология. 20 (4): 619–29. PMID 10499360.
- Lau, F. L .; Wong, C.K .; Ип, С. Х. (1995). «Отравление рыбой фугу». Журнал неотложной медицины. 12 (3): 214–15. Дои:10.1136 / emj.12.3.214. ЧВК 1342486. PMID 8581253.
- Lewendon, G .; Kinra, S .; Nelder, R .; Кронин, Т. (2001). «Следует ли регулярно проверять детей с проблемами развития и поведения на наличие свинца?». Архив детских болезней. 85 (4): 286–88. Дои:10.1136 / adc.85.4.286. ЧВК 1718950. PMID 11567935.
- Лидский Теодор I (2003). «Нейротоксичность свинца у детей: основные механизмы и клинические корреляты». Мозг. 126 (1): 5–19. Дои:10.1093 / мозг / awg014. PMID 12477693.
- Лю, Куанг-Кай; Чен, Мэй-Фанг; Чен, По-И; Ли, Тони Дж. Ф .; Ченг, Чиа-Лян; Чанг, Чиа-Цзин; Хо, Йен-Пэн; Жуй-И, Чао (2008). «Связывание альфа-бунгаротоксина с клеткой-мишенью в развивающейся зрительной системе карбоксилированным наноалмазом». Нанотехнологии. 19 (20): 205102. Дои:10.1088/0957-4484/19/20/205102. PMID 21825732.
- Лю, Юань; Макдермотт, Сюзанна; Лоусон, Эндрю; Элион, К. Марджори (2010). «Взаимосвязь между умственной отсталостью и задержкой развития у детей и уровнями мышьяка, ртути и свинца в образцах почвы, взятых возле места жительства их матери во время беременности». Международный журнал гигиены и гигиены окружающей среды. 213 (2): 116–23. Дои:10.1016 / j.ijheh.2009.12.004. ЧВК 2836425. PMID 20045663.
- Лоренцо, А (1994). «Нейротоксичность β-амилоида требует образования фибрилл и ингибируется Конго Красным». Труды Национальной академии наук. 91 (25): 12243–2247. Дои:10.1073 / пнас.91.25.12243. ЧВК 45413. PMID 7991613.
- Лотти, Марчелло и Анджело Моретто (1989) «Задержанная полинейропатия, индуцированная фосфорорганическими соединениями». Токсикологические обзоры, 24 (1) (2005): 37–49.
- Мартинес-Финли, Эбани Дж .; Гоггин, Саманта Л .; Лабрек, Мэтью Т .; Аллан, Андреа М. (2011). «Снижение экспрессии генов MAPK / ERK в потомстве, подвергающемся воздействию мышьяка в перинатальном периоде, вызванное дефицитом рецепторов глюкокортикоидов». Нейротоксикология и тератология. 33 (5): 530–37. Дои:10.1016 / j.ntt.2011.07.003. ЧВК 3183307. PMID 21784148.
- Мартини, Фредерик, Майкл Дж. Тиммонс и Роберт Б. Таллитч (2009) Анатомия человека. Сан-Франциско: Пирсон / Бенджамин Каммингс.
- Мацуока, Масато; Игису, Хидеки; Кохрияма, Кадзуаки; Иноуэ, Наохиде (1991). «Подавление нейротоксичности аммиака L-карнитином». Исследование мозга. 567 (2): 328–31. Дои:10.1016 / 0006-8993 (91) 90814-с. PMID 1817738.
- МакКлески, Э. У. (1987). «Омега-конотоксин: прямая и стойкая блокада определенных типов кальциевых каналов в нейронах, но не в мышцах». Труды Национальной академии наук. 84 (12): 4327–331. Дои:10.1073 / pnas.84.12.4327. ЧВК 305078. PMID 2438698.
- Meldrum, B .; Гартуэйт, Дж. (1990). «Возбуждающая нейротоксичность аминокислот и нейродегенеративное заболевание». Тенденции в фармакологических науках. 11 (9): 379–87. Дои:10.1016 / 0165-6147 (90) 90184-а. PMID 2238094.
- Меткалф, Джеймс С .; Кодд, Джеффри А. (2009). «Цианобактерии, нейротоксины и водные ресурсы: есть ли последствия для нейродегенеративных заболеваний человека?». Боковой амиотрофический склероз. 10: 74–78. Дои:10.3109/17482960903272942. PMID 19929737.
- Митчелл, Дж. Джин; Пайва, Майкл; Барроу Хитон, Мариета (1999). «Антиоксиданты, витамин Е и β-каротин, защищают от нейротоксичности, вызванной этанолом, в культурах эмбрионального гиппокампа крыс». Алкоголь. 17 (2): 163–68. Дои:10.1016 / s0741-8329 (98) 00051-2. PMID 10064385.
- Мур RE (1977). «Токсины сине-зеленых водорослей». Бионаука. 27 (12): 797–802. Дои:10.2307/1297756. JSTOR 1297756.
- Монтекукко C (1986). «Как столбняк и ботулотоксины связываются с нейронными мембранами?». Тенденции в биохимических науках. 11 (8): 314–17. Дои:10.1016/0968-0004(86)90282-3.
- Моррис, Стефани А., Дэвид В. Ивс, Александр Р. Смит и Кимберли Никсон (2009) «Алкогольное ингибирование нейрогенеза: механизм нейродегенерации гиппокампа в модели злоупотребления алкоголем в подростковом возрасте». Гиппокамп: нет данных.
- Надлер; Victor, J .; Перри, Брюс У .; Котман, Карл В. (1978). «Внутрижелудочковая каиновая кислота преимущественно разрушает пирамидные клетки гиппокампа». Природа. 271 (5646): 676–77. Дои:10.1038 / 271676a0. PMID 625338.
- Национальный центр экологической оценки (2006) «Токсикологические обзоры токсинов цианобактерий: анатоксин-а» NCEA-C-1743
- Норенберг, М. Д .; Рао, К. В. Рама; Джаякумар, А. Р. (2004). «Нейротоксичность аммиака и переход митохондриальной проницаемости». Журнал биоэнергетики и биомембран. 36 (4): 303–07. Дои:10.1023 / б: jobb.0000041758.20071.19. PMID 15377862.
- Olivera, Baldomero M .; Cruz, Lourdes J .; Де Сантос, Виктория; ЛеЧеминант, Гарт; Гриффин, Дэвид; Зейкус, Регина; Макинтош, Дж. Майкл; Галиан, Роберт; Варга, Янош (1987). «Нейрональные антагонисты кальциевых каналов. Различение подтипов кальциевых каналов с использованием омега-конотоксина из яда Conus Magus». Биохимия. 26 (8): 2086–090. Дои:10.1021 / bi00382a004. PMID 2441741.
- Олни, Джон В. (2002). «Новые идеи и новые проблемы в нейротоксикологии развития». Нейротоксикология. 23 (6): 659–68. Дои:10.1016 / s0161-813x (01) 00092-4. PMID 12520755.
- Пираццини, Марко, Орнелла Россетто, Паоло Болоньезе, Клиффорд С. Шон и Чезаре Монтекукко (2011) «Двойное крепление к мембране и неповрежденная межцепочечная дисульфидная связь необходимы для индуцированного низким PH входа нейротоксинов столбняка и ботулотоксина в нейроны. " Клеточная микробиология: № Печ.
- Рабе, Аусма; Хэ Ли, Мун; Шек, Джуди; Вишневский, Хенрик М. (1982). «Дефицит обучения у незрелых кроликов с нейрофибриллярными изменениями, вызванными алюминием». Экспериментальная неврология. 76 (2): 441–46. Дои:10.1016/0014-4886(82)90220-5. PMID 6896495.
- Rocha, R.A .; Gimeno-Alcaniz, J. V .; Мартин-Ибанес, Раймонд; Canals, J.M .; Vélez, D .; Девеса, В. (2011). «Мышьяк и фторид вызывают апоптоз нейральных клеток-предшественников». Письма токсикологии. 203 (3): 237–44. Дои:10.1016 / j.toxlet.2011.03.023. PMID 21439358.
- Роед, А (1989). «Эффекты тетраэтиламмония во время подергивания и тетанической стимуляции подготовки диафрагмы диафрагмального нерва у крысы». Нейрофармакология. 28 (6): 585–92. Дои:10.1016/0028-3908(89)90137-8. PMID 2547180.
- Rothman, S .; Thurston, J .; Хаухарт Р. (1987). «Отсроченная нейротоксичность возбуждающих аминокислот in vitro». Неврология. 22 (2): 471–80. Дои:10.1016/0306-4522(87)90347-2. PMID 3670595.
- Шлезингер, Эдвард Б. (1946). "Curare Обзор его терапевтических эффектов и их физиологической основы". Американский журнал медицины. 1 (5): 518–30. Дои:10.1016/0002-9343(46)90073-3. PMID 21001466.
- Зиблер, М .; Koller, H .; Schmalenbach, C .; Мюллер, Х. (1988). «ГАМК-активированные хлоридные токи в культивируемых нейронах гиппокампа и перегородки крыс могут быть ингибированы кураре и атропином». Письма о неврологии. 93 (2–3): 220–24. Дои:10.1016/0304-3940(88)90085-7. PMID 2468110.
- Симпсон, Л. Л. (1986). «Молекулярная фармакология ботулотоксина и столбнячного токсина». Ежегодный обзор фармакологии и токсикологии. 26 (1): 427–53. Дои:10.1146 / annurev.pharmtox.26.1.427.
- Сорочану, Лилиана; Гиллеспи, Янси; Khazaeli, M. B .; Зонтхаймер, Гарольд (1998). «Использование хлортоксина для лечения первичных опухолей головного мозга». Исследования рака. 58 (21): 4871–879. PMID 9809993.
- Спенсер PS, Schaumburg HH, Ludolph AC (Eds) (2000) Экспериментальная и клиническая нейротоксикология. Издательство Оксфордского университета, Оксфорд, стр. 1310.
- Стэнфилд, Питер Р. (1983). «Ионы тетраэтиламмония и калиевая проницаемость возбудимых клеток». Обзоры физиологии, биохимии и фармакологии. 97: 1–49. Дои:10.1007 / bfb0035345. ISBN 978-3-540-11701-8. PMID 6306751.
- Стюарт I, Сиврайт А.А., Шоу Г.Р. (2008). «Цианобактериальное отравление домашнего скота, диких млекопитающих и птиц - обзор» (PDF). Вредное цветение водорослей цианобактериями: состояние науки и исследовательские потребности. Успехи экспериментальной медицины и биологии. 619: 613–637. Дои:10.1007/978-0-387-75865-7_28. ISBN 978-0-387-75864-0. PMID 18461786.
- Taffe, M. A .; Kotzebue, R.W .; Crean, R.D .; Crawford, E. F .; Эдвардс, S .; Мандьям, К. Д. (2010). «От обложки: длительное снижение нейрогенеза гиппокампа за счет употребления алкоголя у приматов-подростков». Труды Национальной академии наук. 107 (24): 11104–1109. Дои:10.1073 / pnas.0912810107. ЧВК 2890755. PMID 20534463.
- Такадера, Цунео; Сузуки, Риса; Мохри, Тетсуро (1990). «Защита этанолом корковых нейронов от нейротоксичности, вызванной N-метил-d-аспартатом, связана с блокированием притока кальция». Исследование мозга. 537 (1–2): 109–14. Дои:10.1016 / 0006-8993 (90) 90346-д. PMID 1982237.
- Thyagarajan, B .; Кривицкая, Н .; Potian, J. G .; Hognason, K .; Garcia, C.C .; Макардл, Дж. Дж. (2009). «Капсаицин защищает нервно-мышечные соединения мышей от нейропаралитического действия ботулинического нейротоксина А.» Журнал фармакологии и экспериментальной терапии. 331 (2): 361–71. Дои:10.1124 / jpet.109.156901. ЧВК 2775269. PMID 19654265.
- Тимянски, Михаил; Charlton, Milton P .; Карлен, Питер Л .; Татор, Чарльз Х. (2003). «Исходная специфичность ранней нейротоксичности кальция в культивируемых эмбриональных спинномозговых нейронах». Журнал неврологии. 13 (5): 2095–104. ЧВК 6576557. PMID 8097530.
- Цетлин В.И.; Хучо, Ф. (2004). «Токсины змеи и улитки, действующие на никотиновые рецепторы ацетилхолина: фундаментальные аспекты и медицинское применение». Письма FEBS. 557 (1–3): 9–13. Дои:10.1016 / s0014-5793 (03) 01454-6. PMID 14741333.
- USEPA (Агентство по охране окружающей среды США) (1998) Руководства по испытаниям воздействия на здоровье. ОППТС 870.6200. Батарея для скрининга нейротоксичности. Вашингтон, округ Колумбия, USEPA.
- Вахидния, А., Г. Б. Ван дер Воет и Ф.А. де Вольф (2007) "Обзор нейротоксичности мышьяка". Человек и экспериментальная токсикология, 26 (10) : 823–32.
- Уолтон, Дж (2006). «Алюминий в нейронах гиппокампа людей с болезнью Альцгеймера». Нейротоксикология. 27 (3): 385–94. Дои:10.1016 / j.neuro.2005.11.007. PMID 16458972.
- Видмайер, Эрик П., Хершел Рафф, Кевин Т. Стрэнг и Артур Дж. Вандер (2008) Физиология человека Вандера: механизмы функционирования тела ». Бостон: Высшее образование Макгроу-Хилла.
- Уильямсон, Lura C .; Halpern, Jane L .; Монтекукко, Чезаре; Brown, J. E .; Нил, Элейн А. (1996). «Клостридиальные нейротоксины и субстратный протеолиз в интактных нейронах». Журнал биологической химии. 271 (13): 7694–699. Дои:10.1074 / jbc.271.13.7694. PMID 8631808.
- Вуд С. А .; Расмуссен Дж. П .; Холланд П. Т .; Кэмпбелл Р .; Кроу А. Л. М. (2007). «Первый отчет цианотоксина анатоксина-А из Aphanizomenon issatschenkoi (цианобактерии)». Журнал психологии. 43 (2): 356–365. Дои:10.1111 / j.1529-8817.2007.00318.x.
- Ямада, Киёфуми; Танака, Томоко; Хан, Дайкен; Сендзаки, Кодзи; Камеяма, Цутому; Набешима, Тоситака (1999). «Защитные эффекты идебенона и α-токоферола на β-амилоид- (1–42) -индуцированные дефициты обучения и памяти у крыс: влияние окислительного стресса на β-амилоид-индуцированную нейротоксичность in vivo». Европейский журнал нейробиологии. 11 (1): 83–90. Дои:10.1046 / j.1460-9568.1999.00408.x. PMID 9987013.
- Ян; Ду, Ши; Чен, Си; Фу, Джин; Чен, Мин; Чжу, Хуайцзе; Рохер, Алекс; Слэттери, Тимоти; Чжао, Лэй; Нагашима, Марико; Морсер, Джон; Мигели, Антонио; Наврот, Питер; Стерн, Дэвид; Мари Шмидт, Энн (1996). «Нейротоксичность RAGE и амилоид-β-пептидов при болезни Альцгеймера». Природа. 382 (6593): 685–91. Дои:10.1038 / 382685a0. PMID 8751438.
- Ян, Икс (2007) Распространение цианобактериального нейротоксина анатоксина-а в водах штата Нью-Йорк ProQuest. ISBN 978-0-549-35451-2.
- Zhang, J .; Dawson, V .; Dawson, T .; Снайдер, С. (1994). «Активация оксидом азота поли (АДФ-рибоза) синтетазы при нейротоксичности». Наука. 263 (5147): 687–89. Дои:10.1126 / science.8080500. PMID 8080500.
дальнейшее чтение
- Книга фактов о мозге в Обществе неврологии
- Тексты неврологии в Медицинской школе Техасского университета
- Нейротоксикология in vitro: введение в Springerlink
- Биология рецептора NMDA в NCBI
- Достижения в неврологии зависимости, 2-е издание в NCBI
внешняя ссылка
- Агентство по охране окружающей среды в Агентстве по охране окружающей среды США
- Алкоголь и алкоголизм в Oxford Medical Journals
- Нейротоксикология в журналах Elsevier
- Институт нейротоксинов в Институте нейротоксинов
- [постоянная мертвая ссылка ] Нейротоксины в Toxipedia
