Половой отбор - Sexual selection


Половой отбор это режим естественный отбор в котором члены одного биологический пол выбрать товарищей другого секс к приятель с (интерсексуальный отбор) и соревноваться с представителями того же пола за доступ к представителям противоположного пола (интрасексуальный отбор). Эти две формы отбора означают, что у некоторых особей больше репродуктивный успех чем другие в численность населения, например, потому что они более привлекательны или предпочитают более привлекательных партнеров для производства потомство.[1][2] Например, в период размножения половой отбор у лягушек происходит, когда самцы сначала собираются у кромки воды и делают свои брачный зов: карканье. Затем прибывают самки и выбирают самцов с самым глубоким карканьем и лучшими территориями. В целом, самцы получают выгоду от частых спариваний и монополизации доступа к группе фертильных самок. Самки могут иметь ограниченное количество потомков и получать максимальную отдачу от энергии, которую они вкладывают в воспроизводство.
Концепция была впервые сформулирована Чарльз Дарвин и Альфред Рассел Уоллес кто описал это как движущую силу адаптации видов и что многие организмы развили черты, функции которых пагубно сказываются на их индивидуальном выживании,[3] а затем разработан Рональд Фишер в начале 20 века. Половой отбор может привести к тому, что самцы будут прилагать все усилия, чтобы продемонстрировать свое фитнес быть выбранными женщинами, производящими половой диморфизм в вторичные половые признаки, например, богато украшенный оперение из птицы Такие как райские птицы и павлин, или рога олень, или гривы львы, вызванный положительный отзыв механизм, известный как Беглый рыбак, где передача стремления к какой-либо черте одного пола так же важна, как и наличие этой черты у другого пола для создания эффекта бегства. Хотя гипотеза сексуального сына указывает на то, что самки предпочтут потомство мужского пола, Принцип фишера объясняет, почему соотношение полов чаще всего 1: 1. Половой отбор также встречается у растений и грибы.[4][5]
История
Дарвин
Половой отбор был впервые предложен Чарльзом Дарвином в Происхождение видов (1859 г.) и развит в Происхождение человека и отбор в отношении пола (1871), поскольку он считал, что один лишь естественный отбор не может объяснить определенные типы адаптации, не связанной с выживанием. Однажды он написал своему коллеге, что «вид пера в павлин от хвоста, когда я смотрю на него, меня тошнит! »Его работа разделила половой отбор на соревнование между мужчинами и выбор женщин.
... зависит не от борьбы за существование, а от борьбы между самцами за обладание самками; результат - не смерть неудачливого конкурента, а небольшое количество потомства или его отсутствие.[6]
... когда самцы и самки любого животного имеют одинаковые общие повадки ... но отличаются по строению, цвету или орнаменту, такие различия в основном вызваны половым отбором.[7]
Этим взглядам в определенной степени противостояли Альфред Рассел Уоллес, в основном после смерти Дарвина. Он допускал возможность полового отбора, но утверждал, что это относительно слабая форма отбора. Он утверждал, что соревнования самцов и самцов были формой естественного отбора, но что окраска «серой» павлины сама по себе адаптивна, поскольку камуфляж. По его мнению, приписывание выбора партнера самкам означает приписывание способности судить о стандартах красоты животным (таким как жуки ) слишком когнитивно неразвит, чтобы быть способным эстетический чувство.[8]
Рональд Фишер
Рональд Фишер, то английский статистик и эволюционный биолог разработал ряд идей о половом отборе в своей книге 1930 г. Генетическая теория естественного отбора в том числе гипотеза сексуального сына и Принцип фишера. В Беглый рыбак описывает, как половой отбор ускоряет предпочтение определенного украшения, в результате чего предпочтительный признак и женское предпочтение ему возрастают вместе в положительный отзыв безудержный цикл. В замечании, которое не было широко понято[9] еще 50 лет он сказал:
... развитие оперения у самцов и сексуальное предпочтение такого развития у самок, таким образом, должны продвигаться вместе, и до тех пор, пока этот процесс не сдерживается суровым контротбором, он будет продвигаться со все возрастающей скоростью. При полном отсутствии таких проверок легко увидеть, что скорость развития будет пропорциональна уже достигнутой, и поэтому со временем будет увеличиваться. экспоненциально, или в геометрическая прогрессия. —Рональд Фишер, 1930 год.

Это вызывает резкое увеличение как заметных черт самцов, так и их предпочтения самками, в результате чего половой диморфизм, пока практические физические ограничения не остановят дальнейшее преувеличение. А положительный отзыв петля создается, производя экстравагантные физические структуры неограниченного пола. Классическим примером женского выбора и потенциального беглого отбора является длиннохвостая вдова. В то время как у самцов длинные хвосты, выбираемые самками, предпочтения самок в отношении длины хвоста по-прежнему более резкие: самок привлекают более длинные хвосты, чем те, которые встречаются в природе.[10] Фишер понял, что предпочтение самок длинных хвостов может передаваться генетически в сочетании с генами самого длинного хвоста. Потомство длиннохвостой вдовы обоих полов наследует оба набора генов с самками. выражая их генетическое предпочтение к длинным хвостам, а самцы хвастаются желанным длинным хвостом.[9]
Ричард Докинз представляет нематематическое объяснение процесса беглого полового отбора в своей книге Слепой часовщик.[9] У самок, предпочитающих длиннохвостых самцов, как правило, матери выбирают длиннохвостых отцов. В результате они несут в себе оба набора генов. То есть гены длинных хвостов и предпочтения длинных хвостов становятся связаны. Пристрастие к длинным хвостам и сама длина хвоста могут поэтому коррелировать, имея тенденцию к увеличению вместе. Чем больше удлиняются хвосты, тем желательны более длинные хвосты. Любой незначительный начальный дисбаланс между вкусом и хвостом может вызвать взрывной рост длины хвоста. Фишер писал, что:
Экспоненциальный элемент, который является ядром предмета, возникает из-за того, что скорость изменения вкуса курицы пропорциональна абсолютной средней степени вкуса. -Рональд Фишер, 1932 год.[11]

Самка-вдова выбирает для спаривания самого привлекательного длиннохвостого самца, так что ее потомство, если он самец, само будет привлекательным для самок следующего поколения, тем самым породив много потомков, несущих гены самки. Поскольку скорость изменения предпочтений пропорциональна среднему вкусу среди женщин, и поскольку женщины желают пользоваться услугами наиболее сексуально привлекательных мужчин, создается дополнительный эффект, который, если его не остановить, может привести к экспоненциальному увеличению данного вкуса и в соответствующем желаемом сексуальном атрибуте.[9]
Важно отметить, что условия относительной стабильности, вызванные тем или иным способом, будут иметь гораздо более длительный срок, чем процесс, в котором развиваются украшения. У большинства существующих видов процесс бегства, должно быть, уже был остановлен, и мы должны ожидать, что более необычное развитие полового оперения вызвано, как большинство персонажей, не длительным и равномерным ходом эволюционного прогресса, а внезапными скачками изменений. -Рональд Фишер, 1930 год.
Поскольку первоначальная концептуальная модель процесса «бегства» Фишера, Рассел Ланде[12] и Питер О'Дональд[13] предоставили подробные математические доказательства, которые определяют обстоятельства, при которых может иметь место беглый половой отбор.
Теория
Репродуктивный успех

В репродуктивный успех организма измеряется количеством потомство оставленные позади, и их качество или вероятные фитнес.[14]
Сексуальное предпочтение создает тенденцию к ассортативная вязка или же гомогамия. Общие условия дискриминации по признаку пола выглядят так: (1) принятие одного партнера препятствует эффективному принятию альтернативных партнеров и (2) за отклонением предложения следуют другие предложения, безусловно или с такой высокой вероятностью, что риск ненаступления меньше, чем шанс получить преимущество при выборе партнера. Условия, определяющие, какой пол становится более ограниченным ресурсом при интерсексуальном отборе, были выдвинуты с учетом Принцип Бейтмана, в котором говорится, что пол, вкладывающий больше всего в производство потомства, становится ограничивающим ресурсом, за который конкурирует другой пол, что иллюстрируется инвестиции в питание яйца в зигота и ограниченная способность самок к воспроизводству; например, у людей женщина может рожать только каждые десять месяцев, тогда как мужчина может становиться отцом много раз за тот же период.[15] Совсем недавно исследователи усомнились в правоте Бейтмана. Хаббелл и Джонсон предположили, что вариативность репродуктивного успеха может зависеть от времени и распределения спариваний. В 2005 году Говати и Хаббелл предположили, что склонность к спариванию зависит от выбора стратегии; в некоторых случаях самцы могут быть более избирательными, чем самки, тогда как Бейтман предположил, что его парадигма будет «почти универсальной» среди видов, размножающихся половым путем. Критики предположили, что женщины могут быть более подвержены половому отбору, чем мужчины, но не во всех обстоятельствах.[16]
Современная интерпретация

Идеи Дарвина о половом отборе были встречены его современниками со скептицизмом и не имели большого значения, пока в 1930-х годах биологи не решили включить половой отбор в качестве способа естественного отбора.[17] Только в 21 веке они стали более важными в биология;[18] теория теперь рассматривается как общеприменимая и аналогичная естественному отбору.[19]

Десятилетнее исследование экспериментального варьирования полового отбора на мучные жуки при неизменных других факторах, показали, что половой отбор защищает даже инбредный население против вымирания.[20]

В принцип гандикапа из Амоц Захави, Рассел Ланде и В. Д. Гамильтон, считает, что тот факт, что самец способен дожить до репродуктивного возраста и пройти через него с такой, казалось бы, неадаптивной чертой, воспринимается самкой как свидетельство его общей приспособленности. Такие недостатки могут доказать, что он либо свободен, либо сопротивляется болезнь, или что он обладает большей скоростью или большей физической силой, которая используется для борьбы с проблемами, вызванными преувеличенной чертой.[нужна цитата ] Работа Захави стимулировала пересмотр этой области и несколько новых теорий. В 1984 году Гамильтон и Марлен Зук представил гипотезу «Умного самца», предполагающую, что мужские образования могут служить маркером здоровья, преувеличивая последствия болезней и недостатков.[нужна цитата ] В 1990 году Майкл Райан и А.С. Рэнд, работая с Тунгарская лягушка, предложили гипотезу «сенсорной эксплуатации», где преувеличенные мужские черты могут обеспечивать сенсорную стимуляцию, которой женщинам трудно сопротивляться.[21] Впоследствии теории «Гипотезы гравитации» Хорди Мойя-Ларано и др. (2002), используя простую биомеханическую модель для учета адаптивного значения скорости для более мелких пауков-самцов при лазании по вертикальным поверхностям,[22] и "Chase Away" Бретта Холланда и Уильяма Р. Райса были добавлены. В конце 1970-х Янзен и Мэри Уилсон, отметив, что мужские цветки часто больше женских, расширили сферу полового отбора на растения.[23]
В последние несколько лет эта область расширилась, включив в нее другие области исследования, не все из которых соответствуют дарвиновскому определению полового отбора. К ним относятся рогоносец, свадебные подарки, конкуренция спермы, детоубийство (особенно у приматов ), физический Красота, спаривание с помощью уловок, механизмы изоляции видов, родительская забота со стороны мужчин, амбипартильная забота, местонахождение партнера, полигамия и гомосексуальное изнасилование некоторых самцов животных.[нужна цитата ]
Ориентируясь на эффект сексуальный конфликт, как предположили Уильям Райс, Локк Роу и Йоран Арнвист, Тьерри Лоде утверждает, что расхождение интересов является ключом к эволюционному процессу. Сексуальный конфликт приводит к антагонистическая коэволюция в котором один пол стремится контролировать другой, что приводит к перетягиванию каната. Помимо, теория сексуальной пропаганды только утверждал, что спариванием руководили оппортунистически, на основе различных факторов, определяющих выбор, таких как фенотипические характеристики, кажущаяся сила особей, сила сигналов спаривания, трофические ресурсы, территориальность и т. д., которые могли объяснить поддержание генетического разнообразия в популяциях .[24]
Несколько рабочих обратили внимание на тот факт, что разработанные характеристики, которые должны быть так или иначе дорогостоящими для их носителей (например, хвост рыбы-меч Xiphophorus montezumae ) не всегда имеют цену с точки зрения энергии, производительности или даже выживания. Одно из возможных объяснений очевидного отсутствия затрат заключается в том, что «компенсирующие черты» эволюционировали вместе с признаками, отобранными половым путем.[25]
Инструментарий естественного отбора

Половой отбор может объяснить, почему определенные характеристики (например, перья) имели определенную ценность для выживания на ранней стадии их эволюции. Джеффри Миллер предполагает, что половой отбор мог способствовать созданию эволюционных модулей, таких как Археоптерикс поначалу перья как сексуальные украшения. Самые ранние протоптицы, такие как китайские Протархеоптерикс, обнаруженный в начале 1990-х годов, имел хорошо развитые перья, но не имел никаких признаков асимметрии верха / низа, придающей крыльям подъем. Некоторые предположили, что перья служили изоляцией, помогая самкам насиживать яйца. Но, возможно, перья служили сексуальными украшениями, которые до сих пор распространены у большинства видов птиц, особенно у таких птиц, как павлины и другие. райские птицы сегодня. Если бы при ухаживании за протоптицами отображалось сочетание перьев на передних конечностях с энергичными прыжками, то переход от демонстрации к аэродинамическим функциям мог быть относительно плавным.[26]
Половой отбор иногда порождает признаки, которые могут способствовать исчезновению вида, как предполагалось.[26] за гигантские рога Ирландский лось (Мегалоцерос гигантский), которые вымерли в Плейстоцен Европа.[27] Однако половой отбор может и наоборот, приводя к расхождению видов - иногда за счет сложных изменений в гениталии[28]- так появляются новые виды.[29][30][31]
Половой диморфизм
Половые различия напрямую связаны с воспроизводством и не служат прямой цели в ухаживание называются первичные половые признаки. Признаки, поддающиеся половому отбору, которые дают организму преимущество перед его соперниками (например, в ухаживании) без непосредственного участия воспроизведение, называются вторичные половые признаки.
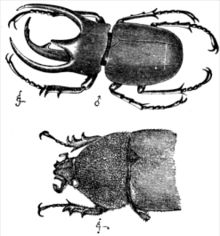
У большинства половых видов самцы и самки иметь разные равновесие стратегии из-за разницы в относительных инвестициях в производство потомства. Как сформулировано в принципе Бейтмана, у самок больше первоначальных вложений в производство потомства (беременность у млекопитающих или производство яйца в птицы и рептилии ), и эта разница в начальных инвестициях создает различия в ожидаемых репродуктивных успехах и стимулирует процессы полового отбора. Классические примеры видов с обратной половой ролью включают игла, и Фаларопа Вильсона.[32] Также, в отличие от женщины, мужчина (кроме моногамный Вид) имеет некоторую неуверенность в том, является ли он истинным родителем ребенка, и поэтому менее заинтересован в том, чтобы тратить свою энергию на воспитание потомства, которое может быть связано с ним, а может и нет. В результате этих факторов можно ожидать, что самцы будут более склонны к спариванию, чем самки, в то время как самки будут делать выбор (кроме случаев принудительные совокупления, что может произойти, например, в некоторых приматы и утки ). Таким образом, эффекты полового отбора часто более выражены у мужчин, чем у женщин.
Различия во вторичных половых признаках между самцами и самками одного вида называются половые диморфизмы. Они могут быть столь же незначительными, как разница в размерах (половой диморфизм размера, часто сокращенно SSD), или столь же резкими, как рога и цветовые узоры. Половые диморфизмы изобилуют в природе. Примеры включают владение рогами только мужчинами. олень, более яркая окраска многих самцов птиц по сравнению с самками того же вида или даже более четкие различия в базовой морфологии, такие как резко увеличенный размах глаз самцов. черноглазая муха. В павлин, с его сложными и красочными перьями хвоста, которые пава недостатков, часто называют, пожалуй, самым необычным примером диморфизма. Мужской и женский чернозобые синие певчие птицы и Гвианский петух из скал также кардинально отличаются оперением. Ранние натуралисты даже считали самок отдельным видом. Самый большой половой диморфизм размера в позвоночные это оболочка цихлида рыбы Neolamprologus callipterus самцы в 30 раз больше самок.[33] Многие другие рыбы, такие как гуппи сексуально диморфны. Чрезвычайный половой диморфизм размеров, когда самки крупнее самцов, довольно часто встречается у пауки и хищные птицы.
Поддержание полового размножения в мире жесткой конкуренции - одна из главных загадок биологии, учитывая, что бесполое размножение могут воспроизводиться намного быстрее, поскольку 50% потомства не являются самцами и не могут сами производить потомство. Было предложено много неисключительных гипотез,[34] включая положительное влияние дополнительной формы отбора, полового отбора, на вероятность сохранения вида.[20]
Внутрисексуальная конкуренция между мужчинами
Соревнование между самцами происходит, когда два самца одного вида соревнуются за возможность спариться с самкой. Половой диморфизм, размер, соотношение полов,[35] и социальная ситуация[36] Все они могут играть определенную роль в влиянии конкуренции между мужчинами на репродуктивный успех самца и выбор партнера самки. Более крупные самцы, как правило, побеждают в конфликтах между самцами из-за своей абсолютной силы и способности удерживать других самцов от захвата их самок. Например, на лету Дриомица анилисовая, размер показывает самую сильную корреляцию с исходом мужско-мужских конфликтов из-за ресурсов, таких как территория и женщины.[37]
Влияющие факторы
Соотношение полов

Существует несколько типов соревнований между мужчинами, которые могут происходить в популяции в разное время в зависимости от условий. Вариация конкуренции происходит в зависимости от частоты различных видов брачного поведения в популяции.[35] Одним из факторов, который может повлиять на тип наблюдаемой конкуренции, является плотность населения мужчин.[35] Когда в популяции присутствует высокая плотность самцов, конкуренция имеет тенденцию быть менее агрессивной, и поэтому чаще используются тактика скрытности и методы разрушения.[35] Эти методы часто указывают на тип соревнования, называемого схваткой.[35] В японской медаке Oryzias latipes, подкрадывание относится к тому, когда самец прерывает спаривающуюся пару во время совокупления, хватаясь либо за самца, либо за самку и выпуская свою собственную сперму в надежде оплодотворить самку.[35] Разрушение - это техника, при которой один самец натыкается на самца, который совокупляется с самкой, прямо перед тем, как его сперма высвобождается и яйцеклетки оплодотворяются.[35]
Однако не все методы одинаково успешны в борьбе за репродуктивный успех. Нарушение приводит к более короткому периоду копуляции и, следовательно, может нарушить оплодотворение яйцеклеток спермой, что часто приводит к более низкой скорости оплодотворения и меньшему размеру кладки.[35]
Ресурсная ценность и социальный рейтинг
Еще один фактор, который может влиять на конкуренцию между мужчинами, - это ценность ресурса для конкурентов. Соревнование между мужчинами может представлять множество рисков для физической формы мужчин, таких как большие затраты энергии, физические травмы, более низкое качество спермы и потеря отцовства.[38] Следовательно, риск конкуренции должен оправдывать ценность ресурса. Мужчина с большей вероятностью будет участвовать в соревновании за ресурс, который улучшает их репродуктивный успех, если ценность ресурса выше. В то время как конкуренция между мужчинами и мужчинами может происходить в присутствии или в отсутствие женщины, конкуренция чаще происходит в присутствии женщины.[36] Присутствие самки напрямую увеличивает ресурсную ценность территории или укрытия, поэтому самцы с большей вероятностью примут на себя риск конкуренции, когда присутствует самка.[36] Более мелкие самцы вида также с большей вероятностью вступят в соревнование с более крупными самцами в присутствии самки.[36] Из-за более высокого уровня риска для подчиненных самцов они, как правило, участвуют в соревнованиях реже, чем более крупные, доминирующие самцы, и поэтому размножаются реже, чем доминирующие самцы.[38] Это наблюдается у многих видов, таких как древесная лягушка Омей, Ракофор омеймонтис, где более крупные самцы получают больше возможностей для спаривания и спариваются с более крупными самками.[39]
Эффекты победителя и проигравшего
Третий фактор, который может повлиять на успех мужчины в соревнованиях, - это эффекты победителя-проигравшего.[40] Роющие сверчки, Веларификтор асперсус, соревнуйтесь за норы, чтобы привлечь самок, используя свои большие челюсти для боя.[40] Самки норных сверчков с большей вероятностью выберут победителя соревнования через 2 часа после боя.[40] Присутствие победившего самца подавляет брачное поведение проигравших самцов, потому что победивший самец имеет тенденцию производить более частые и усиленные брачные крики в этот период времени.[40]
Влияние на женский фитнес
Соревнование между мужчинами может как положительно, так и отрицательно повлиять на женскую физическую форму. Когда в популяции высока плотность самцов и большое количество самцов пытается спариваться с самкой, она с большей вероятностью будет сопротивляться попыткам спаривания, что приведет к снижению уровня оплодотворения.[35] Высокий уровень конкуренции между самцами также может привести к сокращению инвестиций самок в спаривание.[38] Многие формы конкуренции также могут причинить женщинам серьезные страдания, что отрицательно скажется на ее способности к воспроизводству.[35] Увеличение конкуренции между самцами может повлиять на способность самок выбирать лучших партнеров и, следовательно, снизить вероятность успешного воспроизводства.[41]
Однако групповые спаривания по-японски Медака было показано, что он положительно влияет на физическую форму самок из-за увеличения генетической изменчивости, более высокой вероятности заботы со стороны отца и более высокой вероятности успешного оплодотворения.[35]
В разных таксонах
- Подробнее см. Половой отбор в ...

Половой отбор наблюдается у растений, животных и грибов.[5] В определенных гермафродитный улитка и слизняк виды моллюски бросание люблю дартс это форма полового отбора.[42] Определенный мужчина насекомые порядка Чешуекрылые цементируют поры влагалища своих самок.[43]

Сегодня биологи говорят, что некоторые эволюционные черты можно объяснить внутривидовая конкуренция - конкуренция между представителями одного и того же вида - различие между конкуренцией до и после половой акт.

Перед совокупление, интрасексуальный отбор- обычно между мужчинами - может принимать форму бой между мужчинами. Также, интерсексуальный отбор, или же выбор партнера, происходит, когда самки выбирают между самцами.[44] Черты, выбранные в ходе мужских боев, называются вторичными половыми признаками (включая рога, рога и т. Д.), Которые Дарвин описал как «оружие», в то время как черты, выбранные по выбору партнера (обычно женского пола), называются «украшениями». Из-за своей иногда сильно преувеличенной природы вторичные половые признаки могут оказаться помехой для животного, тем самым снижая его шансы на выживание. Например, большие рога лося массивны и тяжелы и замедляют бегство существа от хищников; они также могут запутаться в низко свисающих ветвях деревьев и кустах и, несомненно, привели к гибели многих людей. Яркая окраска и эффектные орнаменты, подобные тем, которые можно увидеть у многих самцов птиц, не только привлекают внимание самок, но и привлекают внимание хищников. Некоторые из этих черт также представляют собой энергетически затратные вложения для животных, которые их несут. Поскольку черты, которые, как считается, являются результатом полового отбора, часто вступают в конфликт с приспособленностью к выживанию индивидуума, возникает вопрос, почему в природе, в какой выживание сильнейшего считается практическим правилом, такие очевидные обязательства могут сохраняться. Однако следует также учитывать, что интерсексуальный отбор может происходить с упором на ресурсы, которыми обладает один пол, а не на морфологические и физиологические различия. Например, самцы Euglossa imperialis, несоциальный вид пчел, образуют скопления территорий, которые считаются токами, для защиты богатых ароматами первичных территорий. Цель этих скоплений носит лишь факультативный характер, поскольку чем больше подходящих участков, богатых ароматными веществами, тем больше там обитаемых территорий для обитания, что дает самкам этого вида большой выбор самцов, с которыми они могут потенциально спариваться.[45]
После совокупления мужское соревнование отличается от обычного агрессия может принять форму конкуренция спермы, как описано Паркер[46] в 1970 году. В последнее время возник интерес к загадочный женский выбор,[47] феномен внутренне оплодотворенных животных, таких как млекопитающие и птицы, когда самка может избавиться от самца сперма без его ведома.

Ну наконец то, сексуальный конфликт считается, что происходит между партнерами по размножению,[48] иногда приводя к эволюционная гонка вооружений между мужчинами и женщинами. Половой отбор также может происходить в результате феромон выпуск, например, с пчелой без жала, Тригона корвина.[49]
Широко признано, что предпочтения самок при спаривании ответственны за быструю и дивергентную эволюцию вторичных половых признаков самцов.[50] Самки многих видов животных предпочитают спариваться с самцами с внешними украшениями - преувеличенными чертами морфологии, такими как сложные половые органы. Эти предпочтения могут возникать, когда произвольное женское предпочтение какому-либо аспекту мужской морфологии - первоначально, возможно, в результате генетический дрейф - создает со временем селекцию самцов с соответствующим орнаментом. Одна интерпретация этого известна как гипотеза сексуального сына. С другой стороны, гены, которые позволяют мужчинам развивать впечатляющие украшения или боевые способности, могут просто демонстрировать большую устойчивость к болезням или более эффективный метаболизм, функции, которые также приносят пользу женщинам. Эта идея известна как гипотеза хороших генов.
Яркие цвета, которые развиваются у животных во время брачного сезона, привлекают партнеров. Было высказано предположение, что существует причинная связь между силой демонстрации украшений, участвующих в половом отборе, и биологией свободных радикалов.[51] Для проверки этой идеи были проведены эксперименты на самцах. нарисованные драконы ящерицы.[52] Самцы ящериц ярко выделяются по своей окраске для размножения, но с возрастом их окраска снижается. Эксперименты с введением антиоксиданты к этим самцам привели к выводу, что племенная окраска является отражением врожденной антиоксидантной способности, которая защищает от окислительное повреждение, включая окислительное повреждение ДНК.[52] Таким образом, цвет может действовать как «сертификат здоровья», который позволяет самкам визуализировать лежащие в основе повреждения, вызванные окислительным стрессом, у потенциальных партнеров.
Дарвин предположил, что наследственный такие черты характера, как бороды и отсутствие волос в различных популяциях человека, являются результатом половой отбор у людей. Джеффри Миллер выдвинул гипотезу о том, что многие виды человеческого поведения, явно не связанные с преимуществами выживания, такие как юмор, музыка, изобразительное искусство, вербальное творчество и некоторые формы альтруизм, являются приспособлениями для ухаживания, которым способствовал половой отбор. С этой точки зрения многие человеческие артефакты могут рассматриваться как подлежащие половому отбору как часть расширенный фенотип, например, одежда, подчеркивающая сексуально выбранные черты[нужна цитата ]. Некоторые утверждают, что эволюция человеческого интеллекта - это признак, выбранный половым путем, так как сам по себе он не обеспечивает достаточной приспособленности по сравнению с высокими затратами на содержание.[53]
Рекомендации
Цитаты
- ^ Сеси Старр (2013). Биология: единство и разнообразие жизни (Ральф Таггарт, Кристин Эверс, Лиза Старр, ред.). Cengage Learning. п. 281.
- ^ Фогт, Ингве (29 января 2014 г.). «Большие яички связаны с изменой». Phys.org. В архиве с оригинала 31 января 2014 г.. Получено 31 января, 2014.
- ^ Дарвин, Чарльз; А. Р. Уоллес (1858). «О склонности видов к формированию разновидностей; и о сохранении разновидностей и видов естественными средствами отбора» (PDF). Журнал трудов Линнеевского общества Лондона. Зоология. 3 (9): 46–50. Дои:10.1111 / j.1096-3642.1858.tb02500.x. В архиве (PDF) из оригинала от 22.10.2012.
- ^ Мур, Джейми С.; Паннелл, Джон Р. (2011). «Половой отбор у растений». Текущая биология. 21 (5): R176 – R182. Дои:10.1016 / j.cub.2010.12.035. PMID 21377091. S2CID 18044399.
- ^ а б Nieuwenhuis, B.P.S .; Aanen, D.K. (2012). «Половой отбор у грибов». Журнал эволюционной биологии. 25 (12): 2397–2411. Дои:10.1111 / jeb.12017. PMID 23163326. S2CID 5657743.
- ^ Дарвин, Чарльз (1859). О происхождении видов (1-е издание). Глава 4, с. 88. «И это заставляет меня сказать несколько слов о том, что я называю половым отбором. Это зависит от ...» «Архивная копия». В архиве из оригинала 2011-11-05. Получено 2011-05-22.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Дарвин, Чарльз (1859). О происхождении видов (1-е издание). Глава 4, с. 89. «Архивная копия». В архиве из оригинала 2011-11-05. Получено 2011-05-22.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Уоллес, Альфред Рассел (1892). «Записка о половом отборе (S459: 1892)». Смит, Чарльз. В архиве из оригинала 17 февраля 2017 г.. Получено 13 января 2017.
- ^ а б c d Докинз, Ричард (1996). Слепой часовщик: почему свидетельства эволюции раскрывают Вселенную без замысла. Нортон. С. Глава 8, Взрывы и спирали. ISBN 978-0-393-31570-7.
- ^ Андерссон, М. (1994). Половой отбор. Издательство Принстонского университета.[страница нужна ]
- ^ Рональд Фишер в письме к Чарльз Гальтон Дарвин, 22 ноября 1932 г., цит. По: Фишер Р. А., Беннетт Дж. Х. 1999. Генетическая теория естественного отбора: полное издание variorum, Oxford University Press, Oxford, p. 308
- ^ Ланде, Р. (1981). «Модели видообразования путем полового отбора по полигенным признакам». PNAS. 78 (6): 3721–3725. Bibcode:1981PNAS ... 78.3721L. Дои:10.1073 / pnas.78.6.3721. ЧВК 319643. PMID 16593036. Ланде, Р. (1981). «Модели видообразования путем полового отбора по полигенным признакам». Труды Национальной академии наук. 78 (6): 3721–3725. Bibcode:1981PNAS ... 78.3721L. Дои:10.1073 / pnas.78.6.3721. ЧВК 319643. PMID 16593036.
- ^ О'Дональд, Питер (1980). Генетические модели полового отбора. Издательство Кембриджского университета, Нью-Йорк. 250 стр.
- ^ Орр, Х.А. (август 2009 г.). «Фитнес и его роль в эволюционной генетике». Природа Обзоры Генетика. 10 (8): 531–9. Дои:10.1038 / nrg2603. ЧВК 2753274. PMID 19546856.
- ^ Бейтман, А.Дж. (1948). «Внутриполовой отбор у дрозофилы». Наследственность. 2 (Pt. 3): 349–368. Дои:10.1038 / hdy.1948.21. PMID 18103134.
- ^ Новичок, Скотт Д.; Zeh, Jeanne A .; Zeh, Дэвид В. (31 августа 1999 г.). «Генетические преимущества повышают репродуктивный успех полиандрических самок». Труды Национальной академии наук. 96 (18): 10236–10241. Bibcode:1999ПНАС ... 9610236Н. Дои:10.1073 / пнас.96.18.10236. ISSN 0027-8424. ЧВК 17872. PMID 10468592.
- ^ Миллер, Джеффи, Брачный разум, п. 24
- ^ Сексуальный отбор и разум В архиве 2015-06-10 на Wayback Machine
- ^ Хоскен, Дэвид Дж .; Хаус, Кларисса М. (январь 2011 г.). «Половой отбор». Текущая биология. 21 (2): R62 – R65. Дои:10.1016 / j.cub.2010.11.053. PMID 21256434. S2CID 18470445.
- ^ а б c Польза для населения от полового отбора объясняет существование самцов Phys.org 18 мая 2015 г. по исследованию Университет Восточной Англии В архиве 21 августа 2015 г. Wayback Machine
- ^ а б Райан, Майкл Дж .; Фокс, Джеймс Х .; Вильчинский, Вальтер; Рэнд, А. Стэнли (1990). «Половой отбор для сенсорной эксплуатации у лягушки Physalaemus pustulosus». Природа. 343 (6253): 66–67. Дои:10.1038 / 343066a0. ISSN 0028-0836. PMID 2296291. S2CID 4358189.
- ^ Моя-Лараньо, Дж. (2007). "Гравитация все еще имеет значение". Функциональная экология. 21 (6): 1178–1181. Дои:10.1111 / j.1365-2435.2007.01335.x. S2CID 7951226.
- ^ Уилсон, Мэри Ф (июнь 1979). «Половой отбор у растений». Американский натуралист. 113 (6): 777–790. Дои:10.1086/283437. S2CID 84970789.
- ^ Тьерри Лоде (2006). La guerre des sexes chez les animaux Редакторы Odile Jacob, Париж. ISBN 2-7381-1901-8
- ^ Уфьеро, Кристофер Э. (май 2015 г.). «Половой отбор, затраты и компенсация». Калифорнийский университет в Риверсайде. Архивировано из оригинал 6 июня 2014 г.
- ^ а б Миллер, Джеффри (2000). Брачный разум. Якорь Букс, подразделение Рэндом Хаус, Инк. (Первое издание якорных книг, 2001). Нью-Йорк. ISBN 0-385-49517-X
- ^ Гулд, Стивен Джей (1974). «Происхождение и функция« причудливых »структур - размер рога и размер черепа у« ирландского лося », Megaloceros giganteus». Эволюция. 28 (2): 191–220. Дои:10.2307/2407322. JSTOR 2407322. PMID 28563271.
- ^ Эберхард, Уильям Г. (24 марта 2009 г.). «Эволюция гениталий: теории, доказательства и новые направления» (PDF). Genetica. 138 (1): 5–18. Дои:10.1007 / s10709-009-9358-у. ISSN 0016-6707. PMID 19308664. S2CID 1409845.
- ^ Хоскен, Дэвид Дж. И Паула Стокли. "Половой отбор и генитальная эволюция В архиве 2017-10-12 в Wayback Machine." Тенденции в экологии и эволюции 19.2 (2004): 87–93.
- ^ Арнквист, Горан. "Сравнительные данные об эволюции гениталий путем полового отбора В архиве 2012-01-27 в Wayback Machine. "Nature 393.6687 (1998): 784.
- ^ Эберхард, В. Г. (1985). Половой отбор и гениталии животных. Издательство Гарвардского университета, Кембридж, Массачусетс.[страница нужна ]
- ^ Sagebakken, Gry; Кварнемо, Шарлотта; Анесйо, Ингрид (22 марта 2017 г.). «Состояние питания - набор для выживания отцов-производителей морских игл». Биологический журнал Линнеевского общества. 121 (2): 312–318. Дои:10.1093 / биолиннеан / blx002. ISSN 0024-4066.
- ^ Ота, Казутака; Кохда, Масанори; Сато, Тецу (1 мая 2010 г.). «Необычная аллометрия полового диморфизма размеров у цихлид, где самцы намного крупнее самок». Журнал биологических наук. 35 (2): 257–265. Дои:10.1007 / s12038-010-0030-6. ISSN 0250-5991. PMID 20689182. S2CID 12396902.
- ^ Хартфилд, Мэтью; П. Д. Кейтли (2012). «Современные гипотезы эволюции пола и рекомбинации» (PDF). Интегративная зоология. 7 (2): 192–209. Дои:10.1111 / j.1749-4877.2012.00284.x. PMID 22691203. В архиве (PDF) из оригинала от 26.08.2016.
- ^ а б c d е ж грамм час я j k Вейр, Лаура К. (22 ноября 2012 г.). «Соревнование между самцами и альтернативная тактика спаривания самцов влияют на поведение и плодовитость самок в японской медаке (Oryzias latipes)». Поведенческая экология и социобиология. 67 (2): 193–203. Дои:10.1007 / s00265-012-1438-9. ISSN 0340-5443. S2CID 15410498.
- ^ а б c d Проктор, Д. С .; Мур, А. Дж .; Миллер, К. В. (2012-03-09). "The form of sexual selection arising from male-male competition depends on the presence of females in the social environment". Журнал эволюционной биологии. 25 (5): 803–812. Дои:10.1111/j.1420-9101.2012.02485.x. ISSN 1010-061X. PMID 22404372. S2CID 594384.
- ^ Otronen, Merja (1984-08-01). "Male contesis for territories and females in the fly Dryomyza Anilis". Поведение животных. 32 (3): 891–898. Дои:10.1016/S0003-3472(84)80167-0. ISSN 0003-3472. S2CID 53188298.
- ^ а б c Nelson-Flower, Martha J.; Ridley, Amanda R. (2015-09-24). "Male-male competition is not costly to dominant males in a cooperatively breeding bird". Поведенческая экология и социобиология. 69 (12): 1997–2004. Дои:10.1007/s00265-015-2011-0. ISSN 0340-5443. S2CID 15032582.
- ^ Luo, Zhenhua; Li, Chenliang; Ван, Хуэй; Shen, Hang; Чжао, Миан; Gu, Qi; Liao, Chunlin; Gu, Zhirong; Wu, Hua (2016-02-23). "Male-male competition drives sexual selection and group spawning in the Omei treefrog, Rhacophorus omeimontis". Поведенческая экология и социобиология. 70 (4): 593–605. Дои:10.1007/s00265-016-2078-2. ISSN 0340-5443. S2CID 13912038.
- ^ а б c d Zeng, Yang; Zhou, Feng-Hao; Zhu, Dao-Hong (2018-06-26). "Fight outcome briefly affects the reproductive fitness of male crickets". Научные отчеты. 8 (1): 9695. Bibcode:2018NatSR...8.9695Z. Дои:10.1038/s41598-018-27866-4. ISSN 2045-2322. ЧВК 6018733. PMID 29946077.
- ^ Cayuela, Hugo; Lengagne, Thierry; Kaufmann, Bernard; Жоли, Пьер; Léna, Jean-Paul (2016-06-24). "Larval competition risk shapes male–male competition and mating behavior in an anuran". Поведенческая экология. 27 (6): arw100. Дои:10.1093/beheco/arw100. ISSN 1045-2249.
- ^ Рассказы о двух улитках: половой отбор и сексуальный конфликт в Lymnaea stagnalis и Helix aspersa Oxford Journals В архиве 2015-09-04 в Wayback Machine
- ^ von Byern, Janek; Grunwald, Ingo (2011). Biological Adhesive Systems: From Nature to Technical and Medical Application. Springer Science & Business Media. п. 124. ISBN 9783709102862.
- ^ Campbell, N. A.; J. B. Reece (2005). Биология. Бенджамин Каммингс. п. 1230. ISBN 978-0-8053-7146-8.
- ^ Кимси, Линн Сири (1980). «Поведение самцов орхидейных пчел (Apidae, Hymenoptera, Insecta) и вопрос о токах». Поведение животных. 28 (4): 996–1004. Дои:10.1016 / с0003-3472 (80) 80088-1. S2CID 53161684.
- ^ Parker, Geoffrey A. (1970). "Sperm competition and its evolutionary consequences in the insects". Биологические обзоры. 45 (4): 525–567. Дои:10.1111/j.1469-185x.1970.tb01176.x. S2CID 85156929.
- ^ Eberhard, WG. (1996) Female control: Sexual selection by cryptic female choice. Princeton, Princeton University Press.
- ^ Locke Rowe, Göran Arnvist. (2013) Sexual conflict, Princeton Univ Press.[страница нужна ]
- ^ Jarau, Stefan; Dambacher, Jochen; Твеле, Роберт; Aguilar, Ingrid; Francke, Wittko; Ayasse, Manfred (2010). "The Trail Pheromone of a Stingless Bee, Trigona corvina (Hymenoptera, Apidae, Meliponini), Varies between Populations". Химические чувства. 35 (7): 593–601. Дои:10.1093/chemse/bjq057. ISSN 0379-864X. PMID 20534775. В архиве from the original on 2016-01-01.
- ^ Andersson M. (1994). Sexual Selection. Princeton Univ Press, Princeton, NJ.[страница нужна ]
- ^ von Schantz T, Bensch S, Grahn M, Hasselquist D, Wittzell H (January 1999). "Good genes, oxidative stress and condition-dependent sexual signals". Proc. Биол. Наука. 266 (1414): 1–12. Дои:10.1098/rspb.1999.0597. ЧВК 1689644. PMID 10081154.
- ^ а б Olsson M, Tobler M, Healey M, Perrin C, Wilson M (August 2012). "A significant component of ageing (DNA damage) is reflected in fading breeding colors: an experimental test using innate antioxidant mimetics in painted dragon lizards". Эволюция. 66 (8): 2475–2483. Дои:10.1111/j.1558-5646.2012.01617.x. PMID 22834746. S2CID 205783815.
- ^ PLoS ONE: Sexual Selection and the Evolution of Brain Size in Primates В архиве 2007-09-30 на Wayback Machine
Источники
- Andersson, M. (1994) Половой отбор. Princeton University Press. ISBN 0-691-00057-3
- Arnqvist, G. & Rowe, L. (2013) Sexual conflict. Princeton University Press
- Lande, C. F. R. (1981). "Models of speciation by sexual selection on polygenic traits" (PDF). Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 78 (6): 3721–3725. Bibcode:1981PNAS...78.3721L. Дои:10.1073/pnas.78.6.3721. ЧВК 319643. PMID 16593036. Архивировано из оригинал (PDF) на 2007-09-27.
- Cronin, H. (1991) The ant and the peacock: altruism and sexual selection from Darwin to today. Издательство Кембриджского университета.
- Дарвин, К. (1871) Происхождение человека и отбор в отношении пола. Джон Мюррей, Лондон.
- Eberhard, W. G. (1996) Female control: Sexual selection by cryptic female choice. Princeton, Princeton University Press.
- Fisher, R. A. (1930) Генетическая теория естественного отбора. Издательство Оксфордского университета, ISBN 0-19-850440-3, Глава 6 Memeoid.net
- Lodé, T. (2006) La guerre des sexes chez les animaux. Eds Odile Jacob. ISBN 2-7381-1901-8
- Miller, G. F. (1998) How mate choice shaped human nature: A review of sexual selection and human evolution. In: C. Crawford & D. Krebs (Eds.) Handbook of evolutionary psychology: Ideas, issues, and applications. Lawrence Erlbaum, pp. 87–129
- Miller, G. F. (2000) Брачный разум: как сексуальный выбор повлиял на эволюцию человеческой природы. Хайнеманн, Лондон. ISBN 0-434-00741-2
- Rosenberg, J. & Tunney, R. J. (2008). Human vocabulary use as display. Evolutionary Psychology, 6, 538–549
внешняя ссылка
- Natural & Sexual Selection Videos by the Cornell Lab of Ornithology
- Sexual Selection: Stanford University
- The Great Debate: Sexual Selection
- Intralocus Sexual Conflict Diminishes the Benefits of Sexual Selection
- A New Interpretation of Natural Beauty and Sexual Selection
- Review of GF Miller's The Mating Mind
- Why Sex Evolved
- "The sexual propaganda theory"
| Базовые концепты | ||
|---|---|---|
| Geographic modes | ||
| Isolating factors | ||
| Гибридизация | ||
| Speciation in taxa | ||
| Биологические термины | |
|---|---|
| Сексуальный воспроизведение | |
| Сексуальность | |
| |

| Узоры |  | |
|---|---|---|
| Причины | ||
| Люди | ||
| Связанный | ||